One of my favourite museums in the world, and certainly one of the best natural history museums in the UK, is Cambridge’s Museum of Zoology, AKA “University Museum of Zoology at Cambridge” (UMZC). It is now nearing a lengthy completion of renovations; the old museum exhibits and collections were excellent but needed some big changes along with the re-fabbed “David Attenborough Building” that houses them. As a longtime fan of the exhibits and user of the collection (and microCT scanner), I hurried to see the new museum once it officially opened.
And that makes a great excuse to present a photo-shoot from my visit. This focuses on the “mammal floor” below the entrance- the upper floor(s?) are still being completed and will have the birds, non-avian tetrapods, fish, etc. But the UMZC is strong in mammals and so it is natural for them to feature them in this chock-full-o-specimens display. Less talk, more images. Here we go!
All images can be clicked to mu-zoom in on them.
Stomach-Churning Rating: 3/10; bones and taxidermy and innocuous jars.
The building. The whale skeleton that hung outside for years is now cleaned up and housed right inside; you walk under it as you enter.
Entrance.
First view past the entryway: lots of cool specimens.
View from the walkway down into the ground/basement level from the entry. As specimens-per-unit-volume goes, the UMZC still scores highly and that is GOOD!
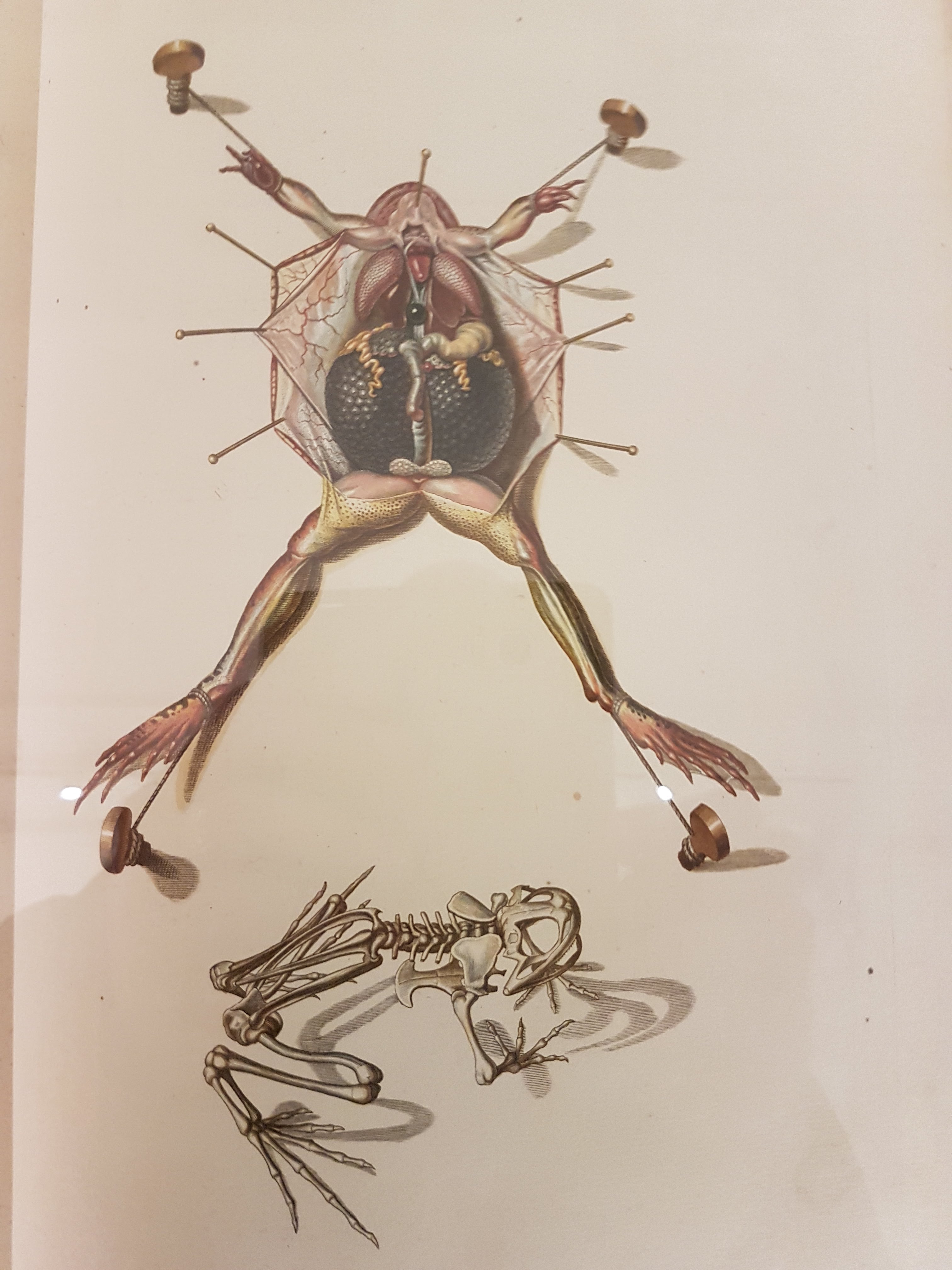
Explanation of frog dissection image below.
Gorgeous old frog dissection illustration; such care taken here.
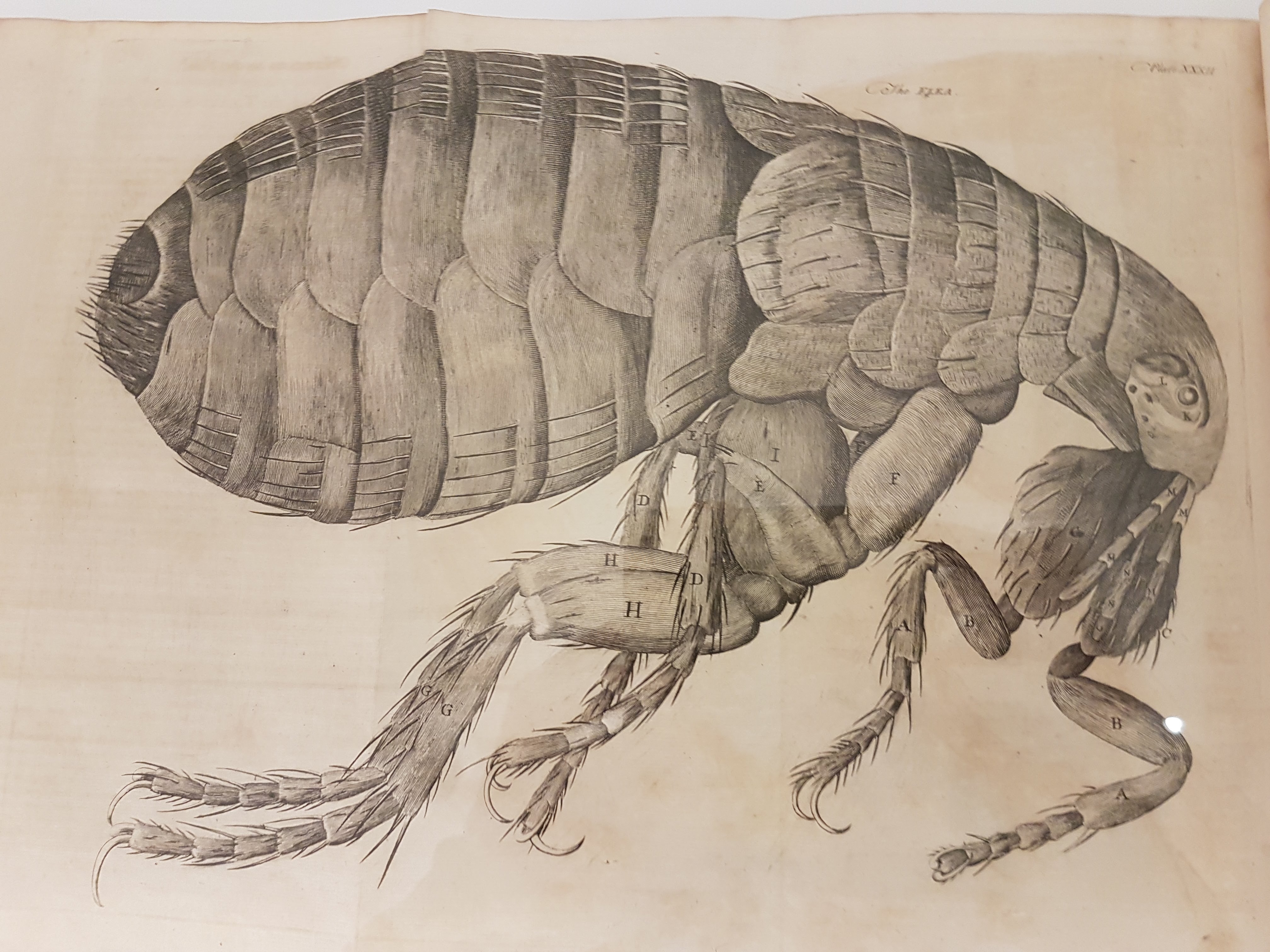
Leeuwenhoek’s flea woodcut; I think from Arcana Naturae Detecta (1695). There is an impressive display of classic natural history books near the entryway.
Dürer/other rhino art image and info.
Darwin was famed for collecting beetles when he should have been studying theology at Cambridge as a youth, and here is some of his collection. Dang.
Darwin’s finches!
Darwin kicked off some of his meticulous work with volumes on barnacles; specimens included here; which helped fuel insights into evolution (e.g. they are “retrograde” crustaceans, not mollusks).
Darwin’s voyage: fish & other preserved specimens.
I think this is a solitaire weka (flightless island bird; see Comment below). I’ve never seen them displayed w/skeleton + taxidermy; it’s effective here.
Eryops cast. More early tetrapods will surely be featured on the upper floor; this one was on the timeline-of-life-on-Earth display.
I LOVE dioramas and this seabird nesting ground display is very evocative, especially now that I’ve visited quite a few such islands.
Mammal introduction; phylogenetic context.
Monotreme glory.
UMZC is well endowed with thylacines and this one is lovely.
“TAZ FEEL NAKED!”
Narwhal above!
Rhinocerotoidea past, present, and fading glory. 😦
Ceratotherium white rhino. The horn is not real; sadly museums (and even zoos) across the world have to worry about theft of such things, given that some people think these horns are magic.
Ceratotherium staring match. You lose.
Ceratotherium stance.
Foot of a Sumatran rhino juxtaposed with a horse’s for Perissodactyla didaction.
A tapir. As a kid, I used to wander around the house pretending to be a tapir but I did not know what noise they’d make so I’d say “tape tape tape!”.
Big Southern Elephant Seal.
Squat little fur seal.
Hippopotamus for the lot of us. (baby included)
Hippo facedown.
Skull of a dwarf Madagascar hippo.
Cave bear and sabretooth cat make an impressive Ice Age demo.
It’s a wombat.
Ain’t no don like a Diprotodon! (also note its modern miniature cousin the wombat, below)
Diprotodon facial.
Diprotodon shoulder: big clavicles bracing that joint region.
Diprotodon knee: even in big marsupials, the “parafibula”/lateral sesamoid of the knee is still generally present. And why it is there/what it does deserves much more study.
Diprotodon hip. I just find this animal’s anatomy fascinating head-to-tail.
Diprotodon front foot. Absolutely freakish.
Diprotodon hind foot. Even weirder.
Your view after having been trampled in a supine position by a Diprotodon. Not a good way to go.
Diprotodon got back.
Elephant seal’s butt continues my series of photos of big animals’ bottoms.
Asian elephant’s butt view.
African elephant butt.
Sectioned elephant skull to show pneumatic resonating chambers.
Paenungulates: hyraxes, Sirenia, elephants & kin (evolutionary demo).
AND MY HYRAX! Sorry. Had to.
Megatherium side view.
Megatherium. Yeah!
Megatherium hindlegs fascinate me. Well-heeled.
Tamandua duo.
Silky anteater; wonderful.
Armadillos.
Anteaters round out a fab display on Xenarthra.
The UMZC has everything from aardvarks to zebus. Here, conceptualized with other Afrotheria.
Golden moles: the more I read about them, the more they fascinate me.
We can all use some more solenodons in our lives!
Example of the phylogenetic context used throughout exhibits.
If you’ve got a good Okapi taxidermy, you’d better use it.
It’s a giraffe. Did you guess right?
Gerenuk showing off its bipedal capacity.
Warthogs have an inner beauty.
Pangolin. Glad to see it back on exhibit.
Nice little brown bear.
Double-barrelled shot of hyenas.
Colugo!
Nice to see some Scandentia featured.
My brain says this is a springhare (Pedetes) so I am going with what my brain says and anyway I really like this display.
When I saw this I thought, “That’s a nice… rodent thingy.” And so “rodent thing” it shall be labelled here. Enjoy the rodent thingy. Some serious taxidermy-fu in action.
Moonrats– now there’s something you seldom see a full display of. Well done!
That’s part I of this sneak peek at the evolving exhibits- I will put up a part II once the upper floor exhibits open. I highly encourage a visit!
Back in November 2016 I got an exciting email from colleague Dr. Richard Thomas, who was building a team of experts for a proposed documentary on Jumbo the elephant; the famed proboscidean of the Victorian era (and arguably most famous elephant of all time, first international celebrity animal, etc.). I knew him from social media and from our mutual interests in chicken anatomy and evolution. And that exciting email, for once, worked out! Over coming months I chatted with the film producers and they could see a place for me in the programme, contributing my expertise in elephant (postcranial) anatomy, locomotion, health/welfare etc. Lo and behold, in May 2017 I met Sir David Attenborough at Heathrow and we flew out to New York City to film with the skeleton at the American Museum of Natural History. And to cap it off, I got to meet another childhood science communication hero: Professor David Suzuki of CBC’s “The Nature of Things“– my adrenaline levels were sky high!
Brooklyn neighborhood by our hotel. Lots to do!
The show has aired in the UK and is coming very soon to Canada and the world (details below). Here’s my part of the story.
Stomach-Churning Rating: 3/10– bad bones but no blood.
We filmed from 15-19 May 2017 at the AMNH’s warehouse of mammalian skeletal remains, which is housed deep in the Brooklyn Army Terminal; a picturesque site in and of itself. And it is a site with a lot of history— WWI and II, Elvis and more.
