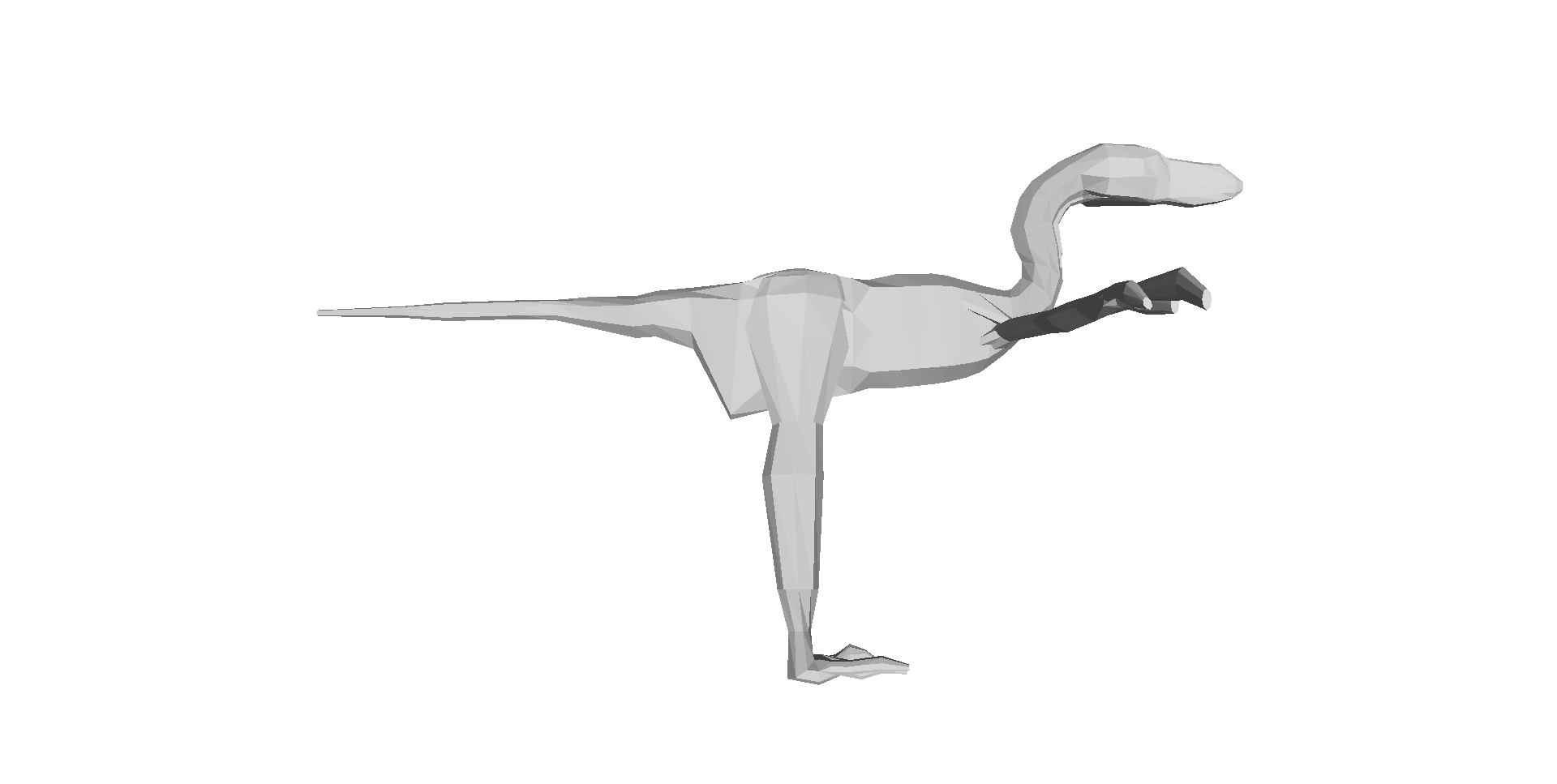If you’re in London, you still have almost one week left to hurry to the Valence House in Dagenham and see a great exhibit on Ray Harryhausen’s dinosaurs and other cool “Dynamation” stop-motion models and art!
This blog post is a photo tour of what I saw, in case you cannot go.
I loooooooooooove Ray Harryhausen’s work, ever since I was a child and saw “Jason and the Argonauts” and many other films, plus “Clash of the Titans” once it came into theatres. There is the attention to detail in anatomy and locomotion, and the wondrous fantastic nature of even the more mundane creatures he animated, and the rich mythology that he drew from to inspire his creations. Modern CGI is great in a different way, but nothing I can think of in recent special effects truly beats (1) the skeleton battle in ‘Jason, and (2) the Medusa encounter in ‘Clash (to name what might be my top two faves). And so when I learned that several of the original (restored) models from those films were on exhibit in northeastern London, I requested to go there with my family for Fathers Day. Results:
Boom! Ole’ stony-gazed, snaky-haired gorgon of yore.
No deadly bow here, but the rattlesnake tail is.
Medusa concept art by Harryhausen; the “bra” was there for American censors but Ray thought it looked wrong and removed it in the final version.
Look out, Jason! Here come the Children of the Hydra! Yep, original (restored) articulated models. Joints are visible. They look ready to kick some Iolcusian butt!
Context of the exhibit- local chap befriended Harryhausen and convinced him to let him restore his models; and so here we are. On with the dinosaurs! (and other palaeo-things)
Gwangi model made in resin; non-poseable but made around time of the “Valley of Gwangi” film to help design the poseable models.
Gwangi climactic scene in church; concept art by Harryhausen.
Other ‘Gwangi characters: “Eohippus” (Hyracotherium), Ornithomimus and boy.
Cowboy lassoing an Ornithomimus as per the movie scene in ‘Gwangi? Yes please. (Harryhausen original) Jurassic Park had its T. rex lurching out of a forest to grab a Struthiomimus, intentionally mirroring the scene in ‘Gwangi where the titular Allosaurus–Tyrannosaurus hybrid chomps the Ornithomimus.
Poseable “Eohippus” original- with real fur! Great Dynamation too; very lifelike in the film.
Original Harryhausen concept art of the “Eohippus” show demo.
Suddenly, Ceratosaurus! (from “One Million Years BC”)
Styracosaurus original resin model. (from “One Million Years BC”)
Old school Polacanthus art by Alan Friswell. SPIKEY!
Old school Iguanodon art by Alan Friswell. MUSCLEY!
Panoply of archosaurs by Alan Friswell: pterodactyl, Tenontosaurus (made for the Frame Store special effects company in 2001) and tyrannosaur head (made at age 9).
Pterodactyl made at age 12, so don’t laugh.
Back to the fantastic beasts– original poseable hydra from ‘Jason!
Original Pegasus from ‘Clash! What a seamless blend of fur and feathers.
Original R2, I mean Bubo, from ‘Clash!
I forget the scene (the 1-eyed fates in ‘Clash?) but I like it. Original Harryhausen concept art.
Lunar leader from “First Men in the Moon.” (original)
Non-original (but based thereon) model by Alan Friswell, of nautiloid thingy from “Mysterious Island”.
Fiji mermaid by Alan Friswell.
“Hand of Glory” by Alan Friswell.
Pithecanthropus by Alan Friswell. Very Harryhausen in spirit.
Oddly, but somehow appropriately, there are ?350 year old whale bones on display in the hall next door, with a mysterious history.
WW2 bomb shelter in a “Victory Garden” outside the House. And the house is supposedly haunted. So take care when you visit…
What can I say? I loved it! Almost a religious experience; like seeing holy relics. Awesome in every sense of awesome.
Downside: you cannot grab the precious Dynamation models and play with them hands-on. I wanted to enact a furious Hydra-Gwangi battle. But alas, only in my imagination…
As 2017 approaches its end, there have been a few papers I’ve been involved in that I thought I’d point out here while I have time. Our DAWNDINOS project has been taking up much of that time and you’ll see much more of that project’s work in 2018, but we just published our first paper from it! And since the other two recent papers involve a similar theme of muscles, appendages and computer models of biomechanics, they’ll feature here too.
Stomach-Churning Rating: 0/10; computer models and other abstractions.
Mussaurus patagonicus was an early sauropodomorph dinosaur from Argentina, and is now widely accepted to be a very close relative of the true (giant, quadrupedal) sauropods. Here is John Conway’s great reconstruction of it:
We have been working with Alejandro Otero and Diego Pol on Mussaurus for many years now, starting with Royal Society International Exchange funds and now supported by my ERC grant “DAWNDINOS”. It features in our grant because it is a decent example of a large sauropodomorph that was probably still bipedal and lived near the Triassic-Jurassic transition (~215mya).
In our new study, we applied one of my team’s typical methods, 3D musculoskeletal modelling, to an adult Mussaurus’s forelimbs. This is a change of topic from the hindlimbs that I’ve myopically focused on before with Tyrannosaurus and Velociraptor [in an obscure paper that I should never have published in a book! pdf link], among other critters my team has tackled (mouse, elephant [still to be finished…], ostrich, horse, Ichthyostega… dozens more to come!). But we also modelled the forelimbs of Crocodylus johnstoni (Australian “freshie”) for a key comparison with a living animal whose anatomy we actually knew, rather than reconstructed.
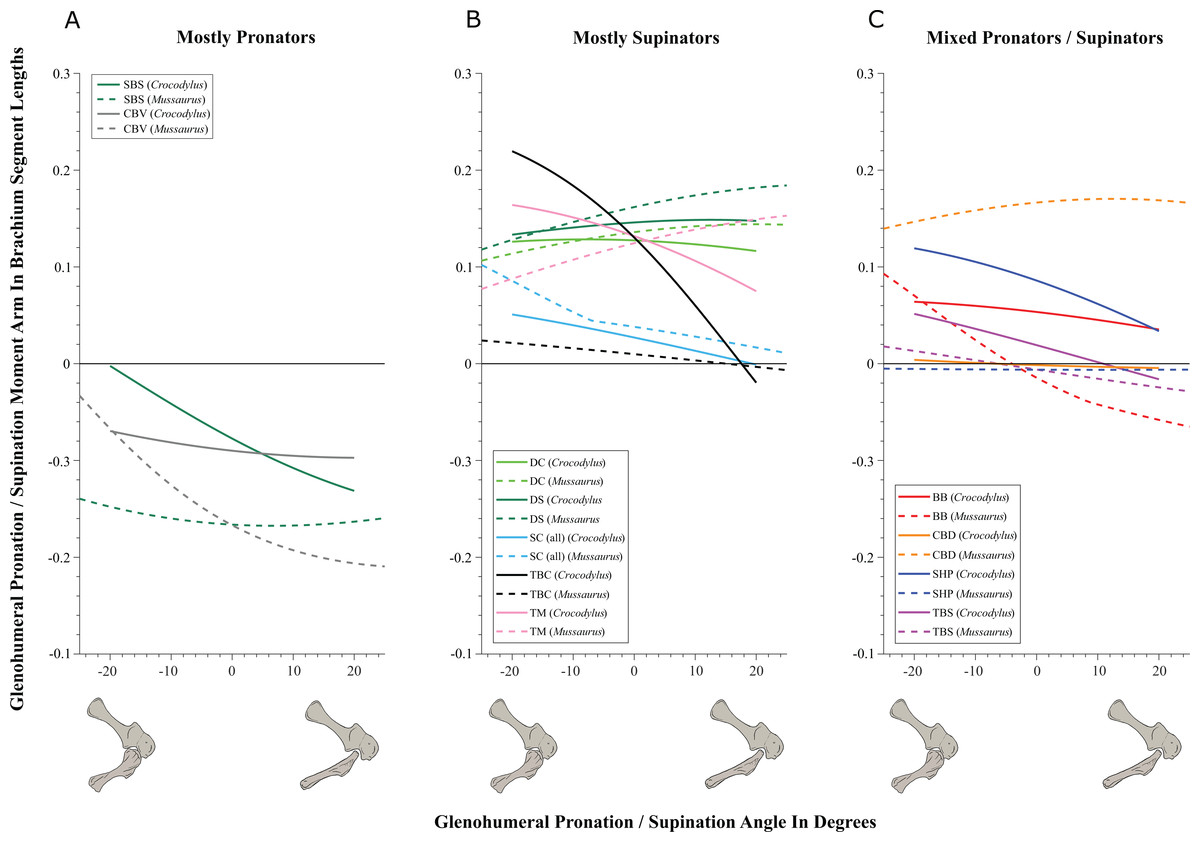
Mussaurus above; Crocodylus below; forelimb models in various views; muscles are red lines.
