Stomach-Churning Rating: 5/10; links show some of my health problems, and discuss them in not-always-so-uplifting detail.
I’ve spent almost 10 years now blogging about my health problems, and it has been nearly 20 years since I had a stroke that changed my life and kickstarted a slow landslide of health issues and declining mental capacity. I have fully integrated being a disabled person into my personal identity. There’s hardly a day that goes by in which I don’t think about how I am struggling with my chronic health problems, and the lack of control involved in them. On the brighter side, I’ve taken up the challenge to wave the flag for this identity. I want to do more; to help others, maybe as a role model or beacon of hope or inspiration (you can move on after disaster, while learning from it), and to show that we’re not all alone even though our problems are unique. I am looking for new ways to reach out to the world and do more good with the badness I’ve endured. I’ve done recent interviews and invited talks on disability and science careers/life, which have been a good start. And another intent is to use this summary to direct others to hear my story, more efficiently disseminating it.
I’ve also thought a lot about what it means to be invisibly disabled and thus minoritised, and as a senior white male of no small privilege. How do those two things balance in determining where I fit into the multidimensional space of equality, diversity, equity and inclusiveness? I haven’t had an epiphany to provide an answer, but things certainly have changed for me. The John of 10 years ago is very different from the one of today. Being visible about being invisible helps me, and can benefit others.
This post is an ongoing summary of my experiences, and a retrospective (in chronological order):
What’s In John’s Brain?– 16 December, 2012: The post that started it all, on the 10th anniversary “Not Dead Yet Day” of my stroke.
Shouldering the Burden of Uncertainty– 9 November, 2014: I feel that this is my finest moment in blogging and being up-front with my problems. And vulnerable. It was a very, very hard time in my life that changed everything. But I am proud that I wrote this. Marvel MCU, I am still available.
Life as a Diminishing Scientist– 22 April, 2016: In which I discuss my acceptance that I am disabled, and what that means for my life.
Epilepsy Epilogue– 1 June, 2016: What it’s like “getting used to” epilepsy.
Year 10.5 of John’s Freezer: WTF?– 20 August, 2022: Resurgence of health problems during the continued challenges of the pandemic. Another very hard year. But not all bad.
There will be more writings to come! Hopefully with a turn for the better.
Stomach-Churning Rating:1/10 unless I change my mind and add some unsavoury images (which I did not).
A most un-creative title for a most un-creative post topic. I’ve been short on new ideas for the blog for a long time, and short on energy for them. Furthermore, I’ve been more busy trying to get specimens out of my freezers and reduced to bones, rather than getting new specimens and doing new anatomical work on them, which might otherwise be good fodder for blog posts. I’d love to do a new “Better Know a Muscle” post, but that is hard work. Yet a good old standby post, a summary of stuff I’ve coauthored lately that grabs my immediate fancy, is fine. So here is that summary; most recent papers first.
1. Hutchinson, J.R., Pringle, E.V. 2024. Footfall patterns and stride parameters of Common hippopotamus (Hippopotamus amphibius) on land. PeerJ 12:e17675. https://doi.org/10.7717/peerj.17675
This one got a ton of media attention, much more than I expected. And in a week that was far from slow news (UK election, ever-crazier USA politics, so on). I think the media liked a fun distraction that reminds us how the natural world still has its wonders and how scientists are still doing goofy stuff. I’ve long wanted to do a study of hippo locomotion, as part of my opus on how giant land animals move, so I recruited vet undergrad Emily Pringle to go film hippos at Flamingo Land Resort in North Yorkshire (who’ve been immensely supportive of our research). She set up GoPro cameras and filmed what she could over 2 days. It wasn’t much. The two hippos just walked slowly in and out of their night-time barn now and then. But we had a cunning plan. Emily also searched the internet (mostly Youtube) for more videos showing a wider range of speeds, and I added some more after her project. The upshot is, hippos only use trotting gaits (diagonal limbs moving in near-unison) from slow to fast speeds (walking trots to running trots), and at fast speeds they go airborne for substantial periods of time. That’s cool for two reasons: (1) it’s odd for a (large) mammal to just trot and not use the usual walk-trot-canter-gallop series of gaits; and (2) it wasn’t known (or even asked, much) in scientific literature if hippos did go airborne or not.
My favourite sequence, of a really fast trot with many airborne periods.
I cringe a little at how simple this work is, but there’s still great value in observation and a “data paper” as I regarded this one. Still, I like it and it fills a lacuna in our understanding, and takes my research a step further toward what I’d like to know (e.g., what constraints influence locomotion at giant sizes, and how those evolved in particular lineages). I’ve never written a paper so quickly. It took maybe a week from start to submission. It’s not rocket science, but it’s useful. It has gotten me more motivated to expand this hippo research into some novel directions. A little more about the paper on Twitter.
Speaking of data, here are some. Duty factor (DF; fraction of stride cycle that a given foot is in contact with the ground; i.e., 1 means the feet never ever leave the ground; 0 means that the animal never touches the ground and thus is truly flying) on the x-axis, and stride frequency (SF; stride cycles per second) on the y-axis. So trotting hippos swing their limbs <1 cycle/second at low DFs (~slow speeds), then can get up to impressive frequencies as they break into a run (e.g., DF<0.5 means there is an airborne phase in a trot). Some of them breached 3 strides/second, which is swinging those stubby limbs awfully quickly!