It was a hectic week of the usual documentary stuff: repeat the same lines and motions again and again from different angles and with different paces and intonations (I cannot help in these cases but think about the Simpsons “Fallout Boy” episode), from ~9am-5pm, with plenty of downtime watching setup or other bits being filmed. I’m used to all that. But having the time to peer around the collection and chat to Richard and colleague Dr. Holly Miller (handling the tissue isotopes side of the story) about Jumbo’s skeleton was a lot of fun during downtime and filming itself. Not to mention the utter joy of studying one of the most famous museum specimens ever, and an animal widely held to be one of the largest of its kind, with much mystery surrounding its history despite its fame. (Wikipedia does a fair job of summarizing some of this)
Here are some photos to tell the story:
Photo of the team, courtesy of Infield Fly Productions (CBC production, “Jumbo: The Life of An Elephant Superstar”.
The Brooklyn Army Terminal, with a view of the harbour beyond.
Inside the terminal: old army staging area and an evocative wooden Liberty/tank artwork.
Army terminal cat. Shipping still comes through the terminal so I guess there are plenty of rats and handouts from cat-lovers to keep it going. I miss our cats when I travel so this moment was appreciated.
Whale skulls and other specimens inside the AMNH warehouse.
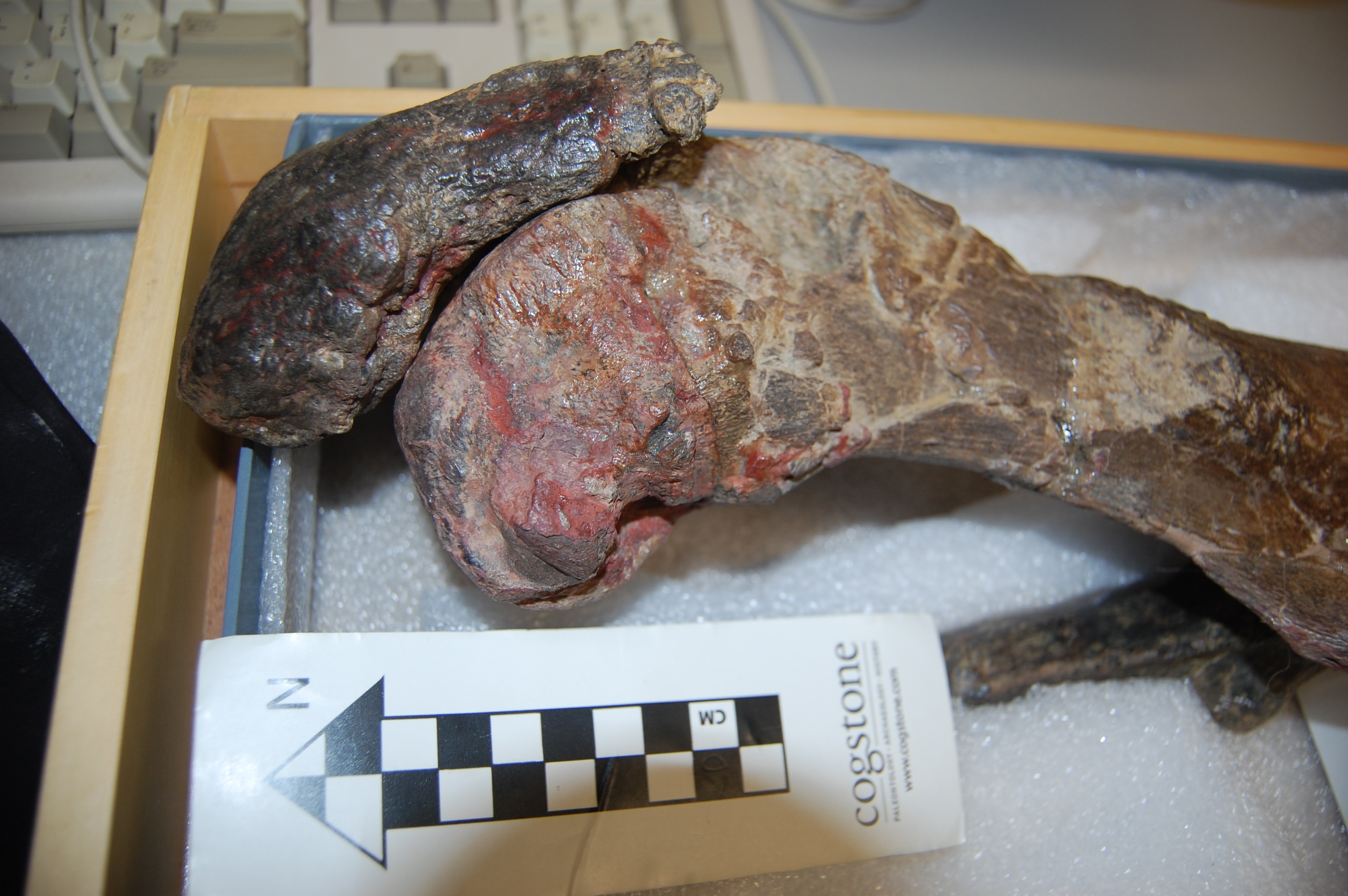
First view of Jumbo’s remains.
Photo opp with Sir David.
Photo opp with Prof Suzuki.
That’s the setup. I’ve done ~15 other documentary episodes/shows but this was like nothing else– simply an awesome experience.
Now the delivery: we set to studying those bones. We’d seen photos before, and Henry Fairfield Osborn had illustrated the specimen as his type of “Elephas africanus rothschildi” (Sudanese elephant; no longer valid but those were different times– it’s now just a nicely preserved Loxodonta africana africana), so we knew some of what to expect.
Looking at Osborn’s classic monograph. Oddly he didn’t address the GLARING MASSIVE PROBLEMS WITH THE TEETH!
Skull with terrible tooth pathologies– and let’s play spot Mumbo, my daughter’s toy elephant! He might even appear in some TV footage!
We had noted some serious issues with some bones (pathologies). I won’t spoil the message here but will show some images. I know some experts have voiced issues with how the tooth pathologies/growth were explained in some footage but I can’t address that here; it’s not my expertise. The important point to me is that the teeth are incredibly messed up and that can easily be linked to bad diet and other management/health issues, as the documentary explains.
Jumbo’s torso in left side view. Glorious preservation.
Right forelimb, showing that the “growth plates” (epiphyses”) were not all fused, consistent with Jumbo still growing– as expected for an African male elephant in his 20’s.
Right elbow with some pathologies consistent with degenerative joint disease.
Surprisingly, Jumbo’s feet were not in nasty condition in terms of pathologies. I’d expected to see that. They’d been painted and drilled for mounting, but were not riddled with arthritic changes that I could see.
Strange bony plaque on the left pelvis (hip) region; something I’d never seen before in any elephant (and I’ve seen many). Why? The programme offers a reasonable explanation.
Jumbo’s right hip, with bad erosion of the bone and thus presumably the overlying cartilage. Ouch!
Strange extra prong on one right rib in Jumbo- we didn’t figure that out. It could conceivably be natural variation.
So, poor Jumbo suffered some jumbo-sized problems, and in complex ways. That’s just scratching the surface of what his skeleton tells us, and there’s plenty more in the show plus plenty more we can say later– there’s real science that came out of this programme! I was surprised to find how little had been stated anywhere in the scientific literature about Jumbo’s pathologies.
Sad as Jumbo’s skeletal story is, the broader story of his life and death is sadder still. For purposes of time I don’t think any of the three versions of the show will get to delve into how Jumbo’s mother may have been slashed to death by a broadsword, as the story below describes was the ancient practice:
I’d hate to be “so pestered by a popinjay”, too.
Adding insult to injury, we can reflect on how Jumbo was taken from the Sudan to the east (across the Suez), then on boat to Italy and then overground to Paris, where he lived for a little while until the zoological garden sold him to London. Luckily Jumbo avoided becoming a meal to starving Parisians during the Prussian siege of 1870-1. So he did not become elephant consommé like some of his co-captives did. The more one learns about Jumbo’s life and the life of elephants in captivity in the 1800s, the more harrowing the tale becomes.
Jumbo is THE celebrity elephant. His name has come to mean ‘big’ and ‘bombastic’, from applications to jumbo jets to hot dogs and other (darkly ironic) forms of consumption and extravagance. He has had a jumbo effect on Western culture, but also symbolizes the complex human-elephant relationship, such as the inspiration for “Dumbo’s” own sad story. We love elephants but our fascination with them can also be their undoing, such as poaching for the ivory trade or mistreatment in captivity. Jumbo’s story writ large is also the story of elephants, and our story to learn from. If anything comes out of my participation in the Jumbo documentary for the public’s benefit, I hope it is increased empathy for how we interact with elephants. They are like us in many ways (maybe over-emphasized with anthropomorphism in many accounts), but also unlike us (maybe even unfathomable) in not only their size and anatomy but also in aspects of their prodigious intellect, emotions and social structure. Elephants aren’t just jumbo spectacles. They are jumbo responsibilities for humans now that we dominate the planet so much.
Want to catch a version of the Jumbo show? I’ll try to keep this list up to date:
BBC iplayer now: https://www.bbc.co.uk/iplayer/episode/b09jcxrj/attenborough-and-the-giant-elephant
An epiphysean Sispyhean task today: solve this mystery that has been bothering me for >15 years. It’s about bird knees. Read on.
Stomach-Churning Rating: 1/10- bones and brief words. Nothing to worry about.
Here is an ostrich. I was interviewing undergrads the other day and looked up to see it, then thought something like: “Oh yeah, that little bit of bone really bothers me. I cannot figure it out.” What little bit of bone?
Right leg, side view, ostrich…
This little bit of bone. Zooming in on that ostrich’s knee:
Who am I? (femur above; tibiotarsus below; “PTE” is the crest of bone with the white arrow on it)
The little bit of bone is not talked about much in the scientific literature on bird knees. But we know it’s there and it is part of the composite bone called the tibiotarsus (ancestral tibia, this bit of bone, and the proximal tarsal [ankle] bones on the other end; the astragalus and calcaneum of earlier dinosaurs).
What is it? We call it something like the proximal tibial epiphysis (PTE for short, here). An epiphysis is an end of a bone that fuses up with the shaft during growth, around the time of skeletal maturity; ultimately ending longitudinal (length-wise) growth of that bone. Mammals almost ubiquitously have them. So do lizards and tuataras. And some fossil relatives. Not much else– except birds, in this particular region (the two ends of the tibiotarsus; also in the foot region; the tarsometatarsus; which also has its share of mysteries such as the hypotarsus; I won’t go there today). You can see the PTE in mostly cartilaginous form if you take apart a chicken drumstick.
This PTE, like other well-behaving epiphyses, fuses with the tibiotarsus in mature birds, forming one bone. But the young ostrich’s knee above shows the PTE nicely; and other living birds show more or less the same thing.
It begs for explanations. I’ve talked about it in a few of my papers. But I’ve always punted on what it really means– does it have anything to do with the patella (they appear at similar times in evolution; we know that much, roughly)? Where does it come from, developmentally? (we sort of know that but more work is needed in different species and in high resolution) When did it evolve? What does it tell us? Why is it there in living birds and almost no other extinct birds/other dinosaurs? Does it have anything to do with why birds, during their evolution, seem to gradually increase the fusion of skeletal elements or ossify new ones (tendons, kneecaps, etc)? Why here and not in the femur or several other long bones of birds? How much do these PTEs vary between (or within) bird species?
This is the challenge in the post’s title. I present to you: solve this puzzle. Developmentally, biomechanically, evolutionarily, genetically, whatever– why does this PTE happen? There are hints– e.g. this paper proposes why growth rates of long bones favour the formation of “secondary centres of ossification” like this. But I’m unable to satisfy myself with any solutions I can find. Maybe you can complete The Bird Knee Challenge?
Have a go at it in the Comments below! There are plenty of papers or even a grant or something involved in sorting out this single mystery; one of the many basic mysteries about animal anatomy.
As 2017 approaches its end, there have been a few papers I’ve been involved in that I thought I’d point out here while I have time. Our DAWNDINOS project has been taking up much of that time and you’ll see much more of that project’s work in 2018, but we just published our first paper from it! And since the other two recent papers involve a similar theme of muscles, appendages and computer models of biomechanics, they’ll feature here too.
Stomach-Churning Rating: 0/10; computer models and other abstractions.
Mussaurus patagonicus was an early sauropodomorph dinosaur from Argentina, and is now widely accepted to be a very close relative of the true (giant, quadrupedal) sauropods. Here is John Conway’s great reconstruction of it:
We have been working with Alejandro Otero and Diego Pol on Mussaurus for many years now, starting with Royal Society International Exchange funds and now supported by my ERC grant “DAWNDINOS”. It features in our grant because it is a decent example of a large sauropodomorph that was probably still bipedal and lived near the Triassic-Jurassic transition (~215mya).
In our new study, we applied one of my team’s typical methods, 3D musculoskeletal modelling, to an adult Mussaurus’s forelimbs. This is a change of topic from the hindlimbs that I’ve myopically focused on before with Tyrannosaurus and Velociraptor [in an obscure paper that I should never have published in a book! pdf link], among other critters my team has tackled (mouse, elephant [still to be finished…], ostrich, horse, Ichthyostega… dozens more to come!). But we also modelled the forelimbs of Crocodylus johnstoni (Australian “freshie”) for a key comparison with a living animal whose anatomy we actually knew, rather than reconstructed.
Mussaurus above; Crocodylus below; forelimb models in various views; muscles are red lines.
The methods for this biomechanical modelling are now standard (I learned them from their creator Prof. Scott Delp during my 2001-2003 postdoc at Stanford): scan bones, connect them with joints, add muscle paths around them, and then use the models to estimate joint ranges of motion and muscle moment arms (leverage) around joints. I have some mixed feelings about developing this approach in our 2005 paper that is now widely used by the few teams that study appendicular function in extinct animals. As a recent review paper noted and I’ve always cautioned, it has a lot of assumptions and problems and one must exercise extreme caution in its design and interpretation. Our new Mussaurus paper continues those ruminations, but I think we made some progress, too.