The methods for this biomechanical modelling are now standard (I learned them from their creator Prof. Scott Delp during my 2001-2003 postdoc at Stanford): scan bones, connect them with joints, add muscle paths around them, and then use the models to estimate joint ranges of motion and muscle moment arms (leverage) around joints. I have some mixed feelings about developing this approach in our 2005 paper that is now widely used by the few teams that study appendicular function in extinct animals. As a recent review paper noted and I’ve always cautioned, it has a lot of assumptions and problems and one must exercise extreme caution in its design and interpretation. Our new Mussaurus paper continues those ruminations, but I think we made some progress, too.
On to the nuts and bolts of the science (it’s a 60 page paper so this summary will omit a lot!): first, we wanted to know how the forelimb joint ranges of motion in Mussaurus compared with those in Crocodylus and whether our model of Mussaurus might be able to be placed in a quadrupedal pose, with the palms at least somewhat flat (“pronated”) on the ground. Even considering missing joint cartilage, this didn’t seem very plausible in Mussaurus unless one allowed the whole forearm to rotate around its long axis from the elbow joint, which is very speculative—but not impossible in Crocodylus, either. Furthermore, the model didn’t seem to have forelimbs fully adapted yet for a more graviportal, columnar posture. Here’s what the model’s mobility was like:
So Mussaurus, like other early sauropodomorphs such as Plateosaurus, probably wasn’t quadrupedal, and thus quadrupedalism must have evolved very close to in the Sauropoda common ancestor.
Second, we compared the muscle moment arms (individual 3D “muscleactions” for short) in different poses for all of the main forelimb muscles that extend (in various ways and extents) from the pectoral girdle to the thumb, for both animals, to see how muscle actions might differ in Crocodylus (which would be closer to the ancestral state) and Mussaurus. Did muscles transform their actions in relation to bipedalism (or reversal to quadrupedalism) in the latter? Well, it’s complicated but there are a lot of similarities and differences in how the muscles might have functioned; probably reflecting evolutionary ancestry and specialization. What I found most surprising about our results was that the forelimbs didn’t have muscles well-positioned to pronate the forearm/hand, and thus musculoskeletal modelling of those muscles reinforced the conclusions from the joints that quadrupedal locomotion was unlikely. I think that result is fairly robust to the uncertainties, but we’ll see in future work.
You like moment arms? We got moment arms! 15 figures of them, like this! And tables and explanatory text and comparisons with human data and, well, lots!
If you’re really a myology geek, you might find our other conclusions about individual muscle actions to be interesting—e.g. the scapulohumeralis seems to have been a shoulder pronator in Crocodylus vs. supinator in Mussaurus, owing to differences in humeral shape (specialization present in Mussaurus; which maybe originated in early dinosaurs?). Contrastingly, the deltoid muscles acted in the same basic way in both species; presumed to reflect evolutionary conservation. And muuuuuuch more!
Do you want to know more? You can play with our models (it takes some work in OpenSim free software but it’s do-able) by downloading them (Crocodylus; Mussaurus; also available: Tyrannosaurus, Velociraptor!). And there will be MUCH more about Mussaurus coming soon. What is awesome about this dinosaur is that we have essentially complete skeletons from tiny hatchlings (the “mouse lizard” etymology) to ~1 year old juveniles to >1000kg adults. So we can do more than arm-wave about forelimbs!
But that’s not all. Last week we published our third paper on mouse hindlimb biomechanics, using musculoskeletal modelling as well. This one was a collaboration that arose from past PhD student James Charles’s thesis: his model has been in much demand from mouse researchers, and in this case we were invited by University of Virginia biomechanical engineers to join them in using this model to test how muscle fibres (the truly muscle-y, contractile parts of “muscle-tendon units”) change length in walking mice vs. humans. It was a pleasure to re-unite in coauthorship with Prof. Silvia Blemker, who was a coauthor on that 2005 T. rex hindlimb modelling paper which set me on my current dark path.
Mouse and human legs in right side view, going through walking cycles in simulations. Too small? Click to embiggen.
We found that, because mice move their hindlimb joints through smaller arcs than humans do during walking and because human muscles have large moment arms, the hindlimb muscles of humans change length more—mouse muscles change length only about 48% of the amount that typical leg muscles do in humans! This is cool not only from an evolutionary (mouse muscles are probably closer to the ancestral mammalian state) and scaling (smaller animals may use less muscle excursions, to a point, in comparable gaits?) perspective, but it also has clinical relevance.
Simulated stride for mouse and human; with muscles either almost inactive (Act=0.05) or fully active (Act=1). Red curve goes through much bigger excursions (along y-axis) than blue curve), so humans should use bigger % of their muscle fibre lengths in walking. Too small? Click to embiggen.
My coauthors study muscular dystrophy and similar diseases that can involve muscle stiffness and similar biomechanical or neural control problems. Mice are often used as “models” (both in the sense of analogues/study systems for animal trials in developing treatments, and in the sense of computational abstractions) for human diseases. But because mouse muscles don’t work the same as human muscles, especially in regards to length changes in walking, there are concerns that overreliance on mice as human models might cause erroneous conclusions about what treatments work best to reduce muscle stiffness (or response to muscle stretching that causes progressive damage), for example. Thus either mouse model studies need some rethinking sometimes, or other models such as canines might be more effective. Regardless, it was exciting to be involved in a study that seems to deliver the goods on translating basic science to clinical relevance.
Muscle-by-muscle data; most mouse muscles go through smaller excursions; a few go through greater; some are the same as humans’.
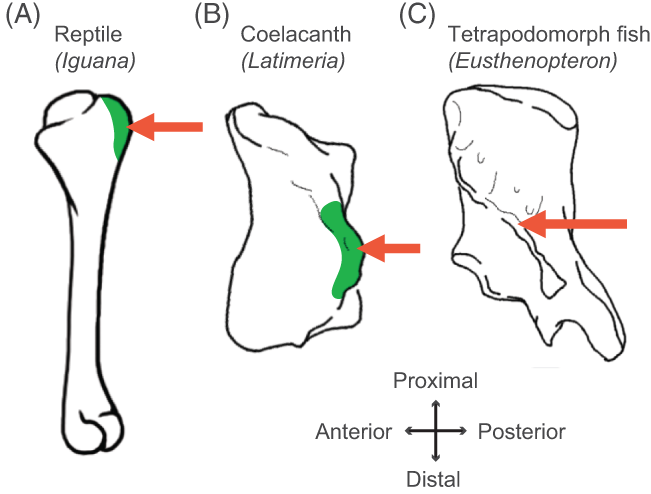
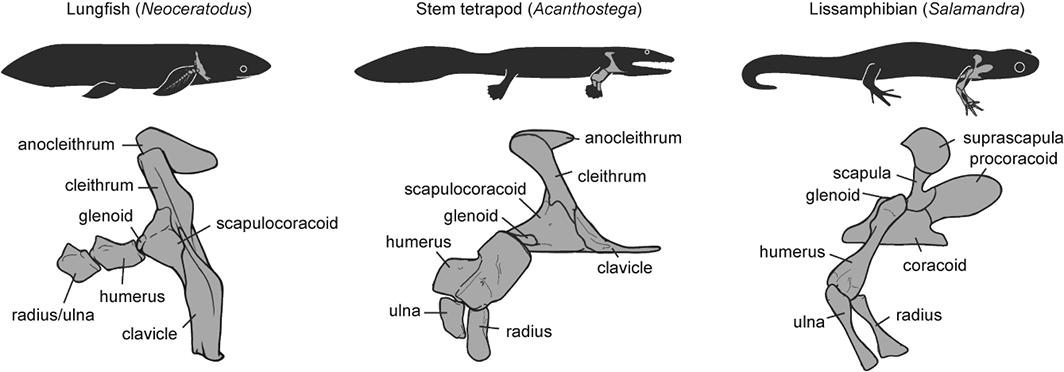
Finally, a third recent paper of ours was led by Julia Molnar and Stephanie Pierce (of prior RVC “Team Tetrapod” affiliation), with myself and Rui Diogo. This study tied together a bunch of disparate research strands of our different teams, including musculature and its homologies, the early tetrapod fossil record, muscle reconstruction in fossils, and biomechanics. And again the focus was on forelimbs, or front-appendages anyway; but turning back the clock to the very early history of fishes, especially lobe-finned forms, and trying to piece together how the few pectoral fin muscles of those fish evolved into the many forelimb muscles of true tetrapods from >400mya to much more recent times.
Humerus in ventral view, showing muscle attachments. Extent (green) is unknown in the fossil but the muscle position is clear (arrow).
We considered the homologies for those muscles in extant forms, hypothesized by Diogo, Molnar et al., in light of the fossil record that reveals where those muscles attach(ed), using that reciprocal illumination to reconstruct how forelimb musculature evolved. This parallels almost-as-ancient (well, year 2000) work that I’d done in my PhD on reconstructing hindlimb muscle evolution in early reptiles/archosaurs/dinosaurs/birds. Along the way, we could reconstruct estimates of pectoral muscles in various representative extinct tetrapod(omorph)s.
Disparity of skeletal pectoral appendages to work with from lobe-fins to tetrapods.
Again, it’s a lengthy, detailed study (31 pages) but designed as a review and meta-analysis that introduces readers to the data and ideas and then builds on them in new ways. I feel that this was a synthesis that was badly needed to tie together disparate observations and speculations on what the many, many obvious bumps, squiggles, crests and tuberosities on fossil tetrapods/cousins “mean” in terms of soft tissues. The figures here tell the basic story; Julia, as usual, rocked it with some lovely scientific illustration! Short message: the large number of pectoral limb muscles in living tetrapods probably didn’t evolve until limbs with digits evolved, but that number might go back to the common ancestor of all tetrapods, rather than more recently. BUT there are strong hints that earlier tetrapodomorph “fishapods” had some of those novel muscles already, so it was a more stepwise/gradual pattern of evolution than a simple punctuated event or two.
Colour maps of reconstructed right fin/limb muscles in tetrapodomorph sarcopterygian (~”fishapod”) and tetrapod most recent common ancestors. Some are less ambiguous than others.
That study opens the way to do proper biomechanical studies (like the Mussaurus study) of muscle actions, functions… even locomotor dynamics (like the mouse study)– and ooh, I’ve now tied all three studies together, tidily wrapped up with a scientific bow! There you have it. I’m looking forward to sharing more new science in 2018. We have some big, big plans!