2. von Baczko, M.B., Zariwala, J., Ballentine, S.E., Desojo, J.B., Hutchinson, J.R. 2024. Biomechanical modelling of musculoskeletal function related to the terrestrial locomotion of Riojasuchus tenuisceps (Archosauria: Ornithosuchidae). Anatomical Record 25528. https://doi.org/10.1002/ar.25528
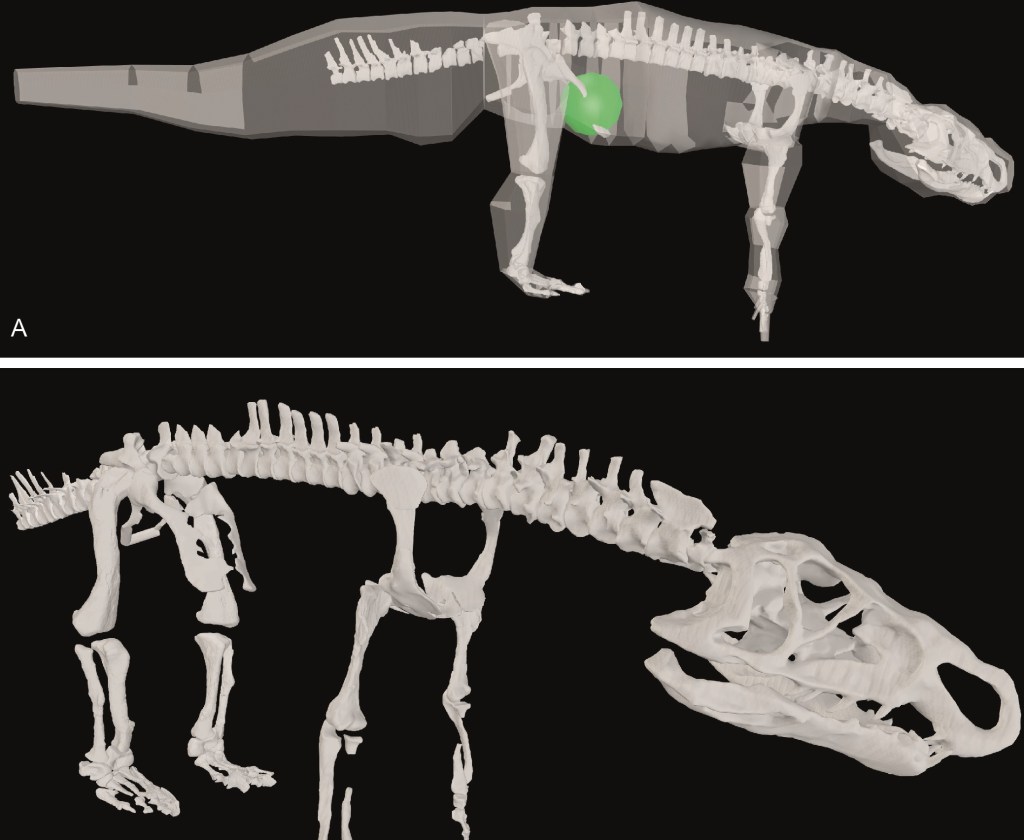
A new DAWNDINOS grant paper; #41 published so far from that project (since 2017), with many many more to come. I enjoyed writing this one, quite a bit. The subject is a member of a small clade of very strange crocodile-line archosaurs (ornithosuchids) from the Triassic period. They have a long history (>100 years) of study and ideas about them have roamed widely, such as what they were more closely related to (theropod dinosaurs? No. Early dinosaurs and pterosaurs? No.). And how they moved: bipedally and/or quadrupedally? Sprawling to erect limb posture? Plantigrade (flat-footed) or digitigrade (tip-toed)? We did our standard approach of scanning the skeletal remains of Riojasuchus tenuisceps, making a 3D musculoskeletal model, and then using that to address lingering questions like those I’ve mentioned. We found the evidence for/against bipedalism to be wonderfully ambiguous, which might sound strange to say, but I like that Riojasuchus tenuisceps remains a conundrum. It is weird. But we were convinced that it used quite erect and plantigrade postures. There’s a lot more to the paper such as consideration of the evolution of hindlimb musculature but that gets really technical. We’ll be doing more with this model, involving deeper biomechanical analysis, in the near future. Here, again, an RVC undergrad, Sarah Ballentine, got involved in the early stages of modelling. A little more about the paper on Twitter.
The model, in the (digital) flesh above, with centre of mass as a green sphere, and just the skeleton below. It’s in our “reference pose”, which is a non-biological straight-limbed orientation having all joint angles at 0 degrees.
3. Etienne, C., Houssaye, A., Fagan, M., Hutchinson, J.R. 2024. Estimation of the forces exerted on the limb long bones of a White Rhinoceros (Ceratotherium simum) using musculoskeletal modelling and simulation. Journal of Anatomy, published online. https://doi.org/10.1111/joa.14041
Much as I’ve long wanted to study hippos as one of the “big four” clades of giant land mammals, I’ve wanted to do so with rhinos; and this is my fifth such study. Here, we (led by French PhD student Cyril) use the same kind of 3D musculoskeletal modelling process as paper #2 above, but including real data from dissections of muscles from a white rhino, and usage of those data in a simple static simulation to estimate how the muscles might be activated to support a quiet standing pose, and what the corresponding muscle and joint forces might be. Surely because it supports more weight, the forelimb had greater muscle activations and forces; especially from some lower limb muscles such as the ulnaris lateralis (an elbow extensor) and digital flexors; and the humerus experienced the greatest forces, reflecting its robust morphology. This is a step toward other kinds of modelling such as finite element analysis of bone stresses. Cyril put in a huge effort to build a nice model and conduct careful analyses with complex input and output data, and he cleverly designed some nice figures of those data.
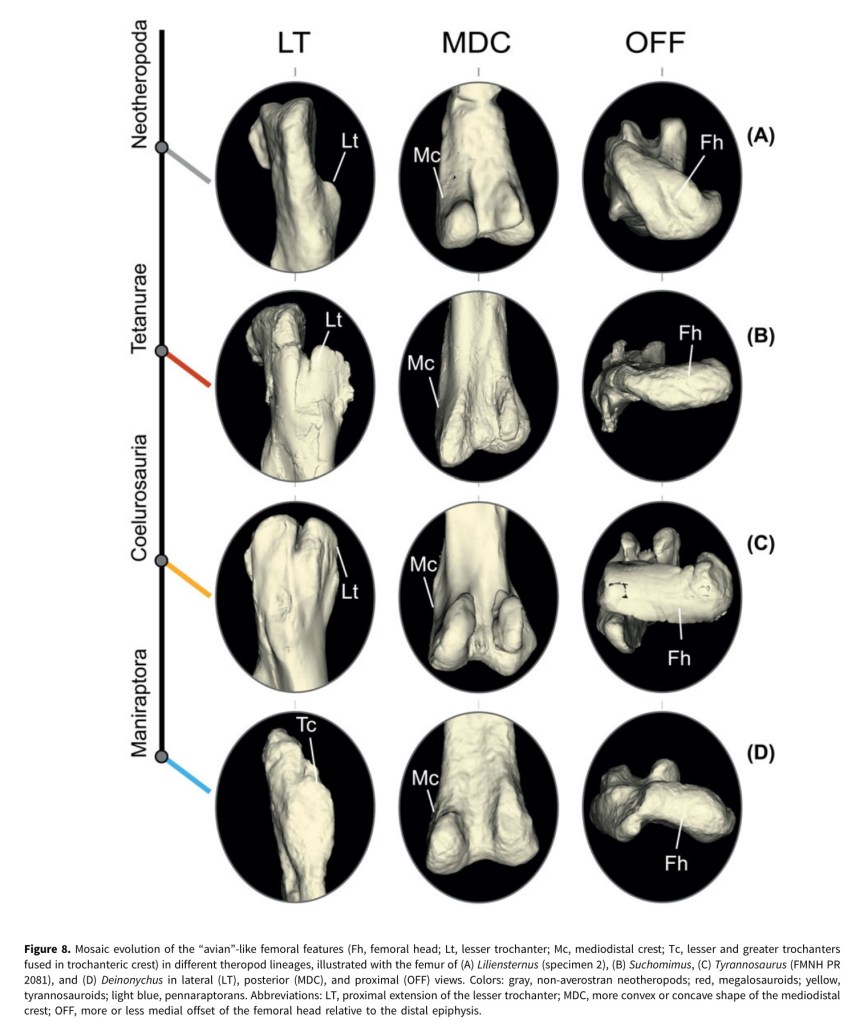
4. Pintore, R., Hutchinson, J.R., Bishop, P.J., Tsai, H.P., Houssaye, A. 2024. The evolution of femoral morphology in giant non-avian theropod dinosaurs. Paleobiology, 1-22. https://doi.org/10.1017/pab.2024.6
A dinosaur paper, imagine that! This one came from 1st author Romain’s PhD and some data we collected during the DAWNDINOS grant; and quite a bit of data that he wrangled during the COVID pandemic lockdowns. The data are 3D scans of 68 specimens from 41 species of Mesozoic theropods, then subjected to 3D geometric morphometrics shape analysis, to see what features related to giant body size or to other things (e.g., more “birdlike” traits). Each time theropods reached giant (>1000 kg) body sizes, their femora (thigh bones) attained many similar aspects of shape, but also more of those “birdlike” traits kept accumulating (and convergently evolving) along the theropod family tree, and even in some giant forms, and especially in “miniaturised” theropods. I like how this paper ties together two of my big research themes, how giant size influences form and function, and how archosaurian/dinosaurian locomotion evolved; and in the context of rich 3D anatomy. And Romain did some stellar work assembling a huge dataset with very rigorous analyses — and gorgeous figures in the paper!