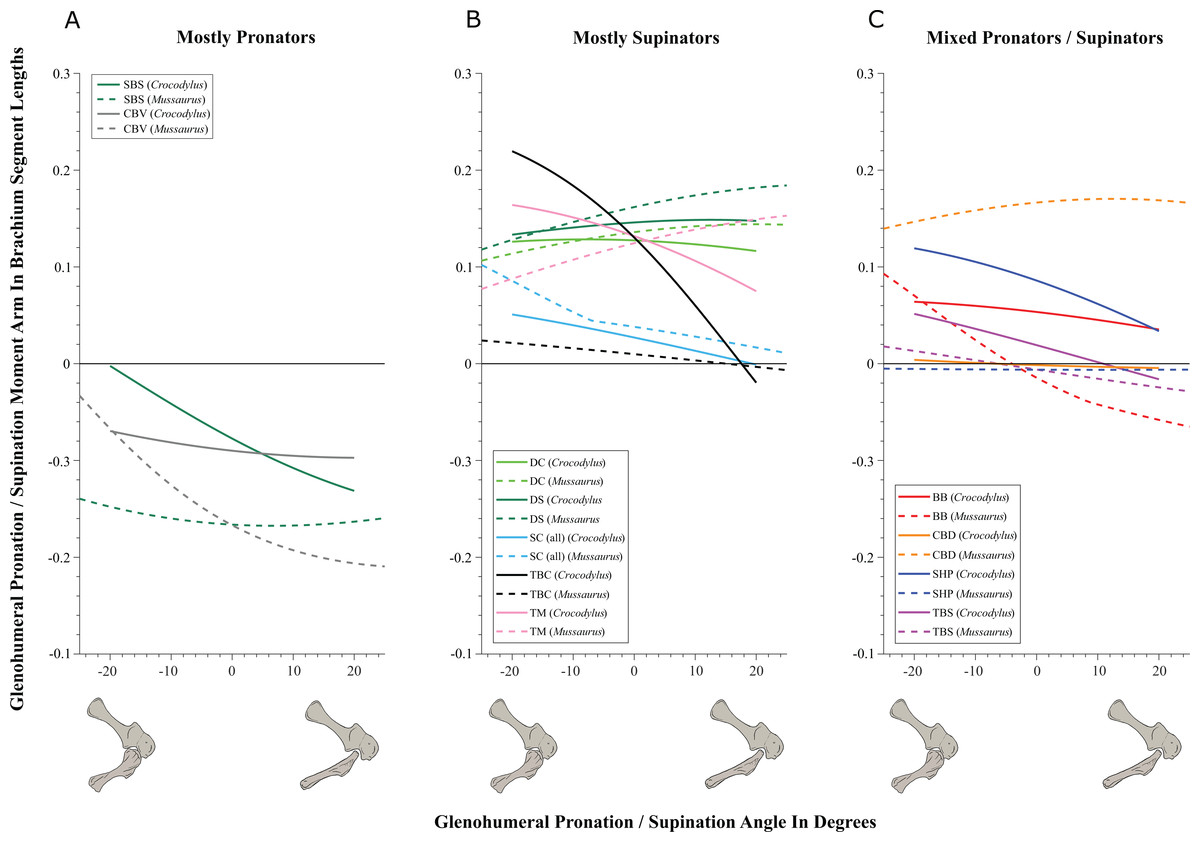
On to the nuts and bolts of the science (it’s a 60 page paper so this summary will omit a lot!): first, we wanted to know how the forelimb joint ranges of motion in Mussaurus compared with those in Crocodylus and whether our model of Mussaurus might be able to be placed in a quadrupedal pose, with the palms at least somewhat flat (“pronated”) on the ground. Even considering missing joint cartilage, this didn’t seem very plausible in Mussaurus unless one allowed the whole forearm to rotate around its long axis from the elbow joint, which is very speculative—but not impossible in Crocodylus, either. Furthermore, the model didn’t seem to have forelimbs fully adapted yet for a more graviportal, columnar posture. Here’s what the model’s mobility was like:
So Mussaurus, like other early sauropodomorphs such as Plateosaurus, probably wasn’t quadrupedal, and thus quadrupedalism must have evolved very close to in the Sauropoda common ancestor.
Second, we compared the muscle moment arms (individual 3D “muscleactions” for short) in different poses for all of the main forelimb muscles that extend (in various ways and extents) from the pectoral girdle to the thumb, for both animals, to see how muscle actions might differ in Crocodylus (which would be closer to the ancestral state) and Mussaurus. Did muscles transform their actions in relation to bipedalism (or reversal to quadrupedalism) in the latter? Well, it’s complicated but there are a lot of similarities and differences in how the muscles might have functioned; probably reflecting evolutionary ancestry and specialization. What I found most surprising about our results was that the forelimbs didn’t have muscles well-positioned to pronate the forearm/hand, and thus musculoskeletal modelling of those muscles reinforced the conclusions from the joints that quadrupedal locomotion was unlikely. I think that result is fairly robust to the uncertainties, but we’ll see in future work.
You like moment arms? We got moment arms! 15 figures of them, like this! And tables and explanatory text and comparisons with human data and, well, lots!
If you’re really a myology geek, you might find our other conclusions about individual muscle actions to be interesting—e.g. the scapulohumeralis seems to have been a shoulder pronator in Crocodylus vs. supinator in Mussaurus, owing to differences in humeral shape (specialization present in Mussaurus; which maybe originated in early dinosaurs?). Contrastingly, the deltoid muscles acted in the same basic way in both species; presumed to reflect evolutionary conservation. And muuuuuuch more!
Do you want to know more? You can play with our models (it takes some work in OpenSim free software but it’s do-able) by downloading them (Crocodylus; Mussaurus; also available: Tyrannosaurus, Velociraptor!). And there will be MUCH more about Mussaurus coming soon. What is awesome about this dinosaur is that we have essentially complete skeletons from tiny hatchlings (the “mouse lizard” etymology) to ~1 year old juveniles to >1000kg adults. So we can do more than arm-wave about forelimbs!
But that’s not all. Last week we published our third paper on mouse hindlimb biomechanics, using musculoskeletal modelling as well. This one was a collaboration that arose from past PhD student James Charles’s thesis: his model has been in much demand from mouse researchers, and in this case we were invited by University of Virginia biomechanical engineers to join them in using this model to test how muscle fibres (the truly muscle-y, contractile parts of “muscle-tendon units”) change length in walking mice vs. humans. It was a pleasure to re-unite in coauthorship with Prof. Silvia Blemker, who was a coauthor on that 2005 T. rex hindlimb modelling paper which set me on my current dark path.
Mouse and human legs in right side view, going through walking cycles in simulations. Too small? Click to embiggen.
We found that, because mice move their hindlimb joints through smaller arcs than humans do during walking and because human muscles have large moment arms, the hindlimb muscles of humans change length more—mouse muscles change length only about 48% of the amount that typical leg muscles do in humans! This is cool not only from an evolutionary (mouse muscles are probably closer to the ancestral mammalian state) and scaling (smaller animals may use less muscle excursions, to a point, in comparable gaits?) perspective, but it also has clinical relevance.
Simulated stride for mouse and human; with muscles either almost inactive (Act=0.05) or fully active (Act=1). Red curve goes through much bigger excursions (along y-axis) than blue curve), so humans should use bigger % of their muscle fibre lengths in walking. Too small? Click to embiggen.
My coauthors study muscular dystrophy and similar diseases that can involve muscle stiffness and similar biomechanical or neural control problems. Mice are often used as “models” (both in the sense of analogues/study systems for animal trials in developing treatments, and in the sense of computational abstractions) for human diseases. But because mouse muscles don’t work the same as human muscles, especially in regards to length changes in walking, there are concerns that overreliance on mice as human models might cause erroneous conclusions about what treatments work best to reduce muscle stiffness (or response to muscle stretching that causes progressive damage), for example. Thus either mouse model studies need some rethinking sometimes, or other models such as canines might be more effective. Regardless, it was exciting to be involved in a study that seems to deliver the goods on translating basic science to clinical relevance.
Muscle-by-muscle data; most mouse muscles go through smaller excursions; a few go through greater; some are the same as humans’.
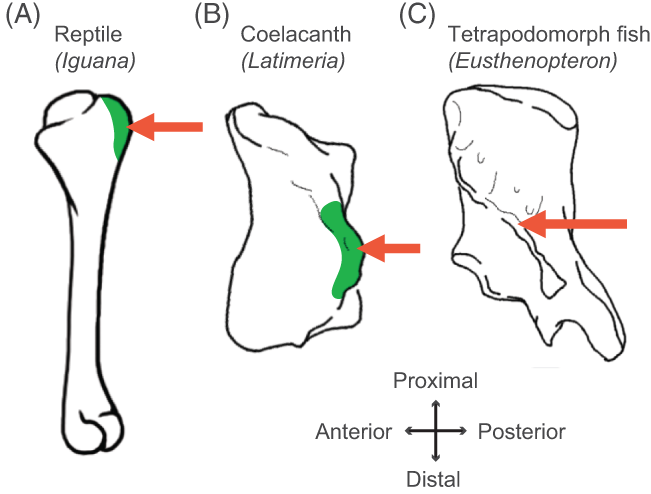
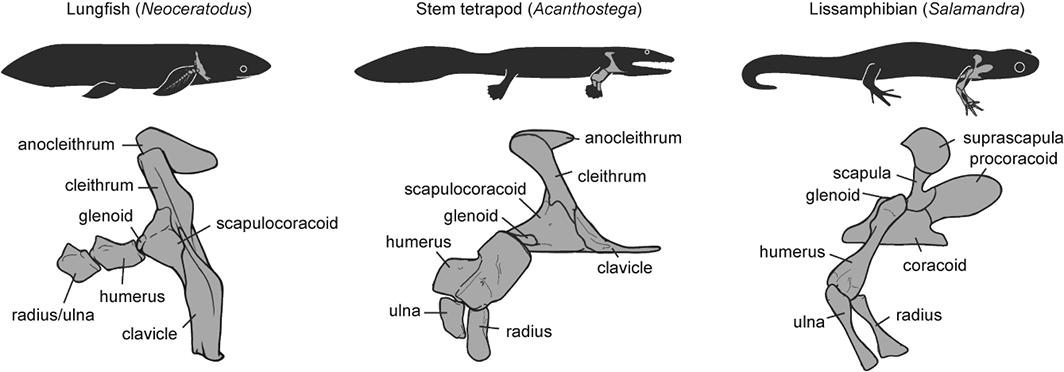
Finally, a third recent paper of ours was led by Julia Molnar and Stephanie Pierce (of prior RVC “Team Tetrapod” affiliation), with myself and Rui Diogo. This study tied together a bunch of disparate research strands of our different teams, including musculature and its homologies, the early tetrapod fossil record, muscle reconstruction in fossils, and biomechanics. And again the focus was on forelimbs, or front-appendages anyway; but turning back the clock to the very early history of fishes, especially lobe-finned forms, and trying to piece together how the few pectoral fin muscles of those fish evolved into the many forelimb muscles of true tetrapods from >400mya to much more recent times.
Humerus in ventral view, showing muscle attachments. Extent (green) is unknown in the fossil but the muscle position is clear (arrow).
We considered the homologies for those muscles in extant forms, hypothesized by Diogo, Molnar et al., in light of the fossil record that reveals where those muscles attach(ed), using that reciprocal illumination to reconstruct how forelimb musculature evolved. This parallels almost-as-ancient (well, year 2000) work that I’d done in my PhD on reconstructing hindlimb muscle evolution in early reptiles/archosaurs/dinosaurs/birds. Along the way, we could reconstruct estimates of pectoral muscles in various representative extinct tetrapod(omorph)s.
Disparity of skeletal pectoral appendages to work with from lobe-fins to tetrapods.
Again, it’s a lengthy, detailed study (31 pages) but designed as a review and meta-analysis that introduces readers to the data and ideas and then builds on them in new ways. I feel that this was a synthesis that was badly needed to tie together disparate observations and speculations on what the many, many obvious bumps, squiggles, crests and tuberosities on fossil tetrapods/cousins “mean” in terms of soft tissues. The figures here tell the basic story; Julia, as usual, rocked it with some lovely scientific illustration! Short message: the large number of pectoral limb muscles in living tetrapods probably didn’t evolve until limbs with digits evolved, but that number might go back to the common ancestor of all tetrapods, rather than more recently. BUT there are strong hints that earlier tetrapodomorph “fishapods” had some of those novel muscles already, so it was a more stepwise/gradual pattern of evolution than a simple punctuated event or two.
Colour maps of reconstructed right fin/limb muscles in tetrapodomorph sarcopterygian (~”fishapod”) and tetrapod most recent common ancestors. Some are less ambiguous than others.
That study opens the way to do proper biomechanical studies (like the Mussaurus study) of muscle actions, functions… even locomotor dynamics (like the mouse study)– and ooh, I’ve now tied all three studies together, tidily wrapped up with a scientific bow! There you have it. I’m looking forward to sharing more new science in 2018. We have some big, big plans!