Tonight is the debut of the ballyhooed BBC1 programme “Attenborough and the Giant Dinosaur“, featuring Sir David and The Titanosaur-With-No-Name, at 6:30pm. Furthermore, this week I presided over a showing of “T. rex Autopsy” to our RVC undergrad Zoological Society, with a very enjoyable Q&A afterwards. So it seemed timely for me to do a post about a theme that links these two documentaries that I helped with, my own research, and science communication and palaeontological research more generally. But first let’s get this out of the way:
It was great. I could gush more but that’s not what this post is about.
Stomach-Churning Rating: ~7/10; mainly the elephant leg dissection that’s not far below, which is a bit messy.
For the titanosaur documentary with Sir David, and the incomparable Ben Garrod as well, we had an old elephant “friend” of mine (subject of many biomechanics studies we’d done) walk across a pressure pad to demonstrate how the elephant locomotion works and some of the basic similarities with how a giant sauropod dinosaur might walk:
A broader feature of that documentary, which elephants are linked into, is how we can use the skeleton to reconstruct some general aspects of the soft tissue anatomy, and thereby the physiology or even behaviour, of a giant titanosaur. Which brings me to this post’s subject: We dig up dinosaurs all the time, but what about digging into dinosaurs and using what’s preserved to reconstruct what isn’t?
Some of the “G-suit” compressive stocking that the fascia wrapped around elephant, and other large mammals, creates, and the underlying, interwoven muscles and tendons (hindlimb of a young Asian elephant that sadly died in captivity). Did some larger dinosaurs have something like this? I’d expect so.
Another view, more superficially, of that G-suit/stocking under the thick, tight skin of an elephant’s leg. You’ll hear more about this in the Attenborough show…
Once the documentary airs, I may be able to share some more images from it showing what they did for the titanosaur, but this BBC photo gives a good idea. Here, blood vessels and other tissues surrounding the skeleton. How would a titanosaur pump blood around its body? A good question.
I wanted to focus more on the end product; the imagery or even physical object; and how it conveys what we think we know about dinosaur anatomy. I’ll do that via examples, using photos of dinosaur anatomy that I’ve collected over the years from museums or other media. There won’t be any profound points or long musings; it’s mainly a photo blog:
What your (inner?) child most needs is a dinosaur to dissect yourself! Why not a T. rex toy like this?
For the price (~$30 in USA), the 4D Vision dinosaurs deliver a pretty good bargain, and the anatomy is satisfactory. I’ve been collecting this series. I could quibble, but hey: it’s a dinosaur you get to build/dissect yourself, and with many major organs in reasonable positions! Not so easy to put/keep together, but it’s fine. I would not pay a ton for it, though.
Poster of Velociraptor’s anatomy I’ve had since grad school, adorning my office. For ~1996 (no feathers; “zombie hands“), it’s damn good, mostly… Closer views below (sorry, photo quality is crap– taking photos of wall poster turned out to be harder than I expected! Bad lighting.) :
Closeup of the leg muscles- hey, not bad! Pretty much the right muscles in the right places more or less, and plausible proportions. No air sacs in the torso, but again, this is mid-1990’s science shown. BUT…
I was happy with this poster until I got it home from the western-USA museum I bought it at and read this final bit of text… Oh, America! You silly place. (And unfortunately, these dinosaurs are not from the very end of the Cretaceous anyway, so “global catastrophe” is not implicated.)
Looking down onto the opened torso of the Barcelona ornithomimid. Strikingly realistic! Breastbone, heart, liver, intestines; not unreasonable positions and sizes. No feathers, but again hey– this was made in the earlier days.
Albertosaurus from the Drexel Academy of Sciences. I forget where I got this pic but I like the display– this is an impressive full-scale physical model. The transition from skeleton-only on the left to skinned in the middle to fully-fleshed and popping out atcha on the right is clever.
?T. rex? leg, photo that I took ages ago as a PhD student, if memory serves. Can anyone remind me where this was? California Academy of Sciences? I am embarrassed that I cannot recall. I remember geeking out about it. It has a toy-ish look, but I reckon they had to build this to withstand kids touching it.
Perhaps the best dino-dissection example I’ve seen in a museum: the AMNH’s sauropod Mamenchisaurus with internal organs and their functions projected onto it, in the “World’s Largest Dinosaurs” exhibit. Bravo! I stood and watched it for quite a while.
This is far from comprehensive– just several kinds of imagery that I mostly like. There’s the tension between showing too much realism, which science simply can’t back up, and being too cartoonish, losing the viewer’s immersion in the time-travelling fantasy. I do, however, like other kinds of more abstract, schematic depictions of dinosaur anatomy that simplify the details to focus on the basics of what organs should have been where and how they may have worked, such as this depiction from T. rex Autopsy, which also took the other extreme favouring ultra-realism (but with physical models, not so much with the CGI):
Air flow through a T. rex: simplified but clear. CGI used to explain, not abused. The real air sac anatomy would be too complex to show. You may see something similar with the titanosaur show.
That’s enough for now. I’ve stuck with relatively recent examples; of course in my particular field I also think back to Romer’s wonderful 1920’s drawings, which I covered in this post.
So, blog readers, help me out here: what examples of dinosaur internal, squishy anatomy from museums, documentaries or other not-entirely-done-by-nitpicky-scientists venues do you like, or not like so much? What works for you, or at least is memorable in some way?
Well, that was an eventful week for me, although today’s post will focus on one event: the debut of the film Jurassic World. Briefly though, the awesome “T. rex Autopsy” documentary debuted (I was going to post more about it but all I’d have left to say is that I was very pleased with the result), I also showed up briefly in “Top 10 Biggest Beasts Ever” talking about the giant rhinocerotoid Paraceratherium and the stresses on its feet, our paper on ostrich musculoskeletal modelling was published (more in a future post) after ~12 years of me diddling around with it, and much more happened. Then to cap it all off, very shortly after I hit the “publish” button on my last post, I had four tonic clonic seizures in a row and spent a hazy night in the hospital, then the past week recovering from the damage. Nothing like another near-death (no exaggeration there, I’m afraid) experience to cap off an exciting week. But strangely, what I feel more interested in talking about is, like I said, Jurassic World, but this is not a review, as you will see here.
Stomach-Churning Rating: 0/10; just SPOILERS if you haven’t seen the film yet!
I guess I have to give a brief review of the film and say that I was entertained, to a degree, but it was not a great piece of film-making. It was a far cry from the original but then so were the sequels, and maybe it was better than them. The mosa-star was the most novel, memorable bit. I didn’t care for the Indominus villain, but then when you bring genetic engineering into a film like this, you’ve basically thrown out the rulebook and can make your dinosaurs as magical as you want; we’re already in “X-Men” territory here and almost in “Pacific Rim”-land.
Chris Pratt has signed on for at least another sequel to Jurassic World and the ending of the film already started that ball rolling. So I find it fun to speculate wildly, and certainly incorrectly, on what the sequel might do. What does the Jurassic future have in store?
First of all, who survived to re-appear in the next film? We’re left with the Bryce Dallas Howard character, who probably will return with Pratt to further develop their rather uninteresting social/romantic dynamic, rather than start afresh with someone else. The kids of course survived, as always, and as always they won’t return, as that’s not interesting and they didn’t have much to do except scream and (highly implausibly) hotwire an old car. Much as I’d like the parents from my hometown of Madison, WI to return, they won’t either for the same reasons. But we really only need Pratt and his high-heeled sweetie for the next film. Everyone else memorable(?) seems to have died, although it would be wonderful to bring Goldblum back for some smarmy wit (please!).
Second of all, the next film can’t be set in Jurassic World. There’s not much left to do there (JW already spent much of its time hearkening back to JP), and there’s no way the park would re-open. We need something new. I think by now we’re (very) tired of characters running around islands full of dinosaurs and the Blackfish parallel was milked dry in the latest movie. We need to spend a film with the dinosaurs amongst humanity (as Lost World briefly did), and much as I’d love to see the crazy drug lord/kidnapping plot happen, it won’t. But JW did set one thing up that has to happen now in its sequel: the paramilitary role of engineered, trained dinosaurs. We now know they can sort of train their dinosaurs and they can forge them to be anything they want to in terms of geno/phenotypes. They’ll learn from some mistakes of JW and engineer (or already did by the end of JW, at some remote site) some more compliant, deadlier animals, having largely given up on the public exhibition angle. The naked raptors and T. rex probably have to re-appear (sigh), but enough already of the giant uber-theropods like Spinosaurus and Indominus. The latter was already enough of a reprise of the former (plus psychic talents and chameleon powers etc.). Something truly novel is needed.
Unless they engineer a hyper-aggressive, intelligent sauropod or ceratopsian, which would admittedly be neat, I have this prediction (which is probably wrong but hey!): they have shown they can hybridize anything. There must be fewer and fewer “normal” (1990s…) dinosaurs now in the JW universe. So the next big step, which someone in the JW universe surely would do, is to hybridize dinosaurs and humans. Maybe some raptor-human hybrids, maybe also saving a tyranno-human hybrid for a surprise late appearance. But this is the sensible next step because it allows them to play with the (tired) Frankenstein monster trope but also touch on the hot topic of human cloning and human GM.
And by unleashing dino-human hybrids, or at least some freaky clicker-trained and engineered super-dinos, they could also explore the military theme, which the JW universe still hasn’t delved into much. What if those hyper-smart, deadly hybrid dinos, led by Pratt and Howard’s expert training, were used to combat an ISIS-analogue terrorist threat? Dino-Avengers in the badlands of Afghanistan or Iraq? Too predictable perhaps, but that’s a film that the public will want to see. Yeah there’s plenty of stupid there, but there’s no turning back– each film ups the ante, as JW ironically reminds us several times. We’re already in firmly in stupid-land, and the science has largely advanced to the point of magic. My idea is too uncomfortably close to the abandoned John Sayles plot, true. Darn. And (groan) kids have to be involved in some way to make it a family film so it rakes in the $$$ again, so either they get caught in the middle of the paramilitary mess or they are the ones that have to be saved… or the hybrid dinos are cute-ish kids themselves that Pratt and Howard must manage… (shades of Teenage Mutant Ninja Turtles/Kick-Ass?) I’ve found that more fun to think about than dwelling on the flaws of the movie, which is frankly too easy.
(Another good theme that Vivian Allen suggested to me would be climate change and invasive species—i.e. planet warms, dinos are already loose and go feral in waterlogged Central America, ecological disaster is looming and something must be done to round up the dinos… could work in some other bits like ecotourists or drug runners?)
That’s as far as my wildly speculative ruminating has taken me, but I wanted to turn it over to you, Freezerinos. If you were to make the next film (will it be “Jurassic World 2”? “Jurassic Army”? or as I’ve proffered in the post’s title, “Jurassic Future”?), what would it be (A) in your ideal world where you call all the shots (yes, lots of colourful feathery dinos, I know), vs. (B) in a more likely (less daring, more Hollywood) reality, along the lines of what I’ve tried to do here? (but I surely will be wrong, although we’ll see in 2-5 years!)