5. Kurz, M.J., Hutchinson, J.R. 2023. Visual feedback influences the consistency of the locomotor pattern in Asian elephants (Elephas maximus). Biology Letters 20230260. https://dx.doi.org/10.1098/rsbl.2023.0260
Hippos, then rhinos, and now elephants! How grand. This paper feels like it was ages ago, and the data collection was — we did the experiments involved back in 2006! Co-author Max Kurz and I wondered, how much would the slow walking patterns of elephants be disrupted if we removed their capacity to see? This question ties into how elephants maintain “stability” (a fraught term in science). Stability is clearly of great importance to giant land animals following “the bigger you are, the harder you fall” principle. Compared to walking with vision, blindfolded elephants were able to keep the duration of each stride close to the average (in one way remaining “stable”) but the noisiness (“consistency”) of their stride decayed across multiple strides, so they tended to take slower or faster strides as they walked across their paddock. This decay would increase the chances of a stumble or fall as blind elephants moved over longer periods. Thus it shows how vision removes that risk, aiding these giant animals in maintaining “stability”. It’s a neat little experiment. We had the paper all written back in 2010 but held back, then in 2023 I decided to go for it and submit what we had; and I am glad we did that. A little more about the paper on Twitter.
A friendly elephant leading an experimental subject, bearing blindfold, belly-mounted GPS unit (to measure speed) and toe-mounted accelerometers (to measure foot on/off timings for determining stride durations).
6. Lacerda, M.B.S., Bittencourt, J.S., Hutchinson, J.R. 2023. Macroevolutionary patterns in the pelvis, stylopodium and zeugopodium of megalosauroid theropod dinosaurs and their importance for locomotor function. Royal Society Open Science 10230481230481 http://doi.org/10.1098/rsos.230481
and
7. Lacerda, M.B.S., Bittencourt, J.S., Hutchinson, J.R. 2023. Reconstruction of the pelvic girdle and hindlimb musculature of the early tetanurans Piatnitzkysauridae (Theropoda, Megalosauroidea). Journal of Anatomy 244:557-593. https://doi.org/10.1111/joa.13983
Brazilian PhD student Mauro Lacerda joined my lab for a sustained period back in 2022, to collaborate on learning how to reconstruct the hindlimb musculature of theropod dinosaurs and use such data in 3D biomechanical modelling. It has been a really fun and productive collaboration, with at least two more papers yet to come. Mauro had been focussing on the “intermediate” theropods (not ‘early’ ones like coelophysoids and ceratosaurs; not ‘late’ ones like carnosaurs and coelurosaurs, but in a hazy in-between phylogenetic position), called Megalosauroidea. As the name indicates, this clade includes the oldest named dinosaur, Megalosaurus (happy 200th birthday!). Many members are Jurassic forms from around the world, often of medium-large size and rather robust build; but also the celebrity dinosaurs, Spinosauridae, are embedded within Megalosauroidea. We set out to first characterise how key skeletal traits of the hindlimbs of these animals evolved, building a nice pictorial and phylogenetic “atlas” of sundry bumps and squiggles on the bones, and mapping how the distribution of those traits compares across groups of early theropods in a “morphospace”. It’s detailed work that should be a helpful reference for all of those working on this interesting clade of theropods and their relatives.
The second study built on the first, by applying methods and evidence I’ve formalised for reconstructing the (hind)limb muscles of theropods and other archosaurs/tetrapods. We estimated where the muscles of three megalosauroids (Condorraptor, Marshosaurus, and Piatnitzkysaurus in order of improved fossil preservation) originated from and inserted onto, where possible. Some bones or regions of bone were unpreserved, limiting what we could estimate, but this is an aspect of the study that I like, as it is a comparative exercise in how much taphonomy influences what we can infer about soft tissues and their evolution across extinct lineages. And it makes possible the usual kinds of 3D biomechanical analyses that my group does — stay tuned for that.
So there you have it. Hippos, early archosaurs, rhinos, earlier and later theropod dinosaurs, and elephants. Lots of locomotor anatomy, biomechanics and evolution. This pretty much sums up my research career and current foci. The variety is part of what makes my job so fun, but the central issues keep it chugging along to push the boundaries of knowledge forwards in my little corner of science. Moreover, it’s a lot of collaboration, around the world and with researchers at different career stages and in varied disciplines from biology to palaeontology to engineering. I love that variety, too. I’m particularly pleased in this set of papers that I’ve included some undergrads, which I’ve been trying to get better at, but which takes some careful planning of studies and persistence (and time!) to get the papers out the door after the students move on.
Stomach-Churning Rating: 0/10; no anatomy, or even images, here.
I got the initial e-mail message (then a formal letter) from the Council back on 22 March 2023, and when I opened it I screamed repeatedly in joy and surprise, for about an hour. I barely could breathe. I could not believe it, even though this process had been going on for so long. I’d trained myself not to expect it, as I knew it was incredibly selective. It was a spectacular feeling that I’ll treasure forever. And it came at a very dark time, when I was full of doubts about myself and my future, with old feelings of ‘imposter syndrome’ resurfacing against a backdrop of terrible health problems plaguing me more intensely for about 2 years. It has tremendously uplifted my mood. It has been weird to hold back any sharing of the news for almost 2 months.
First and foremost, the most important thing is that I thank my family. Most immediately, I would never have had this privileged position without the constant, loving support of my wife before and during raising our daughter (who gave me new delights of fatherhood). We’ve shared some great times. I learned a lot about life and myself. I wasn’t alone; I had someone wonderful to trust and confide in. I can’t put into words how much this helped me, day after day through over 25 years now. And I can’t mention this without mentioning the terrible burden we experienced when my epilepsy struck in 2014, and since then. There were depths of despair that my wife helped me through. I’ve never spoken or written about this particular thing before, but around 2016, when I was having a terrible time getting the right drug to stave off my clonic-tonic “grand mal” seizures, one drug gave me side effects amplifying thoughts and urges to commit suicide to escape the suffering. It was an indescribably scary feeling, and fortunately we switched drugs and now seem to have found a combination that at least temporarily is working for me. That’s but one example. I share it also because suicide is a huge issue that impacts so many others, and I hope my example might be a light of hope, however small it might be. More generally, I have needed a lot of help from my wife, and I got it. Thank you, so much.