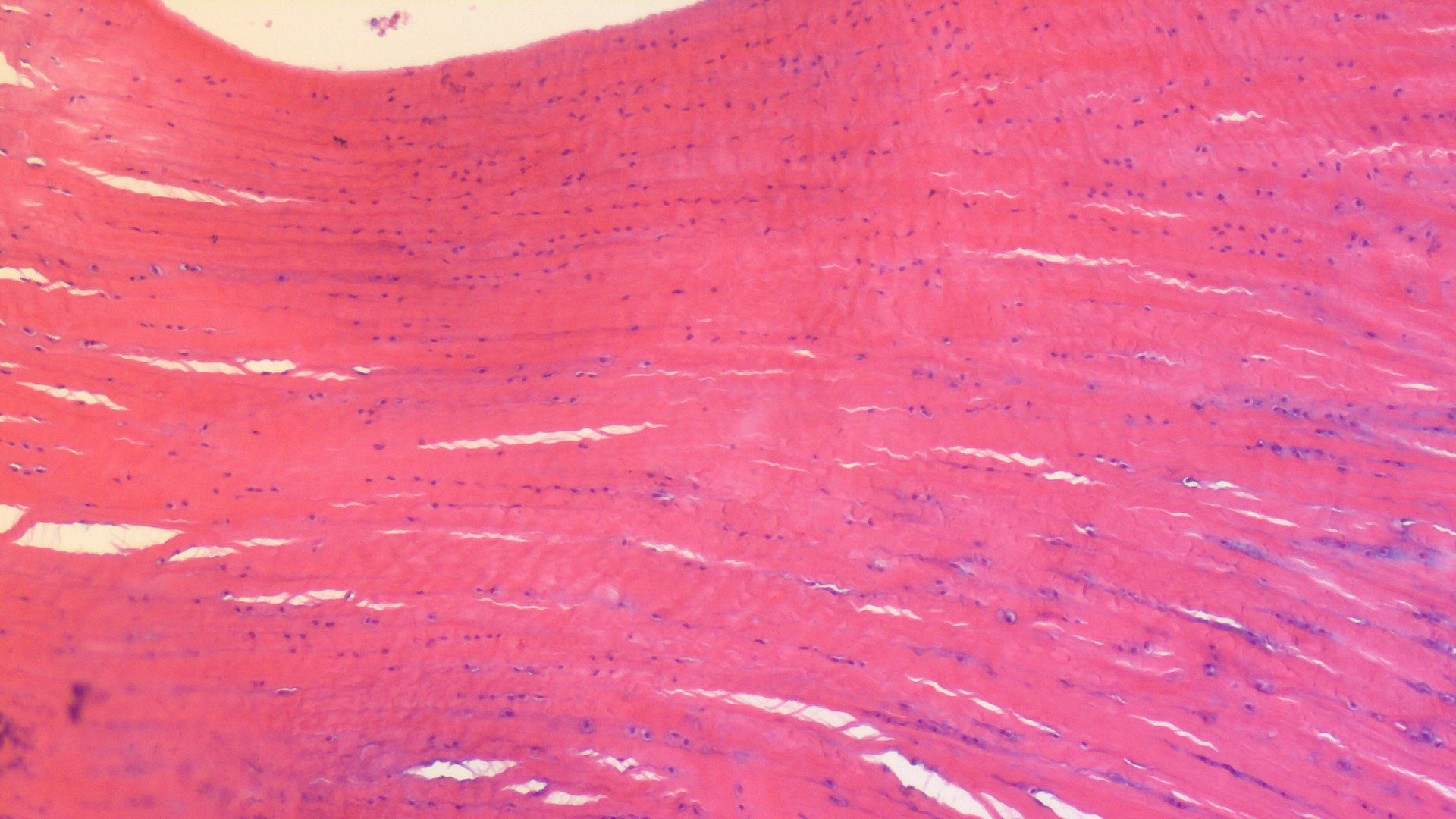
This week we conducted wallaby leg dissections for a study of the kneecaps of marsupials (pouched mammals). Placental (non-pouched) mammals like us almost all have bony kneecaps but many marsupials do not. Kneecaps do important things, acting like gears around the knee joints (e.g. this old post), and yet it is unclear why some marsupials have lost, kept or even re-evolved them as bones. So we’re investigating that and already noticed that one of our wallabies has bony kneecap(s) whereas the other doesn’t, so we’re checking out why and taking tissue samples to do histology (sectioning for microscopic imaging of tissue composition and structure) on so we can see what the knee tendon/kneecap tissues are made of. Some marsupials turn their kneecaps into fibrocartilage rather than bone or tendon and that can be impossible to identify without histology.
The wallabies are small, about 20lbs or so and just take a day or so. Like a turkey. And it’s Thanksgiving today, so here I am with a post about thawing specimens for science, rather than for food. Maybe the title will make sense now.
Stomach-Churning Rating: 7/10; thawed wallaby bits from the get-go.
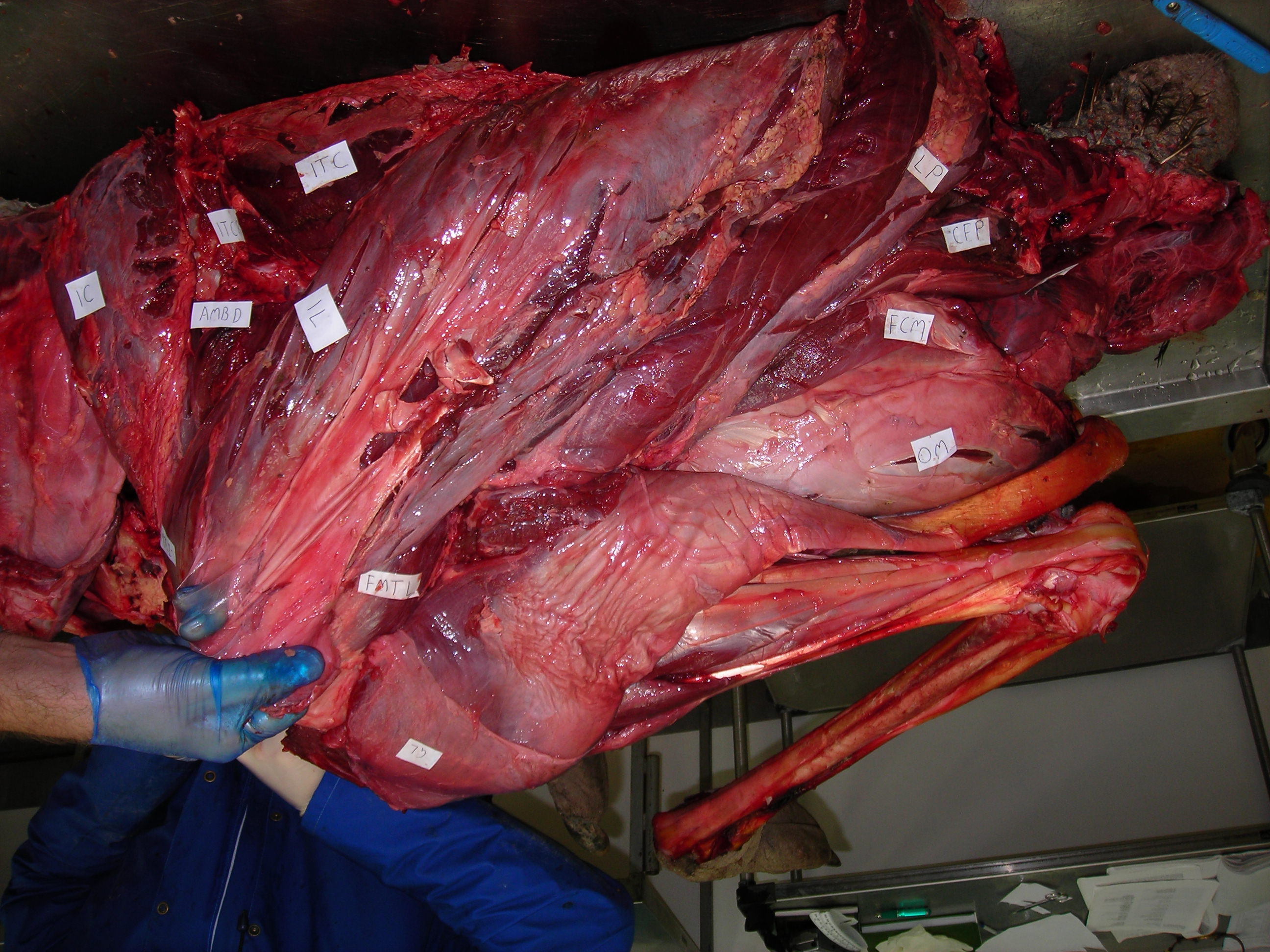
Thawed lower leg and foot of wallaby. The stickers are for an old study that would take too long to explain…
This post was directly inspired by journalist Jason Bittel’s inquiry to me about my tweet on the wallaby thawing; he wondered if there might be a fun story linking thawing-for-science with thawing-for-Thanksgiving. Some highfalutin editors didn’t agree, so no printed/online story came of this, but I am not so highfalutin, hence this blog post.
Thawed wallaby forelimbs. I’m also looking into the “false thumbs” that some marsupials have (“sixth fingers”), much as elephants and other mammals may have.
Thawing is second nature for our lab’s team; we do it all the time. Avid readers will be unsurprised to learn that just about everything I’ve worked on has been frozen at some time, and thus has been thawed out at some time(s). Normally we don’t freeze if we need live tissue or undistorted tissue, e.g. to measure physiology or very fine microstructure– freezing disrupts all of that. We would instead use physiological saline solution or else a preservative like formalin. And you can only freeze and then thaw a specimen for two times or so before it becomes too useless even for anatomical study.
A small specimen like this salamander can be thawed out simply by running it under warm water for a little while or leaving it out for an hour.
We just leave specimens in a cart, or on a table or sometimes in a cold-room shelving area, for slower thawing. Space heaters tend to overdo things. We don’t do any rough calculation from some sort of thermodynamic first principles of time-to-thaw vs. specimen size (I wish we were that smart!); just seat-of-pants guessing and checking (yes, poking specimens to check their thawedness is a method of choice). Cutting things in half along the way, or skinning them, may be used to accelerate the thawing process. But it’s about as unscientific a method as we use.
The hardest specimens to thaw of course have been the largest specimens. Elephant legs can be >2 metres long and hundreds of kilograms (especially when frozen). A week at room temperature tends to work OK for getting them to a dissectable state. One has to balance the outer deterioration with the inner frigidness. We’re not so concerned about microbe growth in most cases, as one would be with a thawing turkey, and not at all about consumption. We just want to be able to dissect it and make observations, mostly via eyeballing the specimens as we dissect them,
Left hindfoot of an Asian elephant. Still frozen; this was bandthawed- I mean bandsawed- to see its internal anatomy nice and clearly. You may see this specimen again somewhere else– stay tuned! 🙂
Moisture and fluids can be a challenge: generally the rooms we thaw in are low humidity so moisture may not be an issue once the ice melts away, and we have drains nearby. We try to remove ice first or have towels to wipe/soak fluids up as thawing progresses. But if a specimen is sitting in a cart or storage bag with too much ice early on, that can thaw first and then turn the specimen into a nasty slurry of the stuff you’re interested in and the less desirable muck. So we try to avoid that.
De-thawing too early is bad. The smell gets progressively worse– and once the interior of the specimen is thawed enough, then bacteria get in there and the interior becomes a brewing ground for heat production (rather than remaining a cooler region), which accelerates decay, so we don’t want that. We have to check on thawing specimens regularly and move them to cooler storage areas, or begin dissection earlier, if the decay process is noticeably getting excessive.
Any insulation affects thawing time- so scales, feathers, thick skin, shells, fat (for a short while until it decays), and other layers will slow thawing—and may keep heat inside, if there begins to be thawing of the core. So sometimes you open up a specimen that seems dry and clean on the outside and the inside is unpleasant. But with experience that is not hard to avoid.
Thawed wallaby patella prepared for histology.
The foulest specimen I’ve thawed by far was a monitor lizard… it was shipped to me in California from Arizona when I was a PhD student. This was in August’s heat and the box of the big lizard sat thawing at the post office for 2 weeks before they contacted me and asked why a smelly box was bleeding. I came and got it and brought it back to our department but the smell was so bad it set off our building health & safety person’s alarm bells (sorry, David!) and they emailed around a “toxic alert” warning, until I bashfully made it clear that my lizard was the cause, not some toxic chemical. I got in some trouble and was very ashamed. But we put the specimen into a big tank of brine solution and the smell was reduced—the specimen may well still be preserved there 20 years later; I do wonder! Anyway, that experience was so horrendous – and I have a strong stomach—that I regularly recall it and seek to avoid a repeat. It was the most disgusting thing I’ve ever experienced. I do not recommend it.
What we tend to want to get from thawed specimens is: (1) descriptive anatomy (what connects where), and maybe (2) quantitative measurements (laborious metrics of “muscle architecture”– how much does each muscle weigh, how long is it, etc; over and over again for many muscles…). These data not only serve to tell us what makes animals different (and how this evolved) but also the data are used to test questions such as how animals work. In the case of things like wallabies, ultimately we’d love to know what their kneecaps do if they are bony or not; what difference does it make and why might there be differences? We’d spotted one wallaby already that seemed to have a bony kneecap on one leg, and a non-bony one on the other leg, so that asymmetry got us excited.
What’s surprising to learn about thawing animals for science? Well, my first thought is that it’s beautiful. I don’t tend to think of it as gross. I’ve rhapsodized about this before. Animals are wonderful inside and out, and I regularly pause during a dissection to marvel at how amazing the anatomical specializations of animals are. Simple details- shapes, colours, configurations- can be gorgeous. (Often the blood is minimal, drained out early, so that doesn’t detract from or hide the detailed imagery) The gentle yet complex path of a tendon around a joint can yield profound visual enchantment in its elegance. This is all the more true once one ponders how these complex structures evolved, and how much diversity of form and function is out there to study—and how little we know about it! We still don’t know well how to fix many problems humans have with their anatomy, and that’s orders of magnitude worst for most animals, because we don’t understand how anatomy works, or even what the anatomy is like in some cases. So that keeps me busy discovering things. Every specimen is different with surprising little variations, or big ones—sometimes there is one muscle, sometimes it is clearly divided into two muscles, in the same species or even the left vs. right legs. I love seeing those intricacies and wondering about them.
Thawed wallaby shank sliced open to show lovely digital flexors and gastrocnemius muscles. So many questions are raised by this!
If you’re thawing for Thanksgiving, or thawing for science, or thawing out family relations during a gathering, or thawing yourself out from the winter’s cold– my best wishes to you! May we all enjoy what we thaw.
Putting my morphologist hat back on today, I had an opportunity to dissect an Elegant-crested tinamou (Eudromia elegans) for the second time in my life. The last time was during my PhD work ~20 years ago. In today’s dissection I was struck by another reminder of how studying anatomy is a lifelong learning experience and sometimes it’s really fun and amazing even when it’s stinky.
Tinamou foot. I did know that tinamous don’t have a hallux; big “perching toe” (1st/”big toe” in us); true of ratites/palaeognaths more generally. Unlike a chicken or many other birds. Just the three main toes (2, 3 and 4) are here.
Stomach-Churning Rating: 7/10; you gotta have guts to learn about intestine-churning stuff.