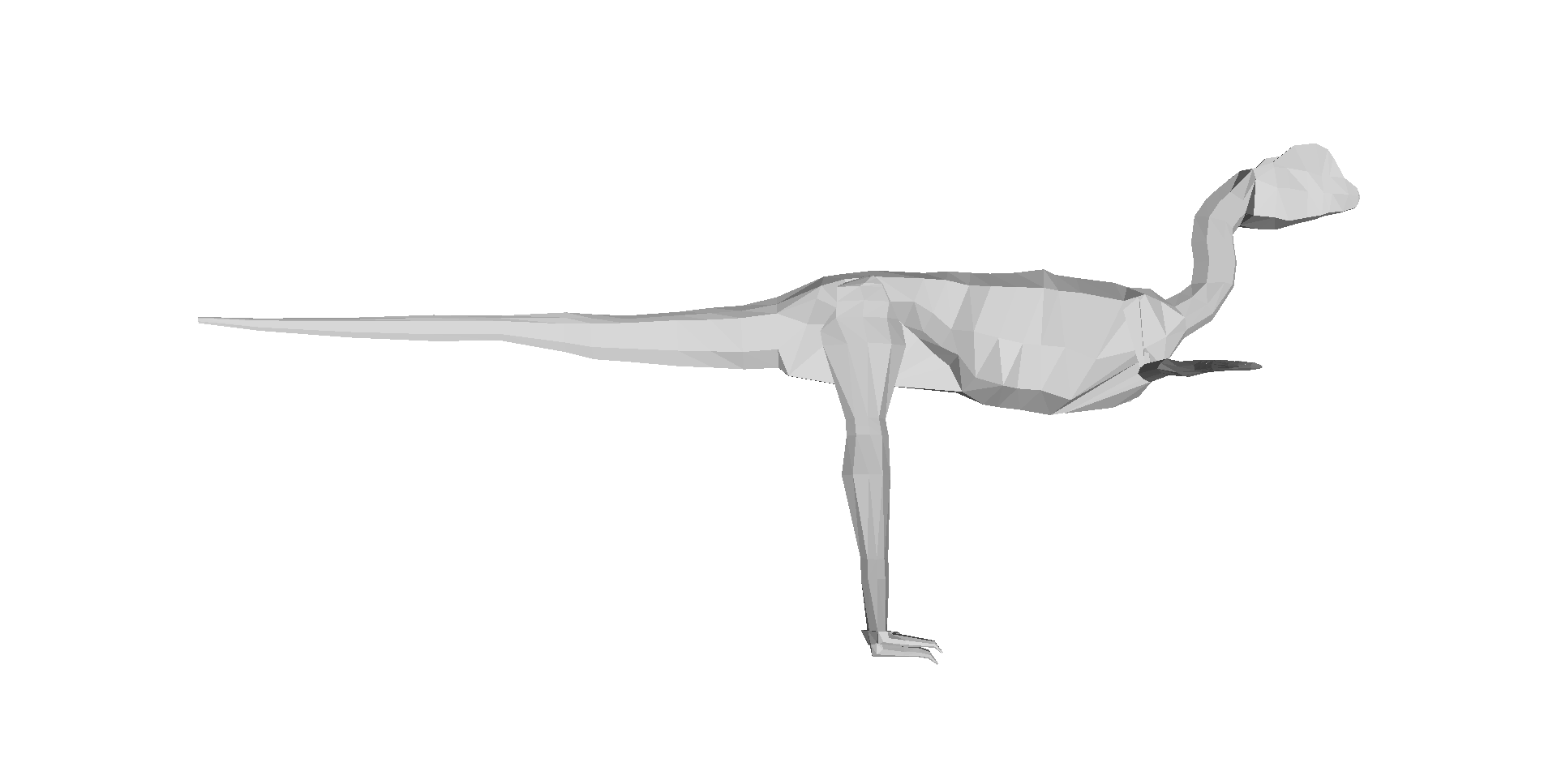
Our 3D computer models of a basal dinosaur and bird, showing methods and key differences in body shape. The numbers at the bottom are museum specimen numbers.
At about the moment I’m posting this, our Nature paper (our more formal page here, and the actual article here) embargo is ending, drawing a 14+ year gestation to a close. The paper is about how dinosaur 3D body shape changed during their evolution, and how that relates to changes in hindlimb posture from early dinosaurs/archosaurs to birds; “morpho-functional evolution” sums up the topic. We used the 3D whole-body computational modelling that I, Allen and Bates (among others) have developed to estimate evolutionary changes in body dimensions, rather than focusing on single specimens or (as in our last study) tyrannosaur ontogeny. We’ve strongly supported the notion (dating back to Gatesy’s seminal 1990 Paleobiology paper) that the centre of mass of dinosaurs shifted forwards during their evolution, and that this shift gradually led to the more crouched (flexed) hind leg posture that characterizes living birds. Here is a movie from our paper showing how we did the modelling:
And here is a summary of our 17 computer models of archosaur bodies, shown as one walks along the tips of the phylogeny shown in the video (the models are not considered to be ancestral to one another; we used a common computer algorithm called squared-change parsimony to estimate ancestral state changes of body dimensions between the 16 numbered nodes of the tree):
But we’ve done much more than just put numbers on conventional wisdom.
We’ve shown, to our own surprise, that the shift of the centre of mass was largely driven by evolutionary enlargements of the forelimbs (and the head and neck, and hindlimbs, to a less strong degree), not the tail as everyone including ourselves has assumed for almost 25 years. And the timing of this shift occurred inside the theropod dinosaur group that is called Maniraptora (or Maniraptoriformes, a slightly larger group), so the change began in animals that were not flying, but not long before flight evolved (depending on whom you ask, what taxonomy they favour and what evidence one accepts, either the smaller clade Eumaniraptora/Paraves or the bird clade Aves/Avialae).
Now, if you don’t like the cliche “rewriting the textbooks”, do have a look through texts on dinosaur/early avian palaeobiology and you probably will find a discussion of how the tail shortened, the centre of mass moved forwards as a consequence, the caudofemoral musculature diminished, and theropod dinosaurs (including birds) became more crouched as a result. We did that to confirm for ourselves that it’s a pretty well-accepted idea. Our study supports a large proportion of that idea’s reasoning, but modifies the emphasis to be on the forelimbs more than the tail for centre of mass effects, so the story gets more complex. The inference about caudofemoral muscles still seems quite sound, however, as is the general trend of increased limb crouching, but our paper approximates the timing of those changes.
Figure 3 from our paper, showing how the centre of mass moved forwards (up the y-axis) as one moves toward living birds (node 16); the funny dip at the end is an anomaly we discuss in the paper.
A final implication of our study is that, because the forelimbs’ size influenced the centre of mass position, and thus influenced the ways the hindlimbs functioned, the forelimbs and hindlimbs are more coupled (via their effects on the centre of mass) than anyone has typically considered. Thus bipedalism and flight in theropods still have some functional coupling– although this is a matter of degree and not black/white, so by no means should we do away with helpful concepts like locomotor modules.
And in addition to doing science that we feel is good, we’ve gone the extra mile and presented all our data (yes, 17 dinosaurs’ worth of 3D whole body graphics!) and the critical software tools needed to replicate our analysis, in the Dryad database (link now working!), which should have now gone live with the paper! It was my first time using that database and it was incredibly easy (about 1 hour of work once we had all the final analysis’s files properly organized)– I strongly recommend others to try it out.
That’s my usual general summary of the paper, but that’s not what this blog article is about. I’ll provide my usual set of links to media coverage of the paper below, too. But the focus here is on the story behind the paper, to put a more personal spin on what it means to me (and my coauthors too). I’ll take a historical approach to explain how the paper evolved.
Embarassing picture of me before I became a scientist. Hardee’s fast food restaurant cashier, my first “real job”, from ~1999- no, wait, more like 1986. The 1980s-style feathered (and non-receding) hair gives it away!
Rewind to 1995. I started my PhD at Berkeley. I planned to use biomechanical methods and evidence to reconstruct how Tyrannosaurus rex moved, and started by synthesizing evidence on the anatomy and evolution of the hindlimb musculature in the whole archosaur group, with a focus on the lineage leading to Tyrannosaurus and to living birds. As my PhD project evolved, I became more interested and experienced in using 3D computational tools in biomechanics, which was my ultimate aim for T. rex.
In 1999, Don Henderson published his mathematical slicing approach to compute 3D body dimensions in extinct animals, which was a huge leap for the field forward beyond statistical estimates or physical toy models, because it represented dinosaurs-as-dinosaurs (not extrapolated reptiles/mammals/whatever) and gave you much more information than just body mass, with a lot of potential to do sensitivity analysis.
I struggled to upgrade my computer skills over the intervening years. I was developing the idea to reconstruct not only the biomechanics of T. rex, but also the evolutionary changes of biomechanics along the whole archosaur lineage to birds– because with a series of models of different species and a working phylogeny, you could do that. To me this was far more interesting than the morphology or function of any one taxon, BUT required you to be able to assess the latter. So Tyrannosaurus became a “case study” for me in how to reconstruct form and function in extinct animals, because it was interesting in its own right (mainly because of its giant size and bipedalism). (Much later, in 2007, I finally finished a collaboration to develop our own software package to do this 3D modelling, with Victor Ng-Thow-Hing and F. Clay Anderson at Honda and Stanford)
Me and a Mystery Scientist (then an undergrad; now a very successful palaeontologist!), measuring up a successful Cretaceous hypercarnivore at the UCMP; from my PhD days at Berkeley, ~2000 or so.
In all this research, I was inspired by not only my thesis committee and others at Berkeley, but also to a HUGE degree by Steve Gatesy, a very influential mentor and role model at Brown University. I owe a lot to him, and in a sense this paper is an homage to his trailblazing research; particularly his 1990 Paleobiology paper.
In 2001, I got the NSF bioinformatics postdoc I badly wanted, to go to the Neuromuscular Biomechanics lab at Stanford and learn the very latest 3D computational methods in biomechanics from Prof. Scott Delp’s team. This was a pivotal moment in my career; I became partly-an-engineer from that experience, and published some papers that I still look back fondly upon. Those papers, and many since (focused on validating and testing the accuracy/reliability of computer models of dinosaurs), set the stage for the present paper, which is one of the ones I’ve dreamed to do since the 1990s. So you may understand my excitement here…
Stanford’s Neuromuscular Biomechanics Lab, just before I left in 2003.
But the new paper is a team effort, and was driven by a very talented and fun then-PhD-student, now postdoc, Dr Vivian Allen. Viv’s PhD (2005-2009ish) was essentially intended to do all the things in biomechanics/evolution that I had run out of time/expertise to do in my PhD and postdoc, in regards to the evolution of dinosaur (especially theropod) locomotor biomechanics. And as I’d hoped, Viv put his own unique spin on the project, proving himself far better than me at writing software code and working with 3D graphics and biomechanical models. He’s now everything that I had hoped I’d become by the end of my postdoc, but didn’t really achieve, and more than that, too. So huge credit goes to Viv for this paper; it would never have happened without him.
We also got Karl Bates, another proven biomechanics/modelling expert, to contribute diverse ideas and data. Furthermore, Zhiheng Li (now at UT-Austin doing a PhD with Dr Julia Clarke) brought some awesome fossil birds (Pengornisand Yixianornis) from the IVPP in Beijing in order to microCT scan them in London. Zhiheng thus earned coauthorship on the paper — and I give big thanks to the Royal Society for funding this as an International Joint Project, with Dr Zhonghe Zhou at the IVPP.
That’s the team and the background, and I’ve already given you the punchlines for the paper; these are the primitive and the derived states of the paper. The rest of this post is about what happened behind the scenes. No huge drama or anything, but hard, cautious work and perseverance.
Me shortly after I moved to the RVC; video still frame from a dinosaur exhibit (c. 2004) I was featured in. Embarassingly goofy pic, but I like the blurb at the bottom. It’s all about the evolutionary polarity, baby!
The paper of course got started during Viv’s PhD thesis; it was one of his chapters. However, back then it was just a focus on how the centre of mass changed, and the results for those simple patterns weren’t very different from those we present in the paper. We did spot, as our Nature supplementary information notes, a strange trend in early theropods (like Dilophosaurus; to a lesser degree Coelophysis too) related to some unusual traits (e.g. a long torso) and suggested that there was a forward shift of centre of mass in these animals, but that wasn’t strongly upheld as we began to write the Nature paper.