My parents gave me a middle-class upbringing that was quick to praise intellectual ability and academic success; my sisters and I all flourished in different ways. I benefitted from excellent resources and experiences with the natural world, from reading library books to visiting museums and exploring the great outdoors near home and during travels; often to my grandparents’ homes in Ohio and Florida, with different nature to investigate. I received a solid education and went to a good university where my intellectual life flowered. I didn’t always excel, and sometimes truly disappointed (cough that ‘D’ in one semester of chemistry, cough), but that was on me. I had fun, too. Too much fun.
Second, I had influential academic mentors. In undergraduate education and research at the University of Wisconsin, from Wendell Burkholder in agricultural entomology, to my own dad’s lab (as a Professor in microbial genetics, to Dianna Padilla’s lab in molluscan marine ecology, to Dana Geary’s and Klaus Westphal’s expertise in (again, mainly invertebrate) palaeontology; and all of their teams. Grad school at the University of California in Berkeley (Integrative Biology department) was marvelous. It transformed the life of my mind and my career. My supervisory committee Kevin Padian, Rodger Kram, Bob Full and Tony Keaveny (and other faculty such as Marvalee Wake, David Lindberg and Bill Clemens) gave me so, so much support and constructive criticism. I struggled for 3 of my 6 PhD years and they got me to my feet. Finally then, I realised what my career trajectory and goals were; I wanted to weave together evolutionary biology/palaeontology (with a morphological foundation) and biomechanics more strongly into the emerging discipline of evolutionary biomechanics. That epiphany has guided my whole career and identity as a scientist. It was a challenging journey but my mentors got me through it, and I had great external role models/mentors; Steve Gatesy at Brown University most prominently amongst them. Next, I was massively fortunate to get an NSF Bioinformatics postdoc to learn more biomechanics and cutting-edge biomedical engineering methods (musculoskeletal modelling and simulation) with Scott Delp at Stanford. This opened up the academic path that I’d dreamed of, giving me the vital technical foundation that I still use today. All of the teams of these people, and fellow graduate students/postdocs, were a fantastic peer group, even more role models, and educators for me.
Third, I’ve had an enormous panoply of collaborators in my own team and externally. I’ve had the opportunity to mentor and work with about 24 postdocs/fellows, 12 technicians and an administrator, 14 Masters students, 12 main and 26 co-supervised PhD students, and lots of undergraduate research projects. I tried to do good for them, too, but I had my share of failures, mistakes and mis-judgements. There has never been a one-way flow of benefits from me to them. I have always prospered from these experiences, crucially by learning new things (e.g., skills; knowledge; understanding of interpersonal relationships) from this interdisciplinary group. I’ve not just benefitted from publishing papers with them, although I love doing that. The same goes for the countless collaborators I’ve had outside of my team. I don’t know how to begin acknowledging all of these wonderful people, but my publications and grants are concrete evidence of their help.
Finally, the RVC has had my back for almost 20 years now. They believed in me enough to give me a job straight out of my first, 2-year postdoc. The aid I got goes from our amazing Structure & Motion Lab (led by Alan Wilson, FRS), which welcomed me into the ‘ground floor’ to help begin building it in 2003 and raising it to global prominence in our fields, to other faculty that became friends and inspirations, to upper management that created a near-perfect environment for me. I always was told to just do good work, in whatever form. They set me free to decide what was best and to just do it, and they gave me (and the rest of us in the SMLab) tremendous, world-class resources (space, funding, infrastructure and more) enabling it. I’d never have had the success I had in getting grant funding without all of this. And I know this kind of situation is a rare privilege. I can’t say I suffered from any biases or other major obstacles that held me back. It was all up to me in the end, to take advantage of what was available. “Onward and upward” was the saying I heard, and what happened.
There’s much more to thank, such as the friends and colleagues, the helpers I never may have fully been aware of, the medical care I got since I first became chronically ill in the mid-1990s (and via NHS since then), and then there’s money, and other concrete support that came from family. I lost my parents in early 2011. They passed on an inheritance that has helped us to prevent lean times. I’m not a financial whiz, and without this money the stress would have been so much harder to bear. Man, do I ever wish my parents were here today. My sisters took care of my parents during their illnesses, and I was in the UK unable to really help. That made a big difference.
But I can’t end without thanking who first nominated me for this Fellowship. I feel very emotional about this. It dredges up profound feelings and memories of good and bad times. Jenny Clack, FRS, came to the RVC to give a college seminar sometime around 2015(?), during our second NERC grant on early tetrapod locomotion. That was another collaboration that gave my career a big boost, and I’ve had a lot of fun with it. I drove Jenny back home after the seminar, and she was in contemplation at one point, carefully asking me something like “What do you think your effect on the field has been?” I came up with an OK answer; I think she might have been satisfied. Not long thereafter, she nominated me for the Fellowship. I didn’t understand the process, but gradually learned and got some guidance through the years. But Jenny died from cancer in 2020, just as the Covid-19 pandemic was about to truly kick off. This occurred during a year that was awful beyond my capacity to express, in innumerable ways; breaking my heart, my mind and my body; from which I’ve not recovered. Losing her deeply wounded me. She was a great friend and mentor. I have enormous respect for her, and I miss her. I wouldn’t have received this award now without her faith in me. Mike Benton kept the ball rolling with the nomination process after she passed, and I thank him too. Whoever else; Fellows or external reviewers; that gave me the thumbs-up during this process, wow, thank you too.
No one pulls themselves up by their bootstraps or is self-made. That’s a stupid myth that serves the privileged.
Now it’s all up to me. I see this moment in my career as a challenge to me. What do I do with this new privilege? Yes, it’s a highlight of my CV and all that, and I can bask in this honour for a while. I will enjoy that. It will be really fun to see what it is like being a Fellow and rubbing elbows with the others in this august academy. Quite a few Fellows have been disabled people, which is a meaningful fact to me, and for which I wrote about in a Fellow nomination section “As a disabled person, Hutchinson has made himself visible to the community as a potential role model and inspiration (see CV), and a major new goal of his career is to maximize the societal benefits from this opportunity.” I’ve thrown down the gauntlet to myself on that. This blog has been a place I’ve done that since 2012; it taught me to do so. Before then, my health troubles had been a more private issue. I’m still thankful that I am, as I’ve mused here before, “not dead yet“. If anyone is seeking to talk with a disabled scientist, either on a smaller scale or publicly, please reach out to me.
What comes next will be exciting new pages for my career and life, and I look forward to that. Blogs haven’t been the same since 2016, but I’ll keep using this one to document my journey, as I like it. Thank you again to the people above, and the many deserving folks that I have not explicitly thanked. This blog has given me the chance to share the joy and pain of life as a scientist, and I am pleased to share this one. I’ll keep sharing.