Tinamous are neat little partridge-like ground birds but they are not close cousins of partridges or guineafowl at all. Their closest cousins are other ratites/palaeognaths such as ostriches and kiwis. And hence they are found in South America, especially Patagonia in Argentina. I’ve seen them there, much to my enjoyment.
Said tinamou.
What struck me today was that, as I delved into the digestive system of this bird, I saw features that were unfamiliar to me even after having dissected many species of birds from many lineages. The intestinal region was very lumpy, with little bud-like pockets full of dense droppings. Furthermore, on separating the tubes of the small and large intestines I realized that most of the intestinal volume itself was caecum (normally a modest side-pocket near the juncture of the small and large intestines). Indeed, that caecum was caeca (plural): it had two massive horns; it was a double-caecum, feeding back into the short rectum and cloaca. Birds have variable caeca and it is typical to see subdivision into two parts, but I’d never seen it to this degree.
Oh why not, here’s the gizzard/stomach showing its grinding pebbles and bits of food, plus the strong outer muscle layers (pink) for driving that grinding. Small intestine heads toward the bottom of the image. Yes, we do need a better dissection light…
I had to question my anatomical knowledge at this point, wondering if I was identifying things incorrectly—did I really screw up somehow and these were other organs, like giant ovaries? But no, they were clearly full of faecal matter; they were digestive organs. I finished the dissection, still puzzled, and hit the literature. Right away, Google-Scholaring for “tinamou caecum” I found the answer, here (free pdf link):
“at least one species (Elegant Crested Tinamou, [Eudromia elegans]), the ceca contain multiple sacculations, resulting in structures that look much like two bunches of fused grapes.”
The caeca in question.
OK buddy, those are the little lumpy buds I saw. Bunches of grapes—exactly.
And later:
“The paired ceca of the Elegant Crested Tinamou are extraordinary and probably unique within Aves (Fig. 3): long and wide (12.5-13.0 X 2.2- 2.5 cm; Wetmore 1926) and internally honeycombed by many small diverticula. These outpocketings gradually diminish in size and organization from the base to the tip of the organ, apically showing a more spiral form of internal ridges like ratite ceca. Externally, the basal diverticula protrude from the ceca as pointed lobes, gradually becoming flatter but still clearly apparent toward the organ’ s tip.”
Whoa! I never knew that! So I happened to be dissecting a bird, fairly common in its homeland, that has a really bizarre and singular form of caeca/ceca! That hit my morphologist sweet-spot so I was very pleased and decided to share with you. It is one of those many examples of times when you quickly go from confusion to illumination as a scientist, emerging with a neat fact about animal biology. And journal articles help you get there!
The bare “brood patch” on the back end of the tinamou’s belly; a nicely hotspot for keeping eggs warm. Perhaps for brooding bad puns, too.
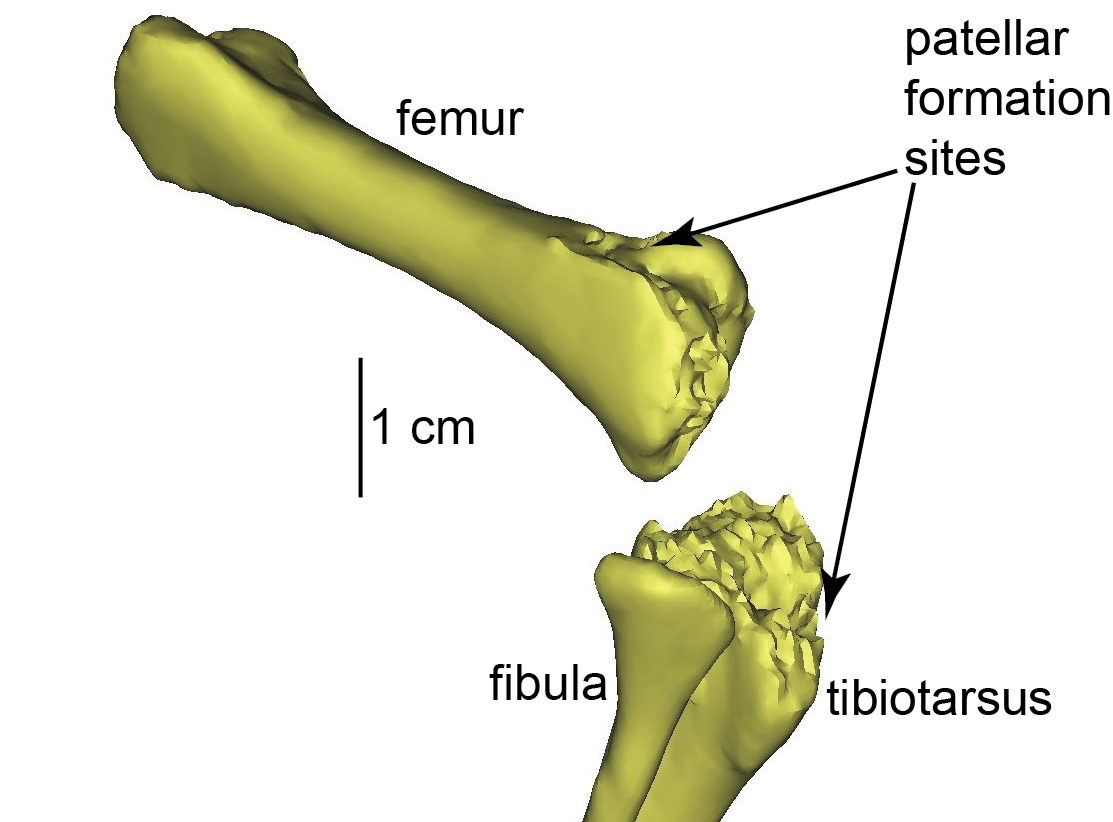
Uh oh, a “why?” question in biology! There are many potential, and not mutually exclusive, answers to such questions. Ultimately there is a historical, evolutionary answer that underpins it all (“ostriches evolved two kneecaps because…”). But we like ostrich knees and their funky double-kneecaps (patellae; singular = patella) so we wanted to know why they get so funky. One level of addressing that question is more like a “how?” they have them. So we started there, with what on the surface is a simple analysis. And we published that paper this week, with all of the supporting data (CT, MRI, FEA).
Stomach-Churning Rating: 6/10 because there is a gooey image of a real dissection later in the post, not just tidy 3D graphics.
First author Kyle Chadwick was my research technician for 2 years on our sesamoid evolution grant, and we reported earlier on the detailed 3D anatomy of ostrich knees (this was all part of his MRes degree with me, done in parallel with his technician post). Here, in the new paper with Sandra Shefelbine and Andy Pitsillides, we took that 3D anatomy and subjected it to some biomechanical analysis in two main steps.
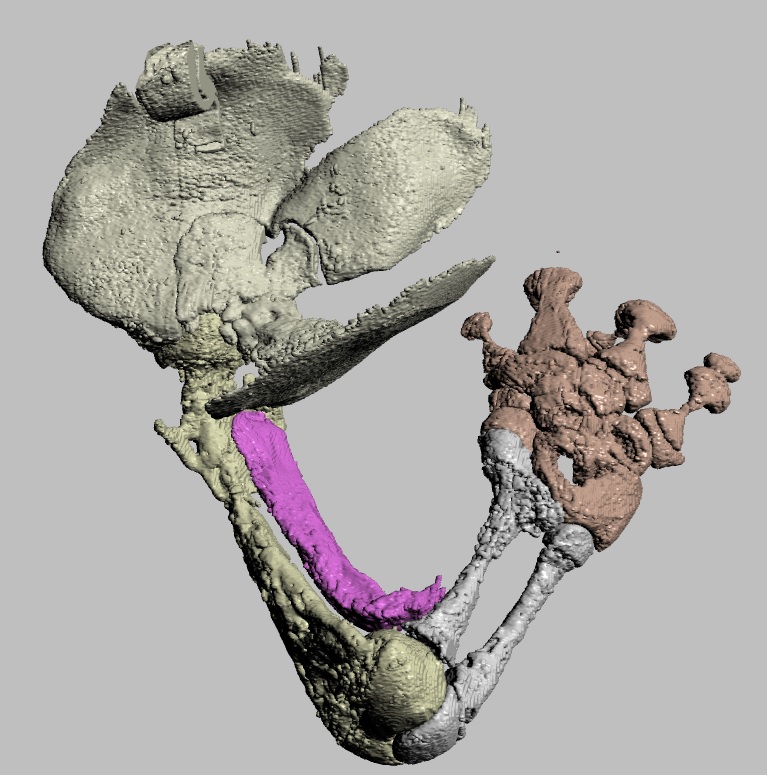
Ostrich (right) knee bones. The patellae are the two knobbly bits in the knee.
First, we used our previous biomechanical simulation data from an adult ostrich (from our paper by Rankin et al.) to estimate the in vivo forces that the knee muscles exert onto the patellar region during moderately large loading in running (not maximal speed running, but “jogging”). That was “just” (Kyle may laugh at the “just”– it wasn’t trivial) taking some vectors out of an existing simulation and adding them into a detailed 3D model. We’ve done similar things before with a horse foot’s bones (and plenty more to come!), but here we had essentially all of the soft tissues, too.
Ostrich knee with muscles as 3D objects.
Second, the 3D model that the muscular forces were applied to was a finite element model: i.e., the original 3D anatomical model broken up into a mesh, whose voxels each had specific properties, such as resistance to shape change under loading in different directions. The response of that model to the loads (a finite element analysis; FEA) gave us details on the stresses (force/area) and strains (deformations from original shape) in each voxel and overall in anatomical regions.
Finite element model setup for our study. If you do FEA, you care about these things. If not, it’s a pretty, sciencey picture.
The great thing about a computer/theoretical model is that you can ask “what if?” and that can help you understand “how?” or even “why?” questions that experiments alone cannot address. Ostriches aren’t born with fully formed bony kneecaps; indeed those patellae seem to mature fairly late in development, perhaps well after hatching. We need to know more about how the patellae form but they clearly end up inside the patellar (knee extensor) tendon that crosses the knee. So we modelled our adult ostrich without bony patellae; just with a homogeneous patellar tendon (using the real anatomy of that tendon with the bony bits replaced by tendon); and subjected it to the loading environment for “jogging”.
The right knee of an ostrich hatchling. The patellae have yet to form; indeed there is little bone around the knee region at all, yet.
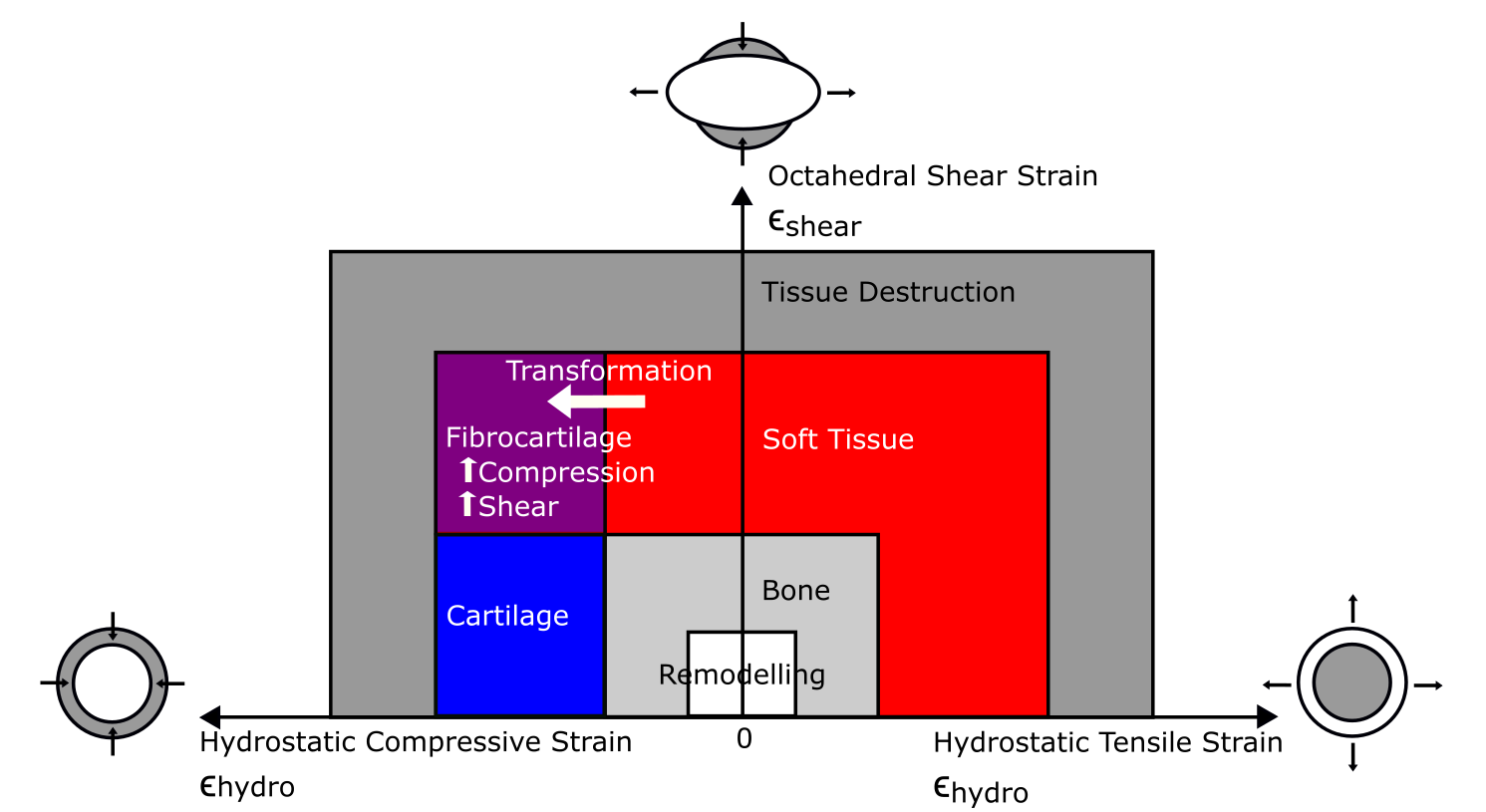
We then inspected our FEA’s results in light of modern theory about how tissues respond to loading regimes. That “mechanobiology” theory, specifically “tissue differentiation”, postulates that tendon will tend to turn into fibrocartilage if it is subjected to high compression (squishing) and shear (pushing). Then, the fibrocartilage might eventually be reworked into bone as it drops the compression and shear levels. So, according to that theory (and all else being equal; also ignoring the complex intermediate states that would happen in reality), the real ostrich’s kneecaps should be located in the same positions where the FEA, under the moderately large loads we applied, predicts the homogeneous tendon to have high compression and shear. But did the real anatomy match the mechanical environment and tissue differentiation theory’s predictions?