On the urging of the PhD exam committee (and later the paper reviewers, too), Viv looked at the contributions of segment (i.e. head, neck, trunk, limbs, tail) mass and centre of mass to the overall whole body centre of mass. And I’m glad he did, since that uncovered the trend we did not expect to find: that the forelimb masses were far more important for moving the centre of mass forwards than the mass (or centre of mass) of the tail was– in other words, the statistical correlation of forelimb mass and centre of mass was strong, whereas changes of tail size didn’t correlate with the centre of mass nearly as much. We scrutinized those results quite carefully, even finding a very annoying bug in the 3D graphics files that required a major re-analysis during peer review (delaying the paper by ~6 months).
The paper was submitted to Nature first, passing a presubmission inquiry to check if the editor felt it fit the journal well enough. We had 3 anonymous peer reviewers; 1 gave extensive, detailed comments in the 3 rounds of review and was very fair and constructive, 1 gave helpful comments on writing style and other aspects of presentation as well as elements of the science, and 1 wasn’t that impressed by the paper’s novelty but wanted lots more species added, to investigate changes within different lineages of maniraptorans (e.g. therizinosaurs, oviraptorosaurs). That third reviewer only reviewed the paper for the first round (AFAIK), so I guess we won them over or else the editor overruled their concerns. We argued that 17 taxa were probably good enough to get the general evolutionary trends that we were after, and that number was ~16 more species than any prior studies had really done.
Above: CT scan reconstruction of the early extinct bird Yixianornis in slab conformation, and then Below: 3D skeletal reconstruction by Julia Molnar, missing just the final head (I find this very funny; Daffy Duck-esque) which we scaled to the fossil’s dimensions from the better data in our Archaeopteryx images. There is also the concern, which the reviewers didn’t focus on but I could see other colleagues worrying about, that some of the specimens we used were either composites, sculpted, or otherwise not based on 100% complete, perfectly intact specimens. The latter are hard to come by for a diversity of extinct animals, especially in the maniraptoran/early bird group. We discussed some of these problems in our 3D Tyrannosaurus paper. The early dinosauromorph Marasuchus that we used was a cast/sculpted NHMUK specimen based on original material, as was our Coelophysis, Microraptor and Archaeopteryx; and our Carnegie ??Caenagnathus??Anzu (now published) specimen was based more on measurements from 1 specimen than from direct scans, and there were a few other issues with our other specimens, all detailed in our paper’s Supplementary Information.
But our intuition, based on a lot of time spent with these models and the analysis of their data, is that these anatomical imperfections matter far, far less than the statistical methods that we employed– because we add a lot of flesh (like real animals have!) outside of the skeleton in our method, the precise morphology of the skeleton doesn’t matter much. It’s not like you need the kind of quality of anatomical detail that you need to do systematic analyses or osteological descriptive papers. The broad dimensions can matter, but those tend to be covered by the (overly, we suspect) broad error bars that our study had (see graph above). Hence while anyone could quibble ad infinitum about the accuracy of our skeletal data, I doubt it’s that bad– and it’s still a huge leap beyond previous studies, which did not present quantitative data, did not do comparative studies of multiple species, or did not construct models based on actual 3D skeletons as opposed to artists’ 2D shrinkwrapped reconstructions (the “Greg Paul method”). We also did directly measure the bodies of two extant archosaurs in our paper: a freshwater crocodile and a junglefowl (CT scan of the latter is reconstructed below in 3D).
One thing we still need to do, in future studies, is to look more carefully inside of the bird clade (Aves/Avialae) to see what’s going on there, especially as one moves closer to the crown group (modern birds). We represented modern birds with simply 1 bird: the “wild-type chicken” Red junglefowl, which isn’t drastically different in body shape from other basal modern birds such as a tinamou. Our paper was not about how diversity of body shape and centre of mass evolved within modern birds. But inspecting trends within Palaeognathae would be super interesting, because a lot of locomotor, size and body shape changes evolved therein; ostriches are probably a very, very poor proxy for the size and shape of the most recent common ancestor of all extant birds, for example, even though they seem to be fairly basal within that whole lineage. And, naturally, our study opens up opportunities for anyone to add feathers to our models and investigate aerodynamics, or to apply our methods to other dinosaur/vertebrate/metazoan groups. If the funding gods are kind to us, later this year we will be looking more closely, in particular, at the base of Archosauria and what was happening to locomotor mechanics in Triassic archosaurs…
Clickum to embiggum:
Australian freshwater crocodile, Crocodylus johnstoni; we CT scanned it in 3 pieces while visiting the Witmer lab in Ohio.
A Red junglefowl cockerel, spotted in Lampang, Thailand during one of my elephant gait research excursions there. Svelte, muscular and fast as hell. This photo is here to remind me to TAKE BLOODY PICTURES OF MY ACTUAL RESEARCH SPECIMENS SO I CAN SHOW THEM!
I’d bore you with the statistical intricacies of the paper, but that’s not very fun and it’s not the style of this blog, which is not called “What’s in John’s Software Code?”. Viv really worked his butt off to get the stats right, and we did many rounds of revisions and checking together, in addition to consultations with statistics experts. So I feel we did a good job. See the paper if that kind of thing floats your boat. Someone could find a flaw or alternative method, and if that changed our major conclusions that would be a bummer– but that’s science. We took the plunge and put all of our data online, as noted above, so anyone can do that, and that optimizes the reproducibility of science.
What I hope people do, in particular, is to use the 3D graphics of our paper’s 17 specimen-based archosaur bodies for other things– new and original research, video games, animations, whatever. It has been very satisfying to finally, from fairly early in the paper-writing process onwards, present all of the complex data in an analysis like this so someone else can use it. My past modelling papers have not done this, but I aim to backtrack and bring them up to snuff like this. We couldn’t publish open access in Nature, but we achieved reasonably open data at least, and to me that’s as important. I am really excited at a personal level, and intrigued from a professional standpoint, to see how our data and tools get used. We’ll be posting refinements of our (Matlab software-based) tools, which we’re still finding ways to enhance, as we proceed with future research.
Above: Two of the 17 archosaur 3D models (the skinny “mininal” models; shrinkwrapped for your protection) that you can download and examine and do stuff with! Dilophosaurus on the left; Velociraptor on the right. Maybe you can use these to make a Jurassic Park 4 film that is better, or at least more scientifically accurate, than Hollywood’s version! 😉 Just download free software like Meshlab, drop the OBJ files in and go!
Now, to bring the story full circle, the paper is out at last! A 4 year journey from Viv’s PhD thesis to the journal, and for me a ~14 year journey from my mind’s eye to realization. Phew! The real fun begins now, as we see how the paper is received! I hope you like it, and if you work in this area I hope you like the big dataset that comes with it, too. Perhaps more than any other paper I’ve written, because of the long voyage this paper has taken, it has a special place in my heart. I’m proud of it and the work our team did together to produce it. Now it is also yours. And all 3200ish words of this lengthy blog post are, as well!
Last but not least, enjoy the wonderful digital painting that Luis Rey did for this paper (another of my team’s many failed attempts to get on the cover of a journal!); he has now blogged about it, too!
Dinosaur posture and body shape evolving up the evolutionary tree, with example taxa depicted. By Luis Rey.
News stories about this paper will be added below as they come out, featuring our favourites:
Synopsis: Decent coverage, but negligible coverage in the general press; just science-specialist media, more or less. I think the story was judged to be too complex/esoteric for the general public. You’d think dinosaurs, evolution, computers plus physics would be an “easy sell” but it was not, and I don’t think we made any big errors “selling” it. Interesting– I continue to learn more about how unpredictable the media can be.
Regardless, the paper has had a great response from scientist colleagues/science afficionados, which was the target audience anyway. I’m very pleased with it, too– it’s one of my team’s best papers in my ~18 year career.
If you want to see a new/reinvigorated, exciting direction that palaeoart is headed, check out the All Yesterdays book by Conway, Kosemen and Naish. This review is fully cognizant that I’m late to the party of hailing this book as part of a palaeoart renaissance. I confess I haven’t read any of the many reviews of this book; I just know it is highly regarded and popular, from excitement on social media sites I frequent. So if my review covers ground others have too, so be it; it’s purely my own thoughts but I expect that mine fall in line with many others’. I’m reviewing the book on this blog because I love the interface between science and art (which is very important in anatomy), and because anatomy, and how one infers it when it is unknown, is the fundamental theme of the book.
You can buy All Yesterdaysfor around £18 (ASIDE: oddly, used copies (“May not include CD, access code, or DJ”– ???) are around £42 on the same site; perhaps those are artist-signed??? I have no idea!). It is a good deal at that price. While you’re at it, get “Dinosaur Art: The World’s Best Paleoart” by White et al. (including Conway) for a similar price. My review will return to some comparisons between these two books, released just a few months apart.
All Yesterdays is about not only how we reconstruct dinosaurs and other prehistoric animals, but also about thinking outside-the-box in the ways we reconstruct them and thereby bucking some recent clichés and tropes. Some of those outside-the-box ideas might seem ludicrous, and some probably are. But one of the main points of All Yesterdays is that there is plenty about extinct animals, and even living animals, that we don’t know, even though the field of paleoart has matured into greater scientific rigor than in the days of Knight, Zallinger and others (1920s-1950s). There is a focus on uncertainties about integument (e.g. feathering, spines, colour/patterning, body contours) and behaviour (e.g. avoiding stereotypes like perpetually aggressive predators and frightened prey animals– amen to that!). And the capstone of the book, which in some ways I loved the most, is turning the issue on its head and pretending that we only had skeletons of extant animals, then proceeding to reconstruct those animals (elephants, whales, horses and swans stand out prominently in this section; some of these are shown below). I wish more scientists in my general area would practice this; e.g. validation of a methodology used to reconstruct extinct animals in science.
The ‘speculative zoology’ of All Yesterdays deserves favourable comparison to one of my favourite science-art books, 1981’s After Man by Dougal Dixon. I fell so in love with that book as a 10-year-old that I wouldn’t let my parents return it to the library and I made them pay the hefty lost-book-fee (yes, I was a little bastard!). I still have it, too. (Sorry, Sequoya Branch Public Library of Madison, WI!) Likewise, the whimsy of the Rhinogrades is evoked by this work, and of course Tetrapod Zoology blog readers will be no strangers to it, either.
The book begins with a clear, succinct (7 page) summary of the history and science of reconstructing animals, with a focus on paleoart’s approach rather than science’s. I would have found it interesting (but space constraints presumably precluded) to feature more of the interface/parallels with scientists at the same time, such as the careful reconstructions of musculature in A.S. Romer’s masterful work in the 1920s (e.g. below), or later efforts by palaeontologists like Alick Walker and Walter Coombs. Many of these luminaries sought not to reconstruct animals for artistic purposes, but for almost purely scientific ones: to understand what skeletal anatomy meant in terms of broader biology (e.g. comparative anatomy) and phylogeny (e.g. origin of birds or archosaur evolution). The quality of their own artistic representations as well as scientific interpretations varied a lot. Indeed, sometimes the choice of model organisms (crocodile for Romer; lizard for Walker; birds in the post-1960’s) reveals much about the author’s preconceptions about phylogeny, marshalled towards a favoured hypothesis (e.g. a crocodile origin of birds for Walker; or an avian origin amongst dinosaurs for Bakker, Paul and others), rather than a circumspect assessment of all relevant evidence.