I’ve been in Paris twice recently, for PhD defenses of co-supervised students, and had a great time. Which included doing one of my favourite things, wandering that amazing city and exploring its streets — and museums! I’ve raved about the perfection of la Galerie de Paléontologie et d’Anatomie comparée before, and that love will never die. But this time, in one of my wanderings I stumbled across a museum that surprised me with how fun it is. (I also got to explore the sewers, which I tweeted about in a thread here.) This is post what it’s about. Désolé, I cannot write this in French.
Stomach-Churning Rating: 3/10; I don’t know what to say, it’s bizarre stuff. But not gory, per se.
This is Le Musée de la Chasse et de la Nature à Paris. The name sounds innocent enough, except for the hunting (chasse) poor beasties part. Tripadvisor said it was a good overlooked destination, so I thought OK, it probably just has some stuffy Victorian-style rooms with dusty old hunting trophies. It had plenty of that, but so, so much more! This museum trip was TRIPPY.
First, there was a special gallery of ceramics, which caught me off guard as I expected boring, fussy relics that my grandparents might have kept in a cabinet. Oh no grandpa and grandma, these would not be for you, not at all. The stuff of nightmares!
As usual, click to embiggen images– and I recommend this, to fully savour their delights!
You could play a scavenger hunt throughout the rest of the galleries, spotting more of these tucked amongst the other displays:
Then at the back of that special gallery there was a fresco-like display, it seemed made out of paper (or was it ceramics like paper?) and it took my breath away. Tons of detail and ghoulish creativity.
Next, lots of just surreal, wacky, disturbing, WTF-why-is-this-here stuff, almost randomly distributed amongst more staid (or sometimes rather naughty/disturbing) artworks and taxidermy of hunting glories as I walked through the labyrinth of galleries:
Like, why a boar festooned with male pheasants?
It’s a hunting museum, and when it went in for hunting imagery and artifacts, it went in with all guns blazing:
And it wasn’t shy about the human figure, either:
Some highlights for me: (1) canned critters!
(2) Kissing apes!
(3) bear… shoes…
How about a 3D view of a weird sculpture of snaky tentacles and… somebody.
As a closer, let’s laugh our way out of the museum. Turn your volume up and have a nice, friendly, entirely not creepy chat with the boar’s head. Remember a certain scene in “Evil Dead 2”? Well I do! One of the greatest scenes in cinema ever, IMO. The boar would fit right in.
Until we meet again, go to the museum if you’re in Paris, and keep up the good fight.
2021-2022: “over and over again & again” sums it up. I do love this band I “discovered” in 2021 though. Finding new music has been a joy for me through these tough recent years.
The pandemic goes on; my life goes on; but it has been another rough 1+ years. I have hardly done any hands-on anatomy as I’m hardly on campus at all, and my team’s work has mostly shifted into digital modelling for now (more about that below; it is not a bad thing though). My main news for 2021-2022 falls into the categories of Life Stuff, and Work Stuff to summarise here. “WTF” sums it up as it has been a… strange time; very challenging at a personal level, due to the Life Stuff.
Stomach-Churning Rating: stuff is weird, but nothing truly stomach-churning is here
Life Stuff: It’s been about the same as 2020-2021; summarised here. Thankfully, no major grief from losing people/pets close to me, this time. But my heath has been really awful instead — my epilepsy returned in May 2021, much to our surprise, after 2 years of remission. I suspect dehydration was a cause, as I later found out that I’d been chronically dehydrated, which came as a shock. I’ve since learned to step up my hydration routine, and I feel better. Right now I’m >5 months seizure-free, after a very hard time of monthly seizures for ~5 months in a row, including a scary one after a flight from Phoenix-London, in which I woke up in a toilet stall at Heathrow baggage claim, very disoriented and alone, eerily with no one in that large men’s loo area. My taxi driver was wondering why I was so late… I am glad I didn’t fall and hurt myself with no one around. Fingers crossed that doesn’t happen again. But on top of that, I’ve been fighting a longstanding chronic illness (details are not necessary) at the same time, and that got very bad in April 2022, sending me to hospital with severe internal infection; very life-threatening, painful and frightening. Again, right now I feel that I’m in recovery, and grateful for some good (overall) care from the NHS. Owing to these health problems though, plus the pandemic and financial challenges, I’ve not been travelling and don’t foresee much of that for my near future. Which also means not enough real holiday; “staycations” in my house just aren’t enough, as I’ve been here for ~2.5 years. I’m starting to do more fun things, finally, again, and that led to this blog post (first one in almost one year). I feel I have some energy to do things that I enjoy again.
Walking tinamou bird XROMM animation
Work Stuff: Mostly that has been pretty good, with a caveat. The DAWNDINOS project still dominated my work life, much to my pleasure. Indeed, just this week I tied the final ribbon on that, formally, with submission of my final grant report to the EU/ERC. The grant ended on March 31, 2022 and I was VERY, VERY sad to have to bid farewell to my team, who I hugely enjoyed working with for those 5.5 years. Now comes the caveat to “work is good”, which is that suddenly I have no funding (feast-to-famine) and “just” one PhD student (Vittorio LaBarbera; reinforcement learning simulations of locomotion); MRes student Georgia Wells just finished; and a Research Fellow (Dr. Masaya Iijima). It looks like I’ll be doing more undergrad research projects than postgrad for awhile, but we’ll see. The grant funding lottery can be hard to predict. Regardless, there’s a lot of fun science going on! With DAWNDINOS, since last summary we’ve cranked out a bunch of cool papers on archosaur locomotor biomechanics — find them here. #25-31 are the newer ones I haven’t blogged about anywhere yet; #25, 28 and 30 are blogged about by Dr. Ashleigh Wiseman here; and #30 (which is, in part, a summary of DAWNDINOS to date) got SICB conference coverage here.
Muscle-bound Euparkeria hindlimbs from our DAWNDINOS paper #28; picture by Oliver Demuth.
DAWNDINOS paper #26 with DAWNDINOS postdoc Dr. Delyle Polet was a serendipitous one inspired by him giving a seminar to our lab when he first came to the UK, and it struck me that his method for using biomechanical simulations with the “Murphy number” (related to pitch moment of inertia; MOI) to test how animals move would work really well with a long-bodied, hefty Triassic pseudosuchian (= large pitch MOI) such as Batrachotomus, whose results we could compare with known fossil trackways of similar archosaurs (e.g., Isocheirotherium). We found evidence for it using at least two running gaits, which was pretty surprising.
And just this week we published another “spin-off” paper (also see van Beesel primate shoulder-modelling studies #17,#29) adapting our 3D digital modelling methods to another taxon. This one came out of left field for me (I’d never expected I’d work on sharks!) but actually fits very well with my research interests in giant animals, biomechanics and palaeontology. We reconstructed the giant shark Otodus megalodon from the best fossils available (including a Belgian vertebral column somewhat neglected since the 1860s), finding that it was ~16m long and >60,000 kg; but this is not the largest it could get, as a vertebra ~50% larger is known! This paper got a LOT of nice press attention, and the video below is perhaps the best science communication release I’ve been involved with (all kudos go to Catalina Pimiento and the animation team she commissioned). Very importantly, the key data are free to use.