Tissue differentiation diagram displaying the theoretical pathways for transformation of tissues. If tendon (red) experiences high shear (going up the y-axis) and high compression (going toward the left), it should turn into fibrocartilage (purple). Transformation into bone (diagonally to the bottom right) would reduce the shear and compression.
Well, sort of. The image below takes some unpacking but you should be able to pick out the red areas on the bottom row where the patellae actually are, and the yellow shaded regions around some of those patellar regions are where the compression and shear regimes are indeed high and overlapping the actual patellar regions. The upper two rows show the levels of compression (or tension; pulling) and shear, but the bottom row gets the point across. It’s not a bad match overall for the first (“real”; common to all living birds) patella, located on top of the upper knee (femur). It’s not a good match overall for the second (unique to ostriches) patella, located below the first one (and attached to the tibia bone).
FEA results! (click to embiggen)
Kyle says, “Being a part of this project was exciting because of the application of engineering concepts to interesting biological (including evolutionary) questions. Also, it never gets old seeing people’s reactions when I tell them I study ostrich knees.”
The study had a lot of nuances and assumptions. We only looked at one instant in slow running and only at one adult ostrich, not at the full development of ostrich anatomy and loading. That’s harder. We started simple. The tissue differentiation theory is used more for fracture healing than for sesamoid bone formation but there’s some reason to suspect that similar mechanisms are at play in both. And there’s much more; if you want the gory details see the paper.
So did we solve why, or how, ostriches have two kneecaps? We felt that the mechanical environment of our FEA was a good theoretical explanation of where the first patella forms. We originally expected the second patella, which evolved more recently and might be more mechanically sensitive as a result, to be a better match than the first one, but it was the opposite. C’est la science!
Enough models, let’s have some reality! I warned you this post would get messy, and here it is. Left leg (skinned) of an ostrich showing the muscles around the knee. The patellar region would be in the gloved hand of the lucky individual shown.
This study, for me, was a fun experience in moving toward more fusion of “evo-devo” and biomechanical analyses, a research goal of mine lately– but there’s still a ways to go with the “how?” and “why?” questions even about ostrich kneecaps.
We felt that the best conclusion supported by our analyses was that, rather than have homogeneous stresses and strains throughout their knee tissues (e.g. the patellar tendon), ostriches have a lot of regional diversity in how those tissues are loaded (in the condition we modelled, which is adequately representative of some athletic exertion). Look at the complex FEA coloured results above again, the top two rows: there are a lot of different shades of compression/tension and shear; not homogeneous strains. That diversity of regional loading sets those tissues up for potential transformation throughout growth and development. And thus ONE of the reasons why ostriches might have two kneecaps is that the heterogeneous loading of their knee tendon favours formation of heterogeneous tissue types.
Another, compatible, explanation is that these different tissues might have consequences for how the muscles, tendon and joint operate in movement behaviours. In due time there will be more about that. In the meantime, enjoy the paper if this post makes you want to know more about the amaaaaaazing knees of ostriches!
A Confuciusornis fossil; not the one from our study but prettier (more complete).
Today almost three years of collaboration come together in a publication that is a fun departure from my normal research, but also makes sense in light of it. Professor Baoyu Jiang from Nanjing University in China has been being working on the taphonomy of the Early Cretaceous Jehol biota from northeastern China (Manchuria) for a while, and he found a lovely Confuciusornis (early bird) fossil; one of thousands of them; from the volcanic pyroclastic flow-based lake deposits there.
Although at first glance the skeletal remains of that fossil are not fabulous compared with some other Confuciusornis, what makes this one lovely is that, on peering at it with multiple microscopic and other imaging techniques, he (and me, and a China-UK collaboration that grew over the years) found striking evidence of very well-preserved fossil soft tissues. Our paper reporting on these findings has gone live in Nature Communications so I can blog about it now.
Reference: Jiang, B., Zhao, T., Regnault, S., Edwards, N.P., Kohn, S.C., Li, Z., Wogelius, R.A., Benton, M., Hutchinson, J.R. 2017. Cellular preservation of musculoskeletal specializations in the Cretaceous bird Confuciusornis. Nature Communications 8:14779. doi: 10.1038/NCOMMS14779
Stomach-Churning Rating: 3/10; gooey, but fossil gooey, except for some colourful, gastrically-tolerable histology of bird tissue.
Front view of the ankle/foot of our specimen.
Back view of the ankle/foot of our specimen.
What has been fun about this collaboration is that, for one, it fits in perfectly with my prior work. Ever since my PhD thesis I’d been wondering about odd bones in the legs of birds, including a very puzzling and very, very neglected bit of bone called the tarsal sesamoid, on the outside of the upper end of the ankle joint. Furthermore, a tunnel of tissue called the tibial cartilage sits next to that sesamoid bone, and then across the ankle joint there is a bony prominence with grooves and tunnels that vary highly among bird species; that is called the hypotarsus. These structures are all known in living birds and, to a degree, in extinct fossil cousins. Our specimen seems to reveal an earlier stage in how these little features of bird ankles originated, which we concluded to be a step along the transition to the more crouched legs that modern birds have.
This study has also challenged me to broaden my horizons as a scientist. Although this was a big collaboration and thus we had several specialists to apply supercharged technological techniques to our fossil, I had to learn something about what all that meant. My kind colleagues helped me learn more about tissue histology, scanning electron microscopy, synchrotron mapping, FTIR and mass spectrometry and more. I won’t go through all of these techniques but there are some pretty pictures sprinkled here and in the paper, and a lot more detail in the paper for those who want the gory techno-detail. Basically we threw the kitchen sink of science at the fossil to crack open some of its secrets, and what we found inside was nifty.
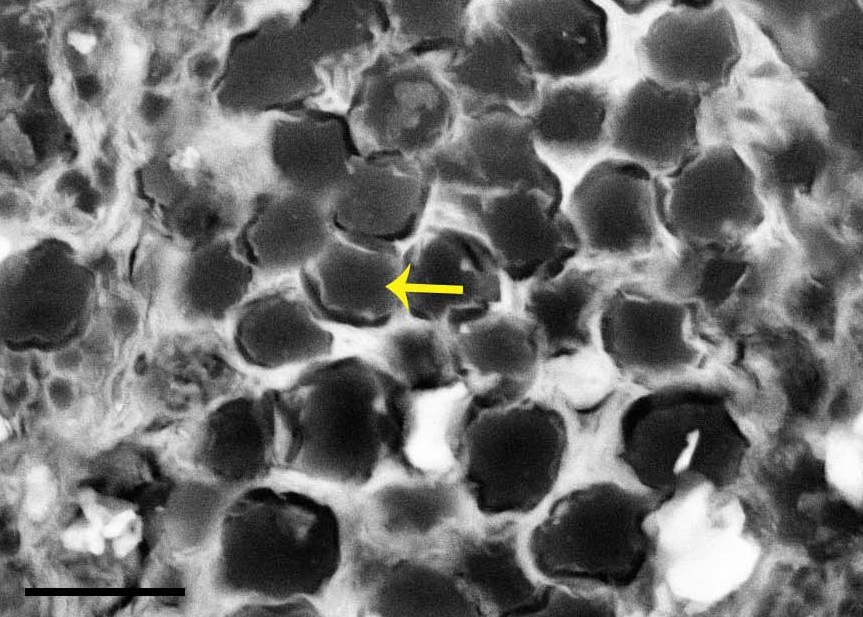
Scanning electron micrograph image of probable tendon or ligament fibres (arrow) in cross-section, from near the ankle joint.
We found preserved cells and other parts of connective tissues including tendons and/or ligaments, fibrocartilage (the tougher kind) and articular cartilage (the softer joint-padding kind). That’s great, although not unique, but the kitchen sink also flushed out even more reductionist data: those tissues included some organic residues, including what appear to be bits of proteins (amino acids); particularly the collagen that makes up tendons.
Fibrocartilage (“fc”) from the ankle joint region.
Hopefully we’re right, and we included as much of the data as we could manage so that others can look at our findings. The specimen is crushed into nearly two dimensions, like all Jehol biota organisms, so its anatomy was hard to interpret but we think we got it right. All of the other kitchen-sinky tools have their own nuances and pitfalls but we did our best with a superb team of experts. We’ve had to wait 125 million years to uncover this specimen and a few more years to find out if we’ve looked at the right way is no greater test of patience.
I thank my coauthors, especially Baoyu Jiang for the kind invitation to participate and the very fun experience of collaborating. I think I’ll remember this study for a long time because, for me, it takes a step beyond just describing Another Case of Jaw-Dropping Fossilization (can you hear the hipsters recounting the excitement and cynicism of the 1990s when this all was dawning? I was there and maybe now I’m one of them). By combining all of those methods we learned new things about the palaeobiology of birds and the evolution of traits within birds. Confuciusornis, not shockingly, had ankles that should have functioned in ways intermediate between those of bog-standard non-avian theropods and modern birds.
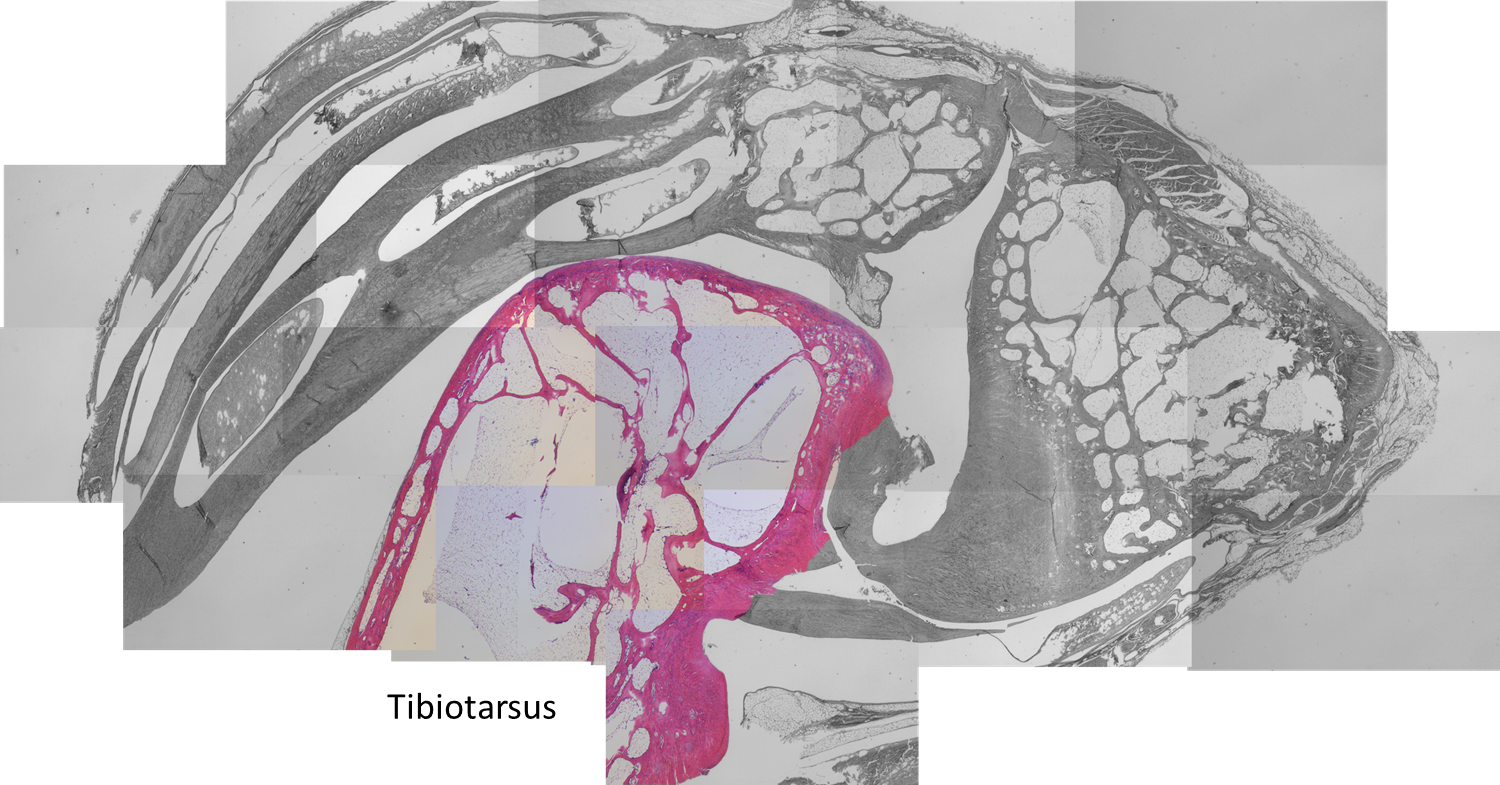
Same anatomical regions in an extant bird as in the main fossil specimen. Left distal tibiotarsus (TT; below) and proximal tarsometatarsus (TMT; above) from an adult helmeted guineafowl (Numida meleagris) after formalin fixation. (from our paper’s Supp Info)
I’m hopeful that more synthesis of molecular/cellular, imaging, biomechanical and other tools (not to mention good old palaeontology and anatomy!) can wash away some more of this mystery. And it was fun to be a part of a study that adds to overwhelming evidence that was heretical ~25 years ago: some hardy biomolecules such as collagen and keratin can survive hundreds of millions of years, not just thousands. Pioneers such as Prof. Mary Schweitzer led the original charge that made reporting on discoveries like ours much easier today.
“I know how the birds fly, how the fishes swim, how animals run. But there is the Dragon. I cannot tell how it mounts on the winds through the clouds and flies through heaven. Today I have seen the Dragon.“– Confucius, ca. 500 BCE.