Figure 6 from Romer, 1923; very crocodylian T. rex right hindlimb muscles.
But eventually the “model organism” approach to reconstructing extinct animals gave way to the extant phylogenetic bracket; very popular today; which itself is an adaptation of the outgroup method for polarity assessment in phylogenetic systematics (cladistics). I am sure many modern paleoartists explicitly consider the “EPB” in their reconstructions, although this leaves many ambiguities (e.g. integument of crocodiles and birds being totally different!) that they must overcome, whereas scientists might just give up. This interface of art and science is part of what make palaeontology so enjoyable.
The EPB mindset has been a big step forward for evolutionary morphology and palaeontology, but still some of the greatest questions (e.g. what were the actual sizes, colour patterns, or behaviours of extinct animals? How did novelties arise and which novelties did dinosaurs have that extant relatives lack?) are left ambiguous by the EPB. This is because either the EPB itself is ambiguous (crocodiles or other taxa do one thing; birds do something altogether different), or because features leave no osteological correlates (e.g. muscle/tendon/ligament scars) on fossils that can be compared with the EPB. This quandary leads to the fun side of this book– filling in the huge gaps left by both basic anatomical interpretation and the restrictions imposed by the EPB, and then playing with the frontiers of anatomical, behavioural and ecological reconstruction, using informed speculation.
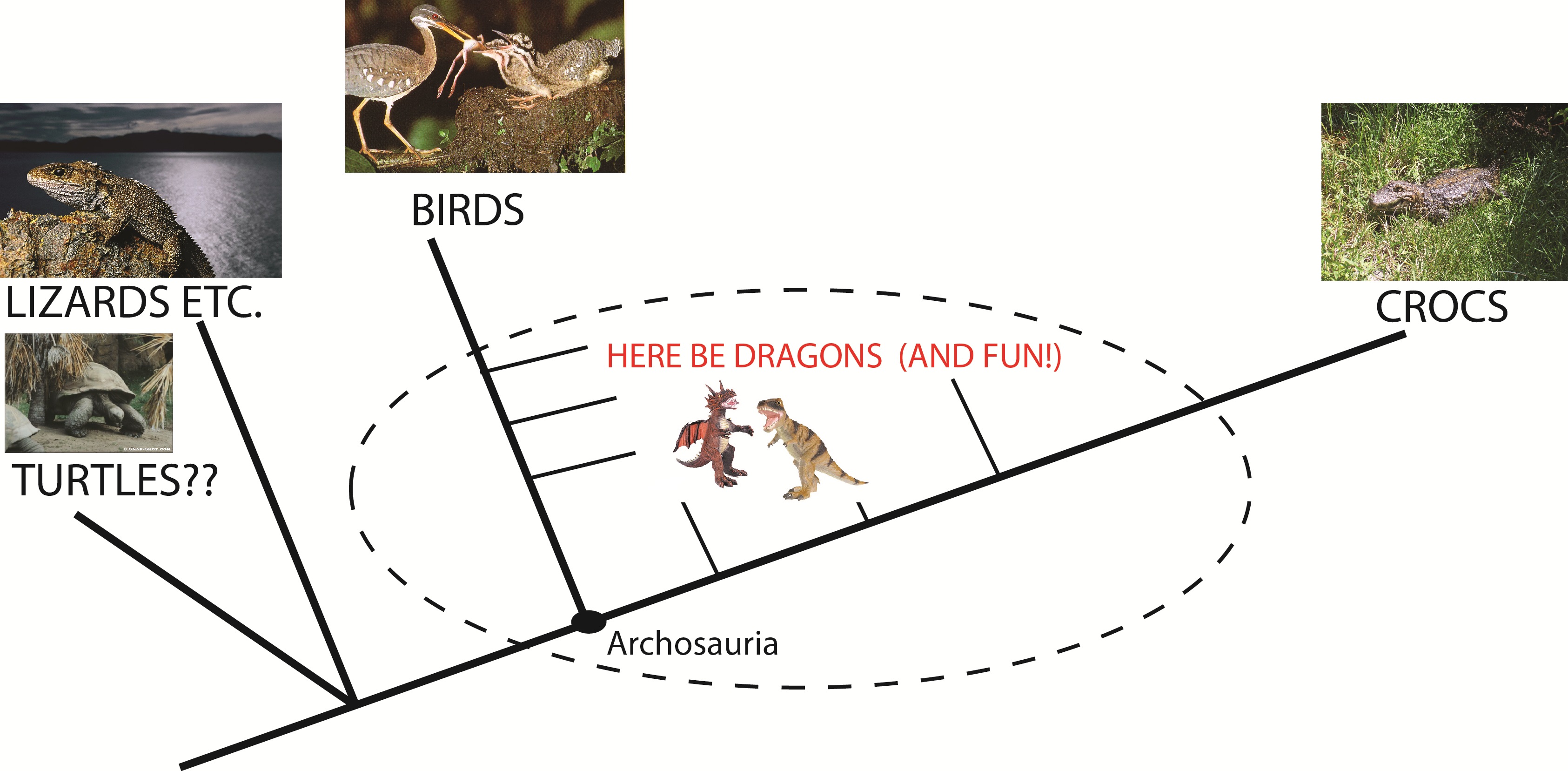
The extant phylogenetic bracket for archosaurs.
In addition to the startling, bizarre “All Todays” reconstructions at the end of the book, the highlights for me were the camouflaged Majungasaurus and plesiosaur, the “feathered mountain” (below) of a therizinosaur (can anyone illustrate a plausible therizinosaur and make it normal and boring? I wager not!) and the neck-swinging elasmosaurs engaged in “honest signalling” of their fitness. Many of the illustrations riff on notions popular in the modern palaeo-zeitgeist (and subject of many conversations at conferences, or even publications), such as evidence for the spiny integument of some ornithischians, fat ornithopods, Microraptor of somewhat-known-colouration, and so on. But plenty of other images riff on a “well why not?” theme, challenging the viewer to consider that extinct animals could have many surprises left in store for us with future discoveries, or else plausible features that we’ll never know of but might seem laughable or unfashionable to illustrate now. Each image has text explaining the logic behind it- this is not just a montage of pictures. This is a thinking person’s book- you should buy it for rumination, to challenge your preconceptions, not to have a flashy coffee table book. It’s not eye candy — it’s more like brain jerky.
John Conway‘s mountain-of-feathers therizinosaurs: eerily beautiful.
I think this is a bold, fun (re)new(ed) direction for palaeoart. There’s always a place for rigorous, conservatively evidence-based, by-comparison-almost-uncreative scientific illustration of extinct organisms. The World’s Best Paleoart presents loads of this, often in vividly colourful, photo-realistic, lavish, glossy detail, whereas the approach in All Yesterdays tends toward a more soft, matte, informal style including sketches or abstractions, toning down the serious and intense (even cluttered?) approach that can characterize modern palaeoart, including The World’s Best Paleoart. Sometimes those reconstructing life of the past (scientists included!) may emphasize that detailed realism too much and lose some of the joyful playfulness that palaeoart can revel in, at its best, most inspirational or thought-provoking. The former style might be considered the more “safe” or technical practice; the latter more risky or unconstrained.
C.M. Kosemen‘s “All Todays” swans, with tadpolefish, might haunt your nightmares.
I’m not casting negative judgement on either style; both are absolutely wonderful — and valuable. I love both books! I’m glad we’re in a new age where the fun is waltzing back into palaeoart, that’s all. All Yesterdays doesn’t just waltz, either. It pounces into your field of view, wiggles its rainbow-coloured, mandrill-esque ankylosaurid bottom at you with a cheeky grin, and proceeds to make you smirk, be bemused, and even gasp at its adventurousness in rapid succession as you turn its pages. At 100 pages it doesn’t overstay its welcome either– that kaleidoscopic thyreophoran rump cartwheels off into the sunset at an opportune moment.
You won’t forget C.M. Kosemen‘s “All Todays” elephant.
If All Yesterdays makes someone uncomfortable with its swashbucklerish daring, they’re probably taking palaeontology way too seriously– and maybe missing not only some good fun, but also some potential truths. Dogma is a terrible thing, and All Yesterdays slaughters it with delightful relish. Bring on the next installment! If you have All Yesterdays too, what’s your favourite part? Or if you don’t have it, I’d be happy to answer queries in the Comments.
I’m letting the dogs out today. Science gone barking mad! Hopefully my puns will not screw the pooch.
Stomach-Churning Rating: 4/10; a dog cadaver’s leg (not messy), then just tame digital images of anatomy.
I am working with Rich Ellis, a former MSc student at Univ. Colorado (see his cool new paper here!), for a fun new collaboration this year. He was awarded a prestigious Whitaker Foundation scholarship to do this research, which focuses on how different animals stand up from a squatting position, with the legs about as bent as they can be.
We want to know how animals do this standing up movement, because it is in some ways a very demanding activity. Very flexed/bent limb joints mean that the muscles (and some tendons) are stretched about as far as they ever will be. So this places them at disadvantageous lengths (and leverage, or mechanical advantage) for producing force. We know almost nothing about how any animal, even humans, does this-– how close to their limits of length are their muscles? Which muscles are closest? Does this change in animals with different numbers of legs, postures, anatomy, size, etc? Such fundamental questions are totally unaddressed. It’s an exciting area to blaze a new trail in, as Rich is doing. So far, we’ve worked with quail, humans, and now greyhounds; in the past I did some simple studies with horses and elephants, too. Jeff Rankin from my team and other collaborators have also worked on six species of birds, of varying sizes, to see how their squat-stand mechanics change. Thus we’ve covered a wide diversity of animals, and now we’re learning from that diversity. “Diversity enables discovery,” one of my former PhD mentors Prof. Bob Full always says. Too true.
Greyhounds are interesting because they are medium-sized, long-legged, quadrupedal, quite erect in posture, and very specialized for fast running. Fast runners tend to have big muscles with fairly short fibres. Short fibres are bad for moving the joints through very large ranges of motion. So how does a greyhound stand up? Obviously they can do it, but they might have some interesting strategies for doing so- the demands for large joint motion may require a compromise with the demands for fast running. Or maybe the two demands actually can both be optimized without conflict. We don’t know. But we’re going to find out, and then we’ll see how greyhounds compare with other animals.
To find out, we first have to measure some dogs standing up. We’ve done that for about 8 greyhounds. Here is an example of a cooperative pooch:
Those harmless experiments, if you follow me on Twitter, were live-tweeted under the hashtag #StandSpotStand… I dropped the ball there and didn’t continue the tweeting long after data collection, but we got the point across– it’s fun science addressing useful questions. Anyway, the experiments went well, thanks to cooperative pooches like the one above, and Rich has analyzed most of the data.
Now the next step involves the cadaver of a dog. We could anaesthetize our subjects and do this next procedure to obtain subject-specific anatomy. But it really wouldn’t be ethically justified (and if I were an owner I wouldn’t allow it either!) and so we don’t. A greyhound is a greyhound as far as we’re concerned; they’ll be more like each other than either is like a quail or a human. Individual variation is a whole other subject, and there are published data on this that we can compare with.
We get a dead dog’s leg — we don’t kill them; we get cadavers and re-use them:
We study the hindlimb because birds and humans don’t use their forelimbs much to stand up normally, so this makes comparisons simpler. We’re collecting forelimb data, though, as we work with quadrupeds, for a rainy day.
We then CT scan the leg, getting a stack of slices like this– see what you can identify here:
It’s not so clear in these images, but I was impressed to see that the muscles showed up very clearly with this leg. That was doggone cool! Perhaps some combination of formalin preservation, fresh condition, and freezing made the CT images clearer than I am used to. Anyway, this turned out to be a treat for our research, as follows.