Explanatory video by @cookedillustra, Ian Cooke-Tapia
LATE ADDITIONS: But it wasn’t all #DAWNDINOS-related research! I was very pleased to have Dr. Chris Basu’s PhD work with me on giraffid locomotor biomechanics published in PNAS. We showed, with experiments and computer simulations, that Giraffa has unusually low overall leverage (“effective mechanical advantage”; EMA) for its forelimbs during walking (and presumably all gaits/speeds); and even its cousin Okapia does, to a degree; and the extinct giant giraffid Sivatherium too. This is because of its long limbs, which one might look at and call it “cursorial adaptation” but our analysis reveals the tradeoffs of that; as limb length goes up, EMA goes down, and that negatively impacts athletic abilities. All the more reason to be wary of simplistic length-speed conclusions from extinct animals (calling T. rex!). This, with the similar paper on elephant EMA we published in 2010, is one of my papers I’m proudest of; even though neither (curiously) got much (if any) media/other attention. So it goes.
Above: OpenSim simulation of left forelimb of Giraffa during walking; in ~real-time, representing one ground-contact (stance) phase. Green arrow = ground-reaction force (GRF); red lines = major muscle lines of actions (the simulation activated/deactivated them, producing forces to counter the GRF). EMA is the ratio of the muscles’ leverage vs. GRF leverage around joints; it is ~0.3 in a giraffe vs. ~1.0 in a horse. EMA tends to be larger in larger mammals, up to horse-sized, then it gets weird in really big animals.
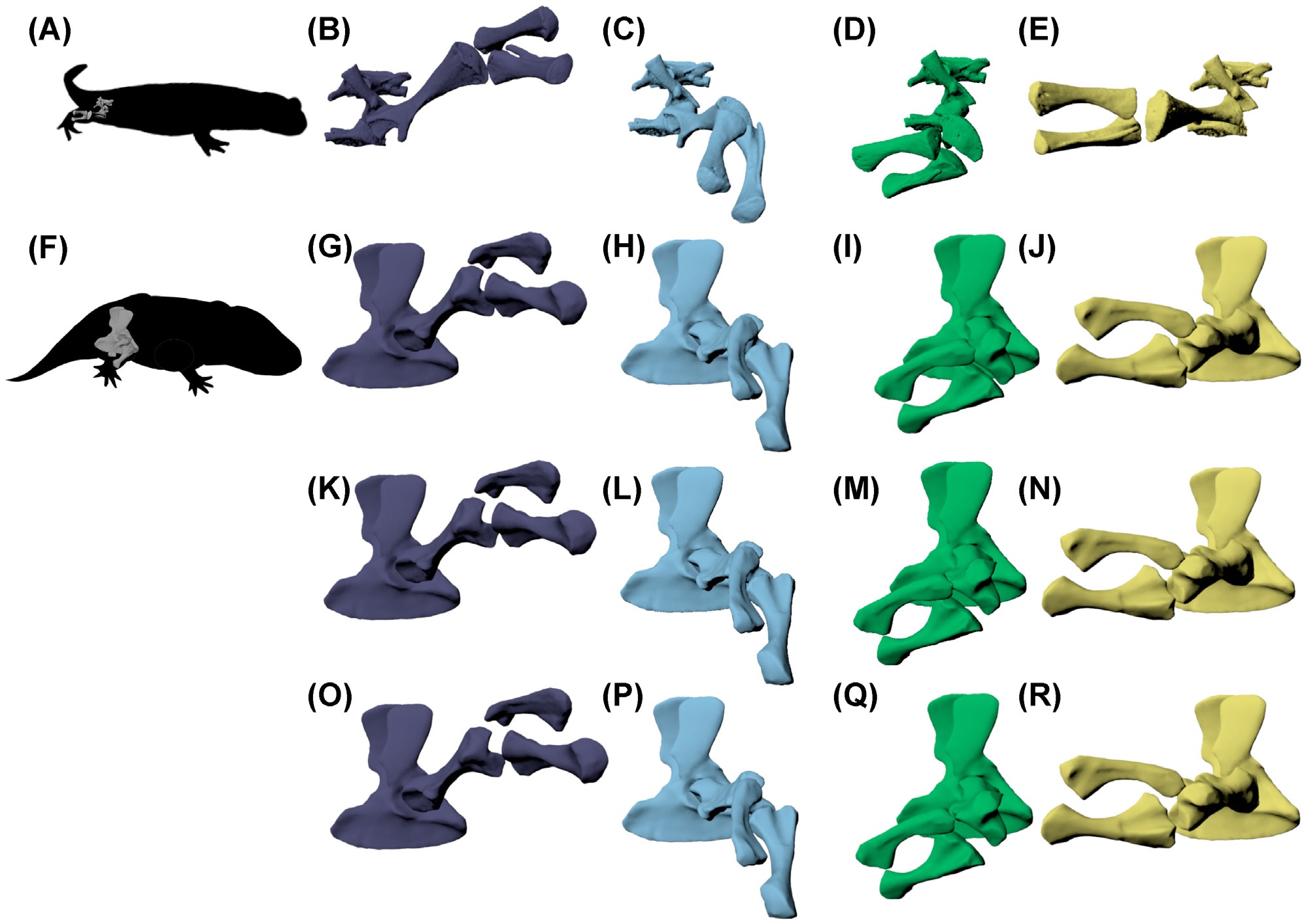
We also scienced the hell out of salamanders. Four papers, all involving Fire salamanders Salamandra salamandra! Three stemmed from my past PhD student Eva Herbst’s work: one explaining a new method to measure joint mobility; another applying that to walking salamandersin vivo and ex vivo; and the third comparing similar data to the Permian ‘giant’ salamander-relative Eryops, showing that its hip and knee joints were about as mobile as a Fire salamander’s. The fourth paper used video analyses of Fire salamanders in a theoretical model and simulation (with other animals) to demonstrate how multi-legged locomotion is controlled. It’s great to have these studies (partly from my old NERC grant on tetrapod locomotor evolution) out after ?5+ years; now Fire salamanders are among the salamanders whose locomotion we understand best. And we have more data still…
Above: Hindlimb configurations in S. salamandra (A) from rotoscoping of in vivo walking, during (B) mid-swing, (C) toe-on,(D) mid-stance, and (E) just before toe-off. These limb configurations were recreated in E. megacephalus (F) with three different knee spacing options: (G–J) tight knee spacing; (K–N) intermediate knee spacing; and (O–R) larger knee spacing, based on the amount of knee spacing present in the rotoscoped salamander at the null pose. S. salamandra configurations in (B–E) were scaled to E. megacephalus knee B.
Oh and I did some science consulting! “Prehistoric Planet” rocked the casbah; glad to see it out ~3 years after I began offering some critiques on the animations. I hope one scene I commented on eventually sees the light of day, as it wasn’t in the final programme. Similarly, “Dinosaurs: The Final Day” did well, and I gave the same kind of input. My experiences with these shows have inspired me to blog someday about how to become, and do, science consulting for documentaries, so watch for that. I may work in some commentary on what it means to be an invisible minority in that context, as I have thoughts.