Let’s finish with some images of a living bird’s ankle region, by co-author and PhD student Sophie Regnault. We considered these for inclusion in the paper but they didn’t fit quite right. I love them anyway so here they are:
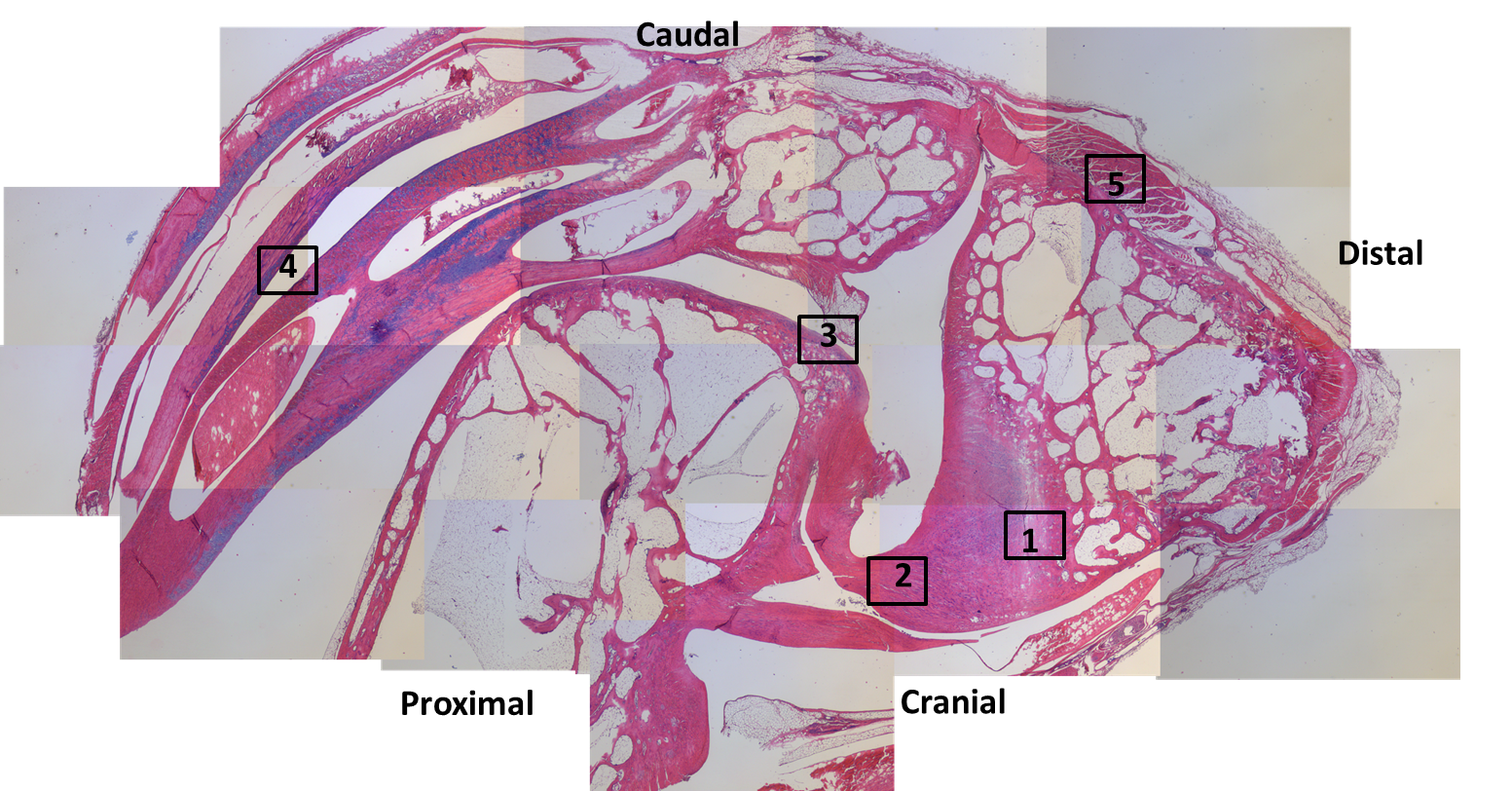
Patchwork of histology slide images, from a guineafowl’s ankle (as per photo above). The numbered squares correspond to zoomed-in images below. The tibiotarsus is on the proximal end (bottom left); the tarsometatarsus is on the distal end (right side); and the enigmatic tarsal sesamoid is at the top. Magnification: 20x overall.
Region 1. nice (fibro)cartilage-bone inferface at ligament insertion.
Region 2: longitudinal slice through ligaments connecting the tibiotarsus to the tarsometatarsus across the ankle joint.
Region 3: front (bottom) of the tibiotarsus/upper ankle.
Region 4: tendon fibres in longitudinal section; on the back of the tibiotarsus. Some show mineralization into ossified tendons (“metaplasia”); another curious feature of modern birds.
Region 5: muscle attachment to the back of the upper tarsometatarsus bone. Small sesamoid fragment visible.
The early, hippo-like mammal Coryphodon. I didn’t know it had a patella but it does. From Yale Peabody Museum.
I’m not shy about my fondness for the patella (kneecap) of tetrapod vertebrates, and neither are the other members of RVC’s “Team Patella”. We’ve had a fun 3+ years studying these neglected bones, and today we’ve published a new study of them. Our attention has turned from our prior studies of bird and lepidosaur kneecaps to mammalian ones. Again, we’ve laid the groundwork for a lot of future work by focusing on (1) basic anatomy and (2) evolutionary history of these sesamoid bones, with a lot of synthesis of existing knowledge from the literature; including development and genetics. This particular paper is a sizeable monograph of the state of play in the perusal of patellae in placental and other synapsids. Here’s what we did and found, focusing mostly on bony (ossified) patellae because that allowed us to bring the fossil record better to bear on the problem.
Reference: Samuels, M., Regnault, S., Hutchinson, J.R. 2016. Evolution of the patellar sesamoid bone in mammals. PeerJ 5:e3103 https://doi.org/10.7717/peerj.3103
Stomach-Churning Rating: 1/10; bones and more bones.
The short version of the story is that mammals evolved bony kneecaps about five times, with marsupials gaining and losing them (maybe multiple times) whereas monotremes (platypus and echidna) and placentals (us and other mammals) didn’t do much once they gained them, and a couple of other fossil groups evolved patellae in apparent isolation.
Evolution of the patella in mammals: broad overview from our paper. Click to zoom in.
The marsupial case is the most fascinating one because they may have started with a fibrocartilaginous “patelloid” and then ossified that, then reduced it to a “patelloid” again and again or maybe even regained it. There needs to be a lot more study of this group to see if the standard tale that “just bandicoots and a few other oddballs have a bony patella” is true for the Metatheria (marsupials + extinct kin). And more study of the development of patellae in this group could help establish whether they truly do “regress” into fibrocartilage when they are “lost” in evolution, or if other, more flexible patterns exist, or even if some of the cases of apparent “loss” of a bony patella are actually instances of delayed ossification that only becomes evident in older adults. Our paper largely punts on these issues because of an absence of sufficient data, but we hope that it is inspiration for others to help carry the flag forward for this mystery.
The higgledy-piggledy evolution of a patella in Metatheria, including marsupials. Click to zoom in.
Some bats, too, do funky things with their kneecaps, analogous to the marsupial “patelloid” pattern, and that chiropteran pattern also is not well understood. Why do some bats such as Pteropus fruit bats “lose” their kneecaps whereas others don’t, and why do some bats and other species (e.g. various primates) seem to have an extra thing near their kneecaps often called a “suprapatella”? Kneecap geeks need to know.
The short-nosed bandicoot (marsupial) Isoodon, showing a nice bony patella as typifies this group. From Yale Peabody Museum.
Otherwise, once mammals evolved kneecaps they tended to keep them unless they lost their hindlimbs entirely (or nearly so). Witness the chunky patellae of early whales such as Pakicetus and join us in wondering why those chunks persisted. The evolutionary persistence of blocky bits of bone in the knees of various aquatic animals, especially foot-propelled diving birds, may help answer why, as the hindlimbs surely still played roles in swimming early in cetacean evolution. Ditto for sea cows (Sirenia) and other groups.
Early whale Ambulocetus, showing hefty kneecaps.
But I’m still left wondering why so many groups of land vertebrates (and aquatic ones, too) never turned parts of their knee extensor tendons into bone. We know a bit about the benefits of doing that, to add leverage to those joints that enables the knee muscles to act with dynamic gearing (becoming more forceful “low gear” or more speedy “high gear” in function). Non-avian (and most early avian/avialan) dinosaurs, crocodiles, turtles, amphibians, early mammal relatives, and almost all other known extinct lineages except for those noted above got by just fine without kneecaps, it seems, even in cases where a naïve biomechanist would expect them to be very handy, such as in giant dinosaurs.
A quoll, Dasyurus, with what is probably a fibrocartilaginous “patelloid”. From Yale Peabody Museum.
However, tendons don’t turn to bone unless the right stresses and strains are placed upon them, so maybe kneecaps are a “spandrel” or “exaptation” of sorts, to abuse Gould’s ghost, whose adaptive importance is overemphasized. Maybe that adaptive myopia overshadows a deeper ontogenetic story, of how tissues respond to their history of mechanical loading environment. It has been speculated that maybe (non-marsupial) mammals have broadly “genetically assimilated” their kneecaps, fixing them into semi-permanence in their genetic-developmental programmes, whereas in contrast the few studies of birds indicate more responsiveness and thus less assimilation/fixation. That “evo-devo-mechanics” story is what now fascinates me most and we’ve poked at this question a bit now, with some updates to come- watch this space! Regardless, whether an animal has a bony vs. more squishy soft tissue patella must have consequences for how the knee joint and muscles are loaded, so this kind of question is important.
Giant marsupial Diprotodon (at NHM London); to my knowledge, not known to have had kneecaps- why?
In the meantime, enjoy our latest contribution if it interests you. This paper came about when first author Dr. Mark Samuels emailed me in 2012, saying he’d read some of my old papers on the avian musculoskeletal system and was curious about the evolution of patellae in various lineages. Unlike many doctors and vets I’ve run into, he was deeply fascinated by the evolutionary and fossil components of patellae and how those relate to development, genetics and disorders of patellae. We got talking, found that we were kindred kneecap-spirits, and a collaboration serendipitously spun off from that, soon adding in Sophie. It was a blast!
Last year on Darwin Day I debuted “Better Know A Muscle” (BKAM), which was intended to be a series of posts focusing on one cool muscle at a time, and its anatomical, functional and evolutionary diversity and history. A year later, it’s another post on another muscle! Several dozen more muscles to go, so I’ve got my work cut out for me… But today: get ready to FLEX your myology knowledge! Our subject is Musculus biceps brachii; the “biceps” (“two-headed muscle of the arm”). Beloved of Arnie and anatomists alike, the biceps brachii is. Let’s get pumped up!
Stomach-Churning Rating: 7/10. Lots of meaty elbow flexion!
While the previous BKAM’s topic was a hindlimb muscle with a somewhat complex history (and some uncertainties), the biceps brachii is a forelimb muscle with a simpler, clearer history. Fish lack a biceps, just having simple fin ab/adductor muscles with little differentiation. Between fish and tetrapods (limb-bearing vertebrates), there was an explosion in the number of muscles; part of transforming fins into limbs; and the biceps is thenceforth evident in all known tetrapods in a readily identifiable anatomical form. In salamanders and their amphibian kin, there is a muscle usually called “humeroantebrachialis” that seems to be an undivided mass corresponding to the biceps brachii plus the brachialis (shorter humerus-to-elbow) muscle:
Most of the humerobrachialis muscle (purplish colour), in dorsal (top) view of the right forelimb of the fire salamander Salamandra salamandra (draft from unpublished work by my team).
In all other tetrapods; the amniote group (reptiles, mammals, etc.); there is a separate biceps and brachialis, so these muscles split up from the ancestrally single “humeroantebrachialis” muscle sometime after the amphibian lineage diverged from the amniotes. And not much changed after then– the biceps is a relatively conservative muscle, in an evolutionary (not political!) sense. In amniote tetrapods that have a biceps, it develops as part of the ventral mass of the embryonic forelimb along with other muscles such as the shorter, humerus-originating brachialis, from which it diverges late in development (reinforcing that these two muscles are more recent evolutionary divergences, too).
Biceps brachialis or humerobrachialis, the “biceps group” tends to originate just in front of the shoulder (from the scapula/coracoid/pectoral girdle), running in front of (parallel to) the humerus. It usually forms of two closely linked heads (hence the “two heads” name), most obviously in mammals; one head is longer and comes from higher/deeper on the pectoral girdle, whereas the other is closer to the shoulder joint and thus is shorter. The two heads fuse as they cross the shoulder joint and we can then refer to them collectively as “the biceps”. It can be harder to see the longer vs. shorter heads of the biceps in non-mammals such as crocodiles, or they may be more or less fused/undifferentiated, but that’s just details of relatively minor evolutionary variation.
The biceps muscle then crosses in front of the elbow to insert mainly onto the radius (bone that connects your elbow to your wrist/thumb region) and somewhat to the ulna (“funny bone”) via various extra tendons, fascia and/or aponeuroses. The origin from the shoulder region tends to have a strong mark or bony process that identifies it, such as the coracoid process in most mammals (I know this well as I had my coracoid process surgically moved!). The insertion onto the radius tends to have a marked muscle scar (the radial tuberosity or a similar name), shared with the brachialis to some degree. A nice thing about the biceps is that, because it may leave clear tendinous marks on the skeleton, we sometimes can reconstruct how its attachments and path evolved (and any obvious specializations; even perhaps changes of functions if/when they happened).