We then use commercial software (we like Mimics; others use Amira or other packages) to “segment” (make digital representations in 3D) the CT scan data into 3D anatomy, partitioning the greyscale CT images into coloured individual objects– two views of one part of the thigh are shown below.
What can you identify as different colours here? There are lots of clues in the images (click to embiggen):
And here is what the whole thigh looks like when you switch to the 3D imaging view:
Quite fetching image, eh?!
The next steps after we finish the limb segmentation are to apply the experimental data we observed for greyhounds of comparable size by importing the model and those data into biomechanics software (SIMM/OpenSim). We’ve done about 40 models like this for various species. I detailed this procedure for an elephant here.
Then, at long last, science will know how a greyhound stands up! Wahoo! Waise the woof! Stay tuned as we hound you with more progress on this research-as-it-happens. Rich just finished the above thigh model this week, and the rest of the leg will be done soon.
Many thanks to Rich Ellis for providing images used here. And thank you for persevering my puns; they will now be cur tailed.
Happy Freezermas! Sing it: “On the fifth day of Freezermas, this blo-og gave to me: one tibiotarsus, two silly Darwins, three muscle layers, four gory hearts, a-and five stages modelling a doggie!” ♪♫
Party time! Let the media onslaught begin! We’ve published a paper in Nature on the limb motions of Ichthyostega (and by implication, some other stem tetrapods). Since we did use some crocodile specimens from Freezersaurus (see below) in this study, I figured WIJF could cover it to help celebrate this auspicious event. Briefly. Particularly since we already did a quasi-blog on it, which is here:
However I want to feature our rockin’ cool animations we did for the paper, to squeeze every last possible drop of science communicationy goodness out of them. So here they are in all their digital glory. Huge credit to Dr. Stephanie Pierce, the brilliant, hardworking postdoc who spearheaded the work including these videos! Dr. Jenny Clack is our coauthor on this study and the sage of Ichthyostega and its relatives- her website is here. Also, a big hurrah for our goddess of artsy science, Julia Molnar, who helped with the videos and other images. Enjoy!
The computer model
The forelimb model
The hindlimb model
We used some of my Nile crocodile collection to do a validation analysis of our joint range of motion (ROM) methods, detailed in the Supplementary info of the paper, which I encourage anyone interested to read since it has loads more interesting stuff and cool pics. We found that a bone-based ROM will always give you a greater ROM than an intact fleshy limb-based ROM. In other words, muscles and ligaments (and articular cartilage, etc.). have a net effect of reducing how far a joint can move. This is not shocking but few studies have ever truly quantitatively checked this with empirical data from whole animals. It is an important consideration for all vert paleo types. Here is a pic of one of the crocodiles from the study, with (A) and without muscles (B; ligaments only):
I’ll close with Julia Molnar’s jaw-droppingly awesome flesh reconstruction from our model. Why Nature wouldn’t use this as a cover pic, I’ll never understand, but I LOVE it! When I first saw it enter my email inbox and then opened it to behold its glory, my squeal of geeky joy was deafening.
(edit: Aha! Fellow Berkeley alum Nick Pyenson’s group made the Nature cover, for their kickass study of rorqual whale anatomy, including a “new” organ! Well, we don’t feel so bad then. Great science– and a win for anatomy!!!)
This post will walk through the basic steps we take to do some of the major, ongoing research in my team. It comes from our lengthy project aiming to determine how elephant legs work at the level of individual muscle/tendon/bone organs. We need fancy computer simulations because anatomy, mechanics, physiology, neural control etc. are all very complex and not only impossible to completely measure in a living, moving animal but also extremely unethical and unjustified in the case of a rare, fragile animal like an Asian elephant. We want to do such complex things to test hypotheses about how animals work. For example, we want to estimate how fast an elephant could run if it wanted to, or why they cannot (or will not) jump or gallop like smaller mammals do— even as baby elephants (~100 kg or 220 lbs), which is an ancillary question we’re tackling. That’s cool basic science, and that’s enough for me. But the applications once such models and simulations are established are manifold– human clinical research now routinely employs such approaches to help treat “crouch gait” in patients with cerebral palsy, plan corrective surgeries, aid in rehabilitation strategies, and even potentially optimize athletic performance. Non-human research is pretty far behind this kind of confident application, because there are too damn many interesting non-humans out there to study and not many people using these approaches to study them (but it’s catching on).
Breaking up the monotony of the text with a baby elephant we met during our research in Thailand (Chiang Mai, here) in 2001. It was just a few days old and VERY cuddly and playful (chewing on everything!) but it’s mother did not want us playing with it so we only gave a quick hello.
I use the term model to refer to a simple abstraction of reality (such as an anatomically realistic computer graphic of a limb), and a simulation as a more complex process that is more open-ended and generally uses a model to ask a question (such as what level of extreme athletic behaviour a modelled limb could support). We use models and simulations to test how all the structures of the limb work together to produce movement. This also reciprocally gives us insight into the question, as I like to say it, of why is there anatomy? What is anatomy for? Why does it vary so much within so many groups and not so much in others? This can more easily be addressed by focusing on the consequences of a given anatomy rather than the more tricky question of why it evolved.
These approaches also can answer the frightening question of “Does anatomy really matter?” Sometimes it does not. And those “sometimes” can be impossible to predict- although sometimes they can be easy to predict, too. I think we are not at a point in the maturity of biomechanics/functional morphology to usually know a priori when either is the case. Many factors in addition to anatomy determine function, behaviour, or performance; that’s why; and biomechanics aims to unravel those relationships. A lot of anatomists, palaeontologists, etc. assume that form can be reliably used to predict function, but plenty of studies have shown already (and if you peer deeply into the details, it comes from first principles) that one cannot be sure without either measuring what anatomy is doing in a particular behaviour or estimating that function in a computer model or simulation.
Anyway, I’ve covered my perspective on this in a paper which you can read if you want to go into deep philosophical details of the science (and read me blabbering on more about this particular hobby horse of mine?). This post will proceed mostly with pretty images and simple explanations, although I welcome comments and queries at the end. As part of this post, I’ll try to give an idea of the timespans involved in doing the research. Some steps are quick and easy; others can take dauntingly long — especially to do well, without building a digital house of cards.
I’ll start, as my posts often do, with a deceased animal, and in this case it will again be an Asian elephant. Incidentally it is the same animal from the “Inside Nature’s Giants” series (see previous post).
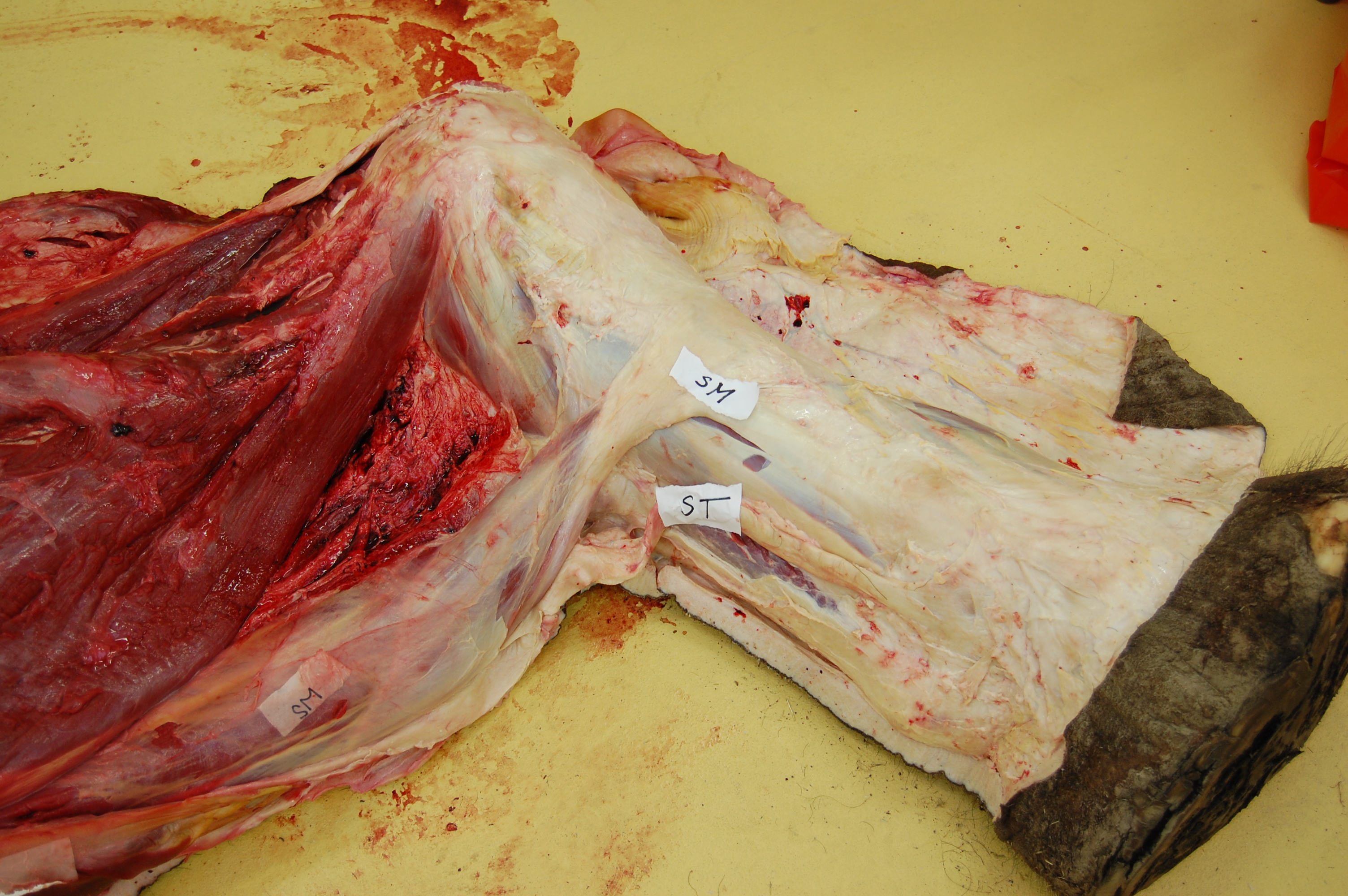
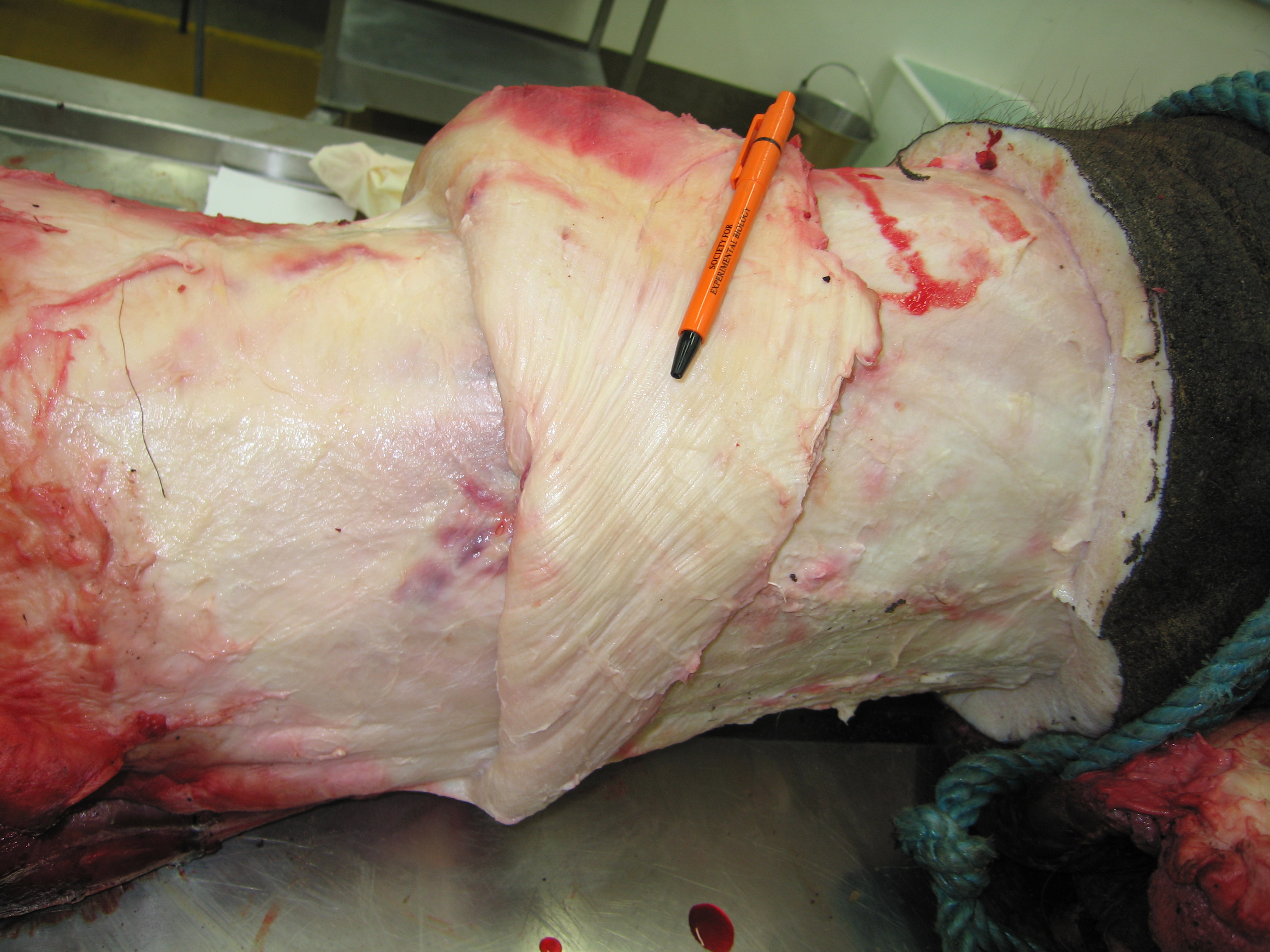
Above: the hindlimb viewed from the rear, showing the medial (inside) region of the thigh skinned down to the superficial musculature. The hip is toward the left of the screen, and the knee is to the far right (whitish rounded area), with the shank (still bearing most of its grey hide) heading to the bottom right corner of the picture. Muscles pictured include ST (semitendinosus) and SM (semimembranosus); major hamstring muscles; as well as the thin, sheet-like gracilis, the straplike sartorius, and the massive adductors toward the top of the image.