Blink and you’ll miss me waving my arms about how Carnotaurus might have waved its arms!
There’s a lot of fun science to come, and that keeps me going. We’ve finished initial biomechanical models of 13 extinct archosaurs for DAWNDINOS, and those will become papers on modelling and simulating locomotor function, ultimately testing how performance differed between Pseudosuchia and Dinosauriformes/Dinosauria; and how locomotion evolved (e.g., bipedalism). Some examples in progress are below; these don’t show the muscles or external dimensions reconstructed. Stay tuned in 2022 and beyond for all that! Beyond this, time will tell what I’ll be doing, but DAWNDINOS is going to keep me very busy for plenty of years, and that is good fun for me.
Top image: top to bottom = Postosuchus (pseudosuchian), Heterodontosaurus (ornithischian dinosaur), Riojasuchus (pseudosuchian), Silesaurus (dinosauriform); Bottom image: Gracilisuchus (pseudosuchian), Lago/Marasuchus (dinosauriform), Coelophysis (theropod dinosaur). These are from ongoing studies with DAWNDINOS team members and collaborators around the world. All use 3D scans of the actual fossil material of one main specimen, wherever possible.
I’ve been reluctant to write this post as it meant confronting dark times that I continue to be immersed in with the rest of the world. When I last wrote an “annual” summary, it was exactly when I was leaving my office for “just a few weeks or months” of remote working away from the COVID-19 pandemic. Cue ironic laughter.
Stomach-Churning Rating: unclassifiable due to COVID-19 insanity
What I should write about is what the experience since March 2020 has been like for me. I can try, briefly. Much I am not willing to delve into publicly. I struggled, and suffered; not so directly but indirectly, and from personal issues unrelated to the pandemic, too. I didn’t deal well with the isolation, the boredom and profound ennui, the mass confusion, the under-exercise and over-snacking, the repetition, the anxiety, the uncertainty, the excessive screen time (Netflix! Zoom!), the retooling of teaching for “blended learning” online, the sometimes overly risk-averse bureaucracy that spiralled out of control, the horrifically ugly selfish underbelly of society laid so bare along with smirking racism and other forms of xenophobia (see: Brexit), the endless bad news, the fury I felt at government officials and other (cov)idiots, the feeling of being trapped in my home office and my body; and trapped in a world with political/social machinery that can feel hopelessly broken. I thrive off the novelty and excitement of travel, and without it I withered. These things wore me down to a numb husk and I’m not much better, but am trying to build my energy back so I can pick some battles to make improvements. On top of that, there were tragedies in my life (deaths of two beloved cats; one fairly swiftly from cancer at a young age, one very slowly from old age and related disease), and other awful personal things. I didn’t lose anyone close to me to COVID-19, but I knew lots of people who got infected. Especially in summer 2020 and winter 2020-21, it felt like the punches just kept coming. There’s no way I can describe it all adequately; neither in person nor on this blog. It seems like a lot of people feel that way, but each in our own private way.
I lost a friend in 2020: Prof. Jenny Clack FRS. I’ve not been able to really come to grips with that, even though there was fair warning. Processing grief is on a slow timetable. We had some great times ~2009-2020 working on fossil tetrapods together. I miss her big smiles and our talks about the latest fossil tetrapod news whenever I visited Cambridge. It is hard for me to write more than that right now; I am so drained by the past >18 months. We’ve been served a buffet of flavours of loss to lament.
There’s hope on the horizon, I think, so I cling to that. And we got two new kittens at home, who are simply a joy, and we’ve been clinging to each other. Many cuddles.
It feels very strange, and privileged, to say that work (even though stuck at home) has been a happier thing for me amidst it all. Even that hasn’t been easy, but it has sometimes been a welcome refuge or distraction. I’ve been able to do some real “hands-on” (computer-based) research for long periods, gained skills, and it felt great to be a “real scientist”; not just a manager of scientists; in that regard. I learned a lot about my team’s DAWNDINOS project… and how I’d do it all so differently if I could begin again. At least I have these lessons going forwards. A 5-year, sole-PI, £2.1M project will teach you much about project management. It was so far beyond anything I’d ever tried before. I’m proud of what we’ve achieved now, such as the paper linked above and what’s coming up as the project sadly draws to a close at the end of March 2022. I feel like we’ll be justified to thumb our noses at a few armchair critics, snide naysayers, and cynics, or at least feel personally satisfied even though we’ll never satisfy everyone with our project’s outcomes vs. what certainly were ambitious goals and risks. And I have plans to move on while keeping one foot firmly planted on the foundation we’ve built…
Other than that, I’ve barely touched a work-related freezer since March 2020 and have done no research-related dissections; only digital form and function for me, I’m afraid. That’s OK though. We are slowly migrating back to a bit of in-person working now, and there are some brief dissections and other hands-on work needed as undergrad/Masters research projects begin. I look forward to that.
I was interviewed for two articles about life in science that are meaningful to me: this one on disability pride month, and this one on “advice to my younger self“. Check them out if you haven’t yet?
And that’s all I can say for >18 months in which time has been such a slow grind, warped by the pandemic and the rest of my world.
We released a publication that, for me, comes full circle with research that started my career off. Back in 1995 when I started my PhD, I thought it would be great to use biomechanical models and simulations to test how extinct dinosaurs like Tyrannosaurus rex might have moved (or not), taking Jurassic Park CGI animations (for which the goal was to look great) into a more scientific realm (for which the goal is to be “correct”, even at the cost of beauty). “It would be great”, or so I thought, haha. I set off on what has become a ~26 year journey where I tried to build the evidence needed to do so, at each step trying to convince my fairly sceptical mind that it was “good enough” science. For my PhD I mainly focused on reconstructing the hindlimb muscles and their evolution, then using very simple “stick figure”, static biomechanical models of various bipeds to test which could support fast running with their leg muscles, culminating in a 2002 Nature paper that made my early career. I since wrote a long series of papers with collaborators to build on that work, studying muscle moment arms, body/segment centre of mass, and finally a standardized “workflow” for making 3D musculoskeletal models. And gradually we worked with many species, mostly living ones, to simulate walking and running and estimate how muscles controlled observed motions and forces from experiments. This taught us how to build better models and simulations. Now, in 2021, our science has made the leap forward I long hoped for, and the key thing for me is that I believe enough of it is “good enough” for me, which long held me back. This is thus my personal perspective. We have a press release that gives the general story for public consumption; here I’ve written for more of a sciencey audience.
Skeleton of the extinct theropod Coelophysis in a running pose, viewed side-on. Image credit: Scott Hocknull, Peter Bishop, Queensland Museum.
Stomach-Churning Rating: 1/10: just digital muscles.
Earlier in 2021, we simulated tinamou birds in two papers (first one here), the second one revealing our first ever fully predictive simulations, of jumping and landing; detailed here and with a nice summary article here. That research was led by DAWNDINOS postdoc Peter Bishop and featuring new collaborators from Belgium, Dr. Antoine Falisse and Prof. Friedl De Groote. Thanks to the latter duo’s expertise, we used what is called direct collocation (optimal control) simulation; which is faster than standard “single-shooting” forward dynamic simulation. The simulations also were fully three-dimensional, although with some admitted simplifications of joints and the foot morphology; much as even most human simulations do. The great thing about predictive simulations is that, unlike tracking/inverse simulations (all of my prior simulation research), it generates new behaviours, not just explaining how experimentally observed behaviours might have been generated by neuromuscular control.