Here are some biceps examples from the world of crocodiles:
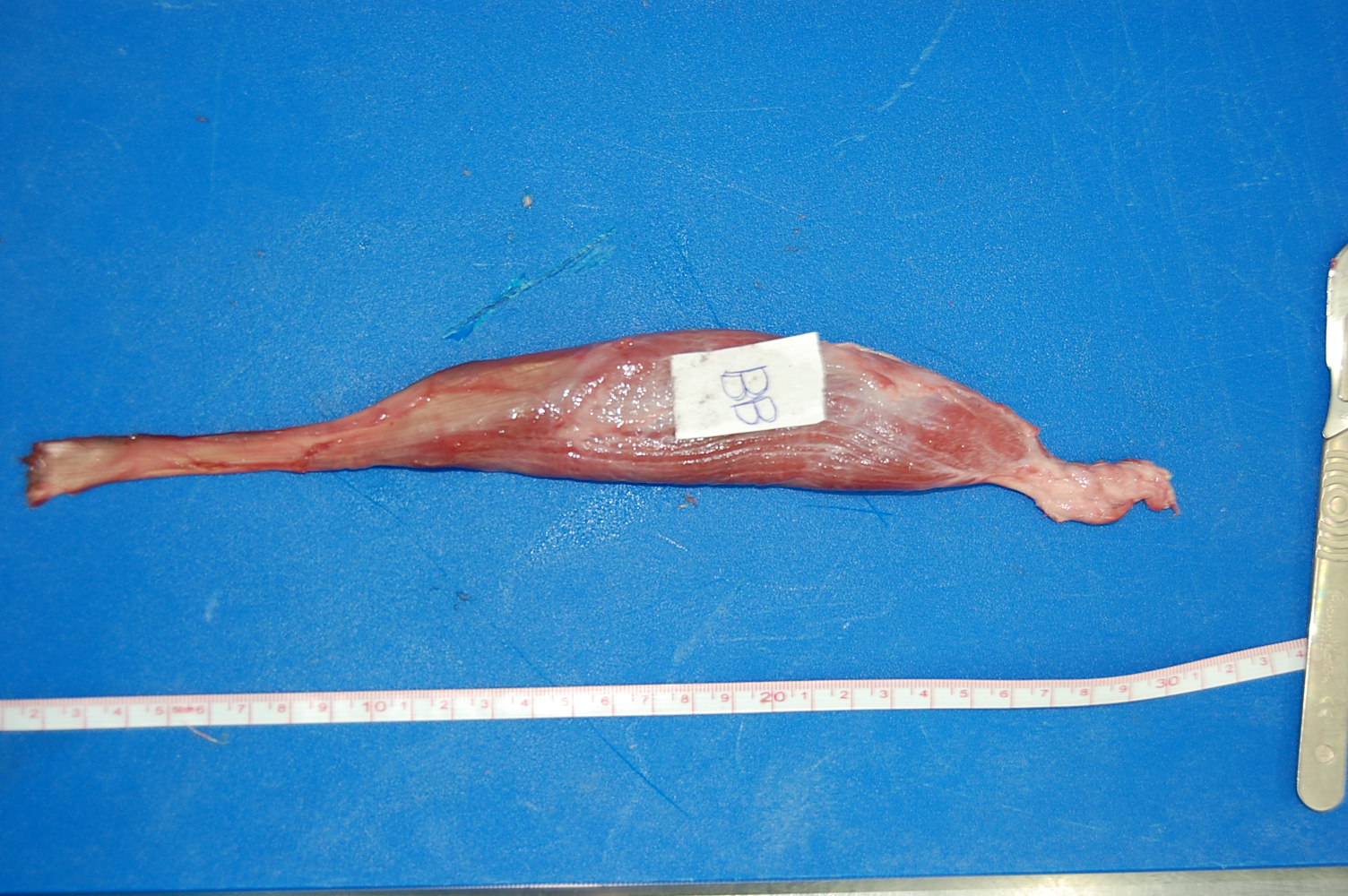
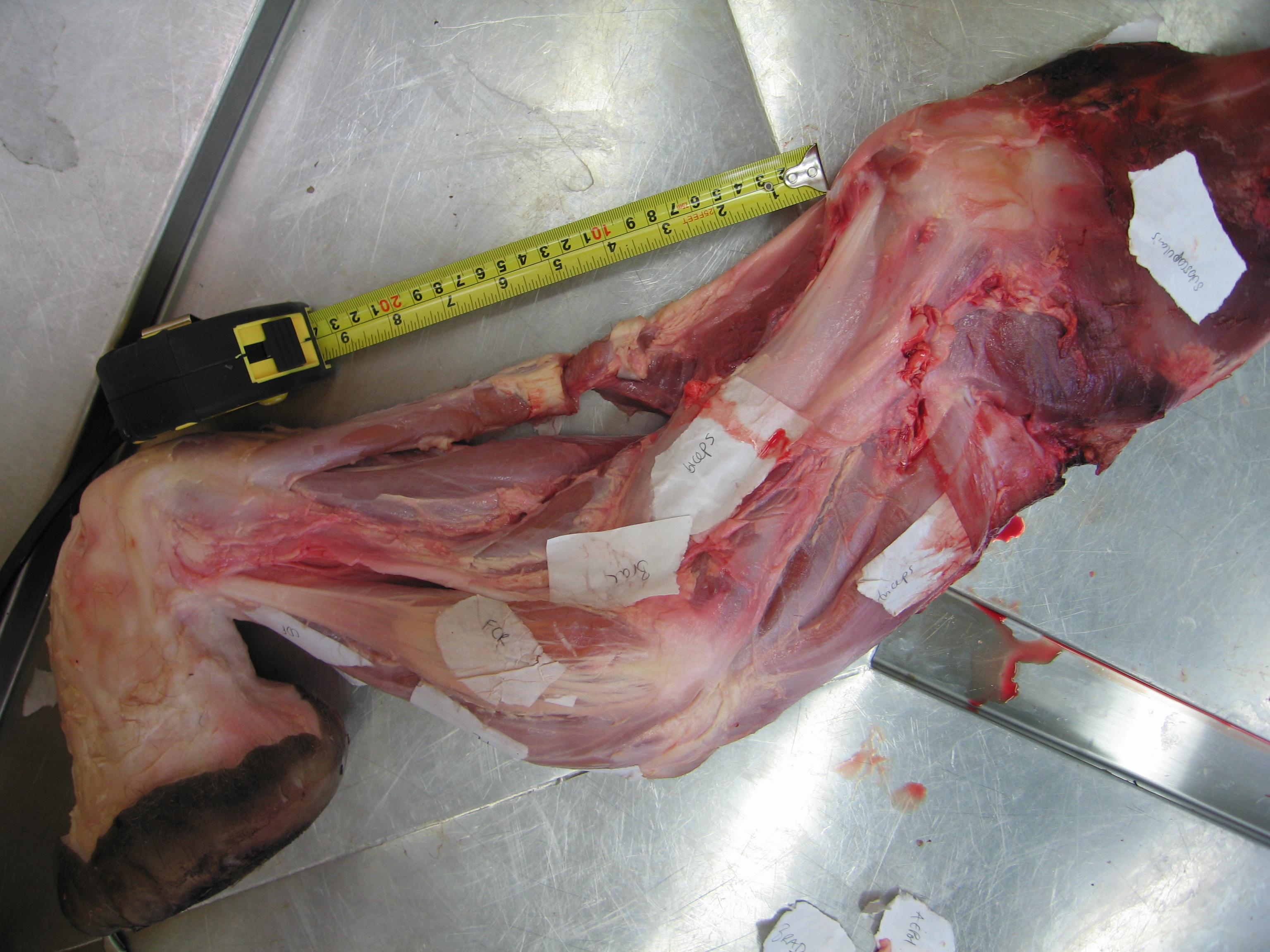
Crocodile’s right forelimb showing the huge pectoralis, and the biceps underlying it; on the bottom right (“BB”- click to embiceps it).
Crocodile left forelimb with biceps visible (“BB”) on the left.
Crocodile biceps muscle cut off, showing the proximal (to right) and distal (to left) tendons (and long parallel muscle fibres) for a typical amniote vertebrate.
What does the biceps muscle do? It flexes (draws forward) the shoulder joint/humerus, and does the same for the elbow/forearm while supinating it (i.e. rotating the radius around the ulna so that the palm faces upwards, in animals like us who can rotate those two bones around each other). In humans, which have had their biceps muscles studied by far the most extensively, we know for example that the biceps is most effective at flexing the elbow (e.g. lifting a dumbbell weight) when the elbow is moderately straight. These same general functions (shoulder and elbow flexion; with some supination) prevail across the biceps muscle of [almost; I am sure there are exceptions] all tetrapods, because the attachments and path of the biceps brachii are so conservative.
And this flexor function of the biceps brachii stands in contrast to our first BKAM muscle, the caudofemoralis (longus): that muscle acts mainly during weight support (stance phase) as an antigravity/extensor muscle, whereas the flexor action of biceps makes it more useful as a limb protractor or “swing phase” muscle used to collapse the limb and draw it forwards during weight support. However, mammals add some complexity to that non-supportive function of the biceps…
Hey mammals! Show us your biceps!
Jaguar forelimb with biceps peeking out from the other superficial muscles, and its cousin brachialis nicely visible, running along the front of the forearm for a bit.
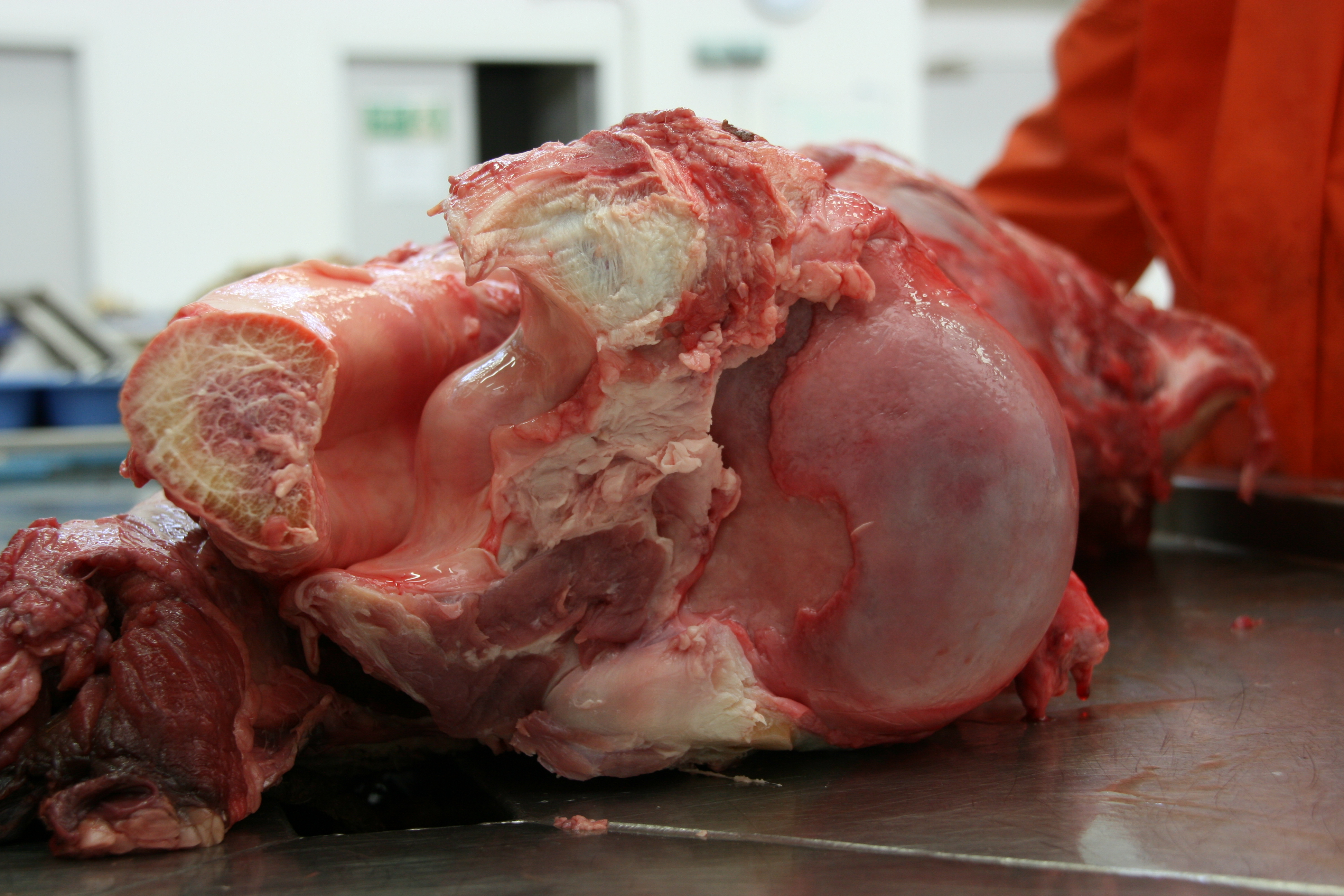
Elephant’s left forelimb with the biceps labelled.
Longitudinal slice thru the biceps of an elephant, showing the internal tendon that helps identify where the two bellies of the biceps fuse.
In certain mammals; the phylogenetic distribution of which is still not clear; the biceps brachii forms a key part of a passive “stay apparatus” that helps keep the forelimb upright against gravity while standing (even sleeping). The classic example is in horses but plenty of other quadrupedal mammals, especially ungulate herbivores, show evidence of similar traits:
Giraffe biceps cut away proximally to show the “stay apparatus” around the shoulder joint (upper right).
Zooming in on the “stay apparatus”; now in proximal view, with the biceps tendon on the left and the humeral head (showing some arthritic damage) on the right, with the groove for the biceps in between.
Hippo’s humerus (upper left) and biceps muscle cut away proximally, displaying the same sort of “stay apparatus” as in the giraffe. Again, note the stout proximal and distal tendons of the biceps. The proximal tendon fits into the groove of the humerus on the far left side of the image; becoming constrained into a narrow circular “tunnel” there. It’s neat to dissect that region because of its fascinating relationships between bone and soft tissues.
The biceps brachii, in those mammals with a stay apparatus, seems to me to have a larger tendon overall, especially around the shoulder, and that helps brace the shoulder joint from extending (retracting) too far backward, whilst also transmitting passive tension down the arm to the forearm, and bracing the elbow (as well as distal joints via other muscles and ligaments). It’s a neat adaptation whose evolution still needs to be further inspected.
Otherwise, I shouldn’t say this but the biceps is sort of boring, anatomically. Whether you’re a lizard, croc, bird or mammal, a biceps is a biceps is a biceps; more or less-ceps. But the biceps still has a clear evolutionary history and Darwin would gladly flex his biceps to raise a pint in toast to it.
So now we know a muscle better. That’s two muscles now. And that is good; be you predator or prey. Let’s shake on it!