When collecting data from dissections for functional analysis including computer models and simulations, we dissect the muscles one by one as we identify and photograph/sketch them, then remove them and do a suite of measurements to characterize how their form relates to some basic functional parameters. From the mass (weight) of the muscle and the length and angulation (pennation) of its fibres (bundled as fascicles) we can estimate what is called the physiological cross-sectional area (PCSA) of each muscle, which is known to strongly correlate with the force the muscle can produce. Different muscles have different PCSAs; for example check out these pictures of a long-fibred, lower-PCSA muscle and a short-fibred, highly pennate and high PCSA muscle:
Above: the long muscle fibres (bands running from left to right, somewhat diagonally from the bottom left corner toward the top right) of a hip adductor muscle in our specimen. The adductors are fairly simple muscles that run from the underside of the pelvis to the inside of the thigh (femur).
Above: the tensor fasciae latae (TFL; pretty sure of ID but going from memory) hip muscle of our specimen, cut open to show the short, angled fibres (each leading at around a 45 degree angle to attach onto a thick central internal tendon). The TFL is just out of view at the top of the screen in the whole leg anatomy picture above; it is on the front outer, upper margin of the hip/thigh and runs down to the outer side of the knee, invested with thick sheets of connective tissue (fascia).
The maximal isometric force (Fmax) of a muscle is computed as the PCSA times the muscle stress (force/unit area), which is fairly conservative in vertebrates. A square meter of PCSA can produce around 200-300 kilonewtons of force, or about 60,000 cheeseburger-weights (the standard unit of force on this blog). That’s a lot of quarter pounders! And an elephant has pretty close to that many cheeseburgers worth of leg muscle (around 150 kg mass, very close to a square meter of PCSA; total Fmax would be around 80,000 cheese-burger weights!). That much muscle is important because an Asian elephant like this one weighed 3550 kg or about 9000 cheeseburger-weights. So if all the muscles in one elephant hindlimb could push in one direction at once, in theory they could hold about 9 elephants aloft. However, as the picture above shows, they do not all act in the same direction. Furthermore, there are many other factors involved in determining how hard a leg can push, such as the leverage of the muscle forces versus the actions of gravity and inertia (mechanical advantage). All those factors, again, are why we need computer models to address the complexity. But the end result is that elephants cannot support 9 times their body weight on one hind leg.
Enough talk about cheeseburgers and enough possibly savory pictures of giant steak-like leg muscles. I don’t want to be blamed for hunger-induced health problems in my beloved blog-readership, dear Freezerinos! The above steps take about a week to complete for 2 legs of a big elephant, rushing against decomposition to try to get the best quality data we can. On to the digital stuff- let’s turn the geekitude dial up to 11 with some videos of computer modelling.
Our next step, often featured on this blog because I do this so often, is to take CT (and/or MRI) scans of the specimen that we wisely did before we cut it to bits, and use those to make a computer model. That’s the easy step; a scan nowaways takes me less than an hour to complete, including moving the specimen back and forth between the freezer and imaging centre. MRI scans can take quite a bit longer. Here is a CT scan of a similar hindlimb (right leg for the toes up to the knee, from a juvenile elephant; the above leg was too big for our scanner!). See what you can identify here:
And then here is a resulting computer model of the same animal (just knee down to toes), showing how we took each CT slice of even the muscles and turned them into fully or partially 3D digital organs, in our case using commercial software that makes this procedure (a step called segmentation) very easy:
The segmentation step for bones is usually incredibly simple; it can take anywhere from an hour to a day or so, depending on anatomical complexity and image quality. For muscles, this is harder because the images are often more hazy and muscles tend to interweave with each other, segue into tiny tendons, take sudden turns through bones or other narrow spaces, or even fuse with other muscles. So when we do this kind of musculoskeletal modelling, it gets pretty laborious, and can take weeks or months to finish.
Ahh, but once you’re done with the basic anatomy, the real fun begins! We take the 3D images of bones, muscles, etc. and import them into our biomechanics software. We use two packages: one commericial item called SIMM (Software for Integrative Musculoskeletal Modeling) for making models, and a nice freebie called OpenSim for doing simulations (although actually we’re finding SIMM is often better at doing both modelling and simulation for more unusual animals). Quite a bit more anatomical work is required to get the joints to move properly, then position the muscles in accurate or at least realistic 3D paths (depending on segmented image quality), then check the muscles to ensure they move properly throughout the joints’ ranges of motion, then import all the PCSA and Fmax and other data we need from dissections, then do a lot more debugging of the model… this takes months, at least.
But the greatest joy and pain comes in getting the biomechanics done with the models and simulations. You can get quite simple data out of the models alone; such as the leverages (moment arms) of individual muscles and how these change with limb joint position, across a gait cycle, etc… That’s pretty interesting to us, and can just take a few days to crank out from a finished model. Yet the ultimate goal is to do either a tracking simulation, in which we make the model try to follow forces and motions that we measured in experiments from the same or a similar animal (standard, harmless gait analyses), or a theoretical simulation, in which we set the model a task and some rules (‘optimization criteria’) and then set it to run (for hours, days or weeks) to solve that task while following the rules. In both cases, the simulations estimate the muscle activation timings (on/off and intensity) and forces, as well as the kinematics (motions) and kinetics (forces) of the limbs. Then we check the results, play around with the inputs (unknown parameters) as part of a sensitivity analysis, and re-run the analyses again, and again, and again… Here is a draft of a tracking simulation we’ve run for our elephant’s hindlimb:
Above: again, a right hindlimb of an Asian elephant. This test of our tracking simulation is replicating real experimental data (from motion capture and force platform analysis) of an elephant running at near its top speed; over 4 meters/second (>10 mph/16kph). The red lines are the individual muscles, and the green arrow is the ground reaction force, equal and opposite to the force that the limb applies to the ground. In a fast elephant that force can exceed the elephant’s body weight, so the muscles need to crank out kilo-cheeseburger-units of force!
And that’s about as far as I’ll get today. My team’s previous research (explore links for some fun videos) has shown that elephants can run about 7 meters/second (~15mph; 24kph) and that they have pretty poor mechanical advantage when they do run, so their muscles must have to work pretty hard (about 6 times more cheeseburger units in a fast run vs. a slower walk). So how do they do it? And what prevents them from going faster? What would happen if they jumped? What limits speed more; muscles, tendons or bones? Stay tuned. I’m still not sure how much longer this final step of the research will take… (presumably will precede the heat death of the universe by a long shot) But overall, the whole process when everything works nicely can take a year or so to do, proceeding from whole limbs to a simulated limbs.
As a final teaser, here is work we’ve done on using a different kind of model, called finite element analysis (FEA), to estimate how many cheeseburgers it would take to break an elephant’s femur (thigh bone), for example. How “overbuilt” are bones vs. muscles or tendons? This is still a poorly resolved question in biology. We’ve established some rigorous methodology for doing this, now we just need to see what answers it gives us…
(the colour shows the strain (deformation) in the bone in a simple bending experiment; “hot” colours are higher strain. The visualization of the strain is greatly exaggerated; in the real results they are barely visible, as bone only bends a tiny amount before fracturing)
We use Mimics software, which is pricey but sooooo easy to do stuff like this. Here I’ve just had a very quick pass at reconstructing the distal hindlimb (limb except the thigh, which had bad scan artifacts). With more effort, I could remove the pesky artifacts around the knee and ankle (2 uppermost joints), although some of those are unavoidable because the leg was too damn big for the CT scanner gantry (70cm diameter; leg was around 60cm across at largest).
So how long did this process take? The hardest part was moving the leg around and positioning it on the CT table. Then the scan (608 cross-sectional slices) took about 15 minutes to do, then 15 more minutes to transfer over to my PC. Loading the DICOM slices of data and making a movie (previous post) took 5 minutes, and then making this 3D reconstruction movie took just another 5 minutes, although I waited a few days because I was busy.
So, when operating at peak efficiency, we can obtain decent 3D models from frozen specimens in less than an hour. This is but one example of how modern technology, especially X-ray computed tomography and computer hardware/graphics software, have massively transformed any research that deals with anatomy. When I was doing my PhD back in the 90’s, this would have been a much more time-intensive procedure (probably weeks of work, and difficulties getting CT access); in the 80’s it would basically have been impossible.
In the future, now, we’ll be using these models (once cleaned up a bit) with data from dissection to model how the limbs work in a real giraffe. More about that later. The Giraffe-A-Thon is over for now. I hope you enjoyed it! More of the same to come on this blog!