OK, so what’s this new paper really about and why do I care? We first used our tinamou model to predict how it should walk and sprint, via some basic “rules” of optimal control goals. We got good results, we felt. That is the vital phase of what can be called model “validation”, or better termed “model evaluation”; sussing out what’s good/bad about simulation outputs based on inputs. It was good enough overall to proceed with a fossil theropod dinosaur, we felt.
Computer simulation of modern tinamou bird running at maximum speed. Grey tiles = 10 cm.
And so we returned to the smallish Triassic theropod Coelophysis, asking our simulations to find optimal solutions for maximal speed running. We obtained plausible results for both, including compared against Triassic theropod footprints and our prior work using static simulations. Leg muscles acted in ways comparable with how birds use them, for example, and matching some of my prior predictions (from anatomy and simple ideas of mechanics) of how muscle function should have evolved. The hindlimbs were more upright (vertical; and stiff) as we suspect earlier theropods were; unlike the more crouched, compliant hindlimbs of birds.
TENET: Thou shalt not study extinct archosaur locomotion without looking at extant archosaurs, too!
Computer simulation of extinct theropod dinosaur Coelophysis running at maximum speed. Grey tiles = 50 cm.
We observed that the simulations did clever things with the tail, swinging it side-to-side (and up-down) with each step in 3D; and in-phase with each leg: as the leg moved backward, the tail moved toward that leg’s side. With deeper analyses of these simulations, we found that this tail swinging conserved angular momentum and thus mechanical energy; making locomotion effectively cheaper, analogous to how humans swing their arms when moving. This motion emerged just from the physics of motion (i.e., the “multi-body dynamics”); not being intrinsically linked to muscles (e.g. the big caudofemoralis longus) or other soft tissues/neural control constraints (i.e., the biology). That is a cool finding, and because Coelophysis is a fairly representative theropod in many ways (bipedal, cursorial-limb-morphology, big tail, etc.), these motions probably transfer to most other fully bipedal archosaurs with substantial tails. Curiously, these motions seem to be opposite (tail swings left when right leg swings backward) in quadrupeds and facultatively bipedal lizards, although 3D experimental data aren’t abundant for the latter. But then, it seems beavers do what Coelophysis did?
Tail swings this-a-way (by Peter Bishop)
Computer simulation of extinct theropod dinosaur Coelophysis running at maximum speed, shown from behind to exemplify tail lateral flexion (wagging). Grey tiles = 50 cm.
The tail motions, and the lovely movies that our simulations produce, are what the media would likely focus on in telling the tale of this research, but there’s much more to this study. The tinamou simulations raise some interesting questions of why certain details didn’t ideally reflect reality: e.g., the limbs were still a little too vertical, a few muscles didn’t activate at the right times vs. experimental data, the foot motions were awkward, and the forces in running tended to be high. Some of these have obvious causes, but others do not, due to the complexity of the simulations. I’d love to know more about why they happen; wrong outputs from such models can be very interesting themselves.
Computer simulation of modern tinamou bird (brown) and extinct theropod dinosaur Coelophysis (green) running at maximum speed. Grey tiles = 10 cm for tinamou, 50 cm for Coelophysis.
Speaking of wrong, in order to make our Coelophysis walk and run, we had to take two major shortcuts in modelling the leg muscles. The tinamou model had standard “Hill-type” muscles that almost everyone uses, and they’re not perfect models of muscle mechanics but they are a fair start; it also had muscle properties (capacity for force production, length change, etc.) that were based on empirical (dissection, physiology) data. Yet for our fossil, because we don’t know the lengths of the muscle fibres (active contractile parts) vs. tendons (passive stretchy bits), we adopted a simplified “muscle” model that combined both into one set of properties rather than more realistically differentiating them. It was incredibly important, then, that we try this simple muscle model with our tinamou to see how well it performed; and it did OK but still not “perfect”, and that simple muscle model might not work so well in other behaviours. That was the first major shortcut. Second, again because we don’t know the detailed architecture of the leg muscles in Coelophysis, we had to set very simple capacities for muscular force production: all muscles could only produce at most 2.15 body weights of force. This assumption worked OK when we applied it to our simulation of sprinting in the tinamou (vs. average 1.95 body weights/muscle in the real bird), so it was sufficiently justifiable for our purposes. In current work, we’re examining some alternative approaches to these two shortcuts that hopefully will improve outputs while maximising realism and objectivity.
Computer simulation of extinct theropod Coelophysis running at maximum speed, shown alongside running human (at 4 m/s) for scale and context. Image credit: Peter Bishop.
If you pay close attention, our simulations of Coelophysis output rather high leg-forces, and it’s unclear if that’s due to the simple muscle model, the simple foot modelling, or is actually realistic due to the more vertical (hence stiff) hindlimbs; or all of these. Another intriguing technical finding was that shifting the body’s centre of mass forwards slowed down the simulation’s running speed, as one might expect from basic mechanics (greater leg joint torques), but unlike some prior simulations by other teams.
Computer simulation of extinct theropod Coelophysis shown alongside running human for scale and context. Shown from above to illustrate tail wagging behaviour. Image credit: Peter Bishop.
Users of models and simulations are very familiar with catchphrases like “all models are wrong, but some are useful” or the much more cynical (or ignorant) “garbage in, garbage out”; or the very dangerous attitude that “if the mathematics is correct, then the models can’t be that wrong” (but if the biology is wrong, fuggedaboutit!). These are salutary cautionary tales and catechisms that keep us on our toes, because the visual realism that realistic-looking simulations produce can seduce you into thinking that the science is better than it is. It’s not a field that’s well-suited for those fearful of being wrong. I’ll never think these outputs are perfect; that is a crazy notion; but today I feel pretty good. This was a long time coming for me, and it is satisfying to get to this stage where we can push forwards in some new directions such as comparing simulations of different species to address bigger evolutionary questions.
The wrestling with scepticism never ends, but we can make progress while the match goes on.
The blog is back! Briefly. With dinosaurs. Back in 2005, I published a paper in which I used a “SIMM” 3D musculoskeletal biomechanical model of Tyrannosaurus rex to analyse its muscle actions and infer a relatively upright hindlimb pose. This was an outcome from my NSF-funded postdoctoral research at Stanford University, in which engineers kindly taught me how to use SIMM (handing me a loaded gun?). Part of my plan all along was to build multiple such models along a rough evolutionary sequence to revisit old questions I had with past, qualitative functional morphology papers from 2000 onwards, and see if biomechanics could quantitatively reveal more about the functional evolution of dinosaur hindlimb muscles. So I got data for modelling some extinct dinosaurs (theropods Dilophosaurus, Allosaurus, Velociraptor) and living birds (Struthio, others) and published nuggets of that but held others back…
Stomach-Churning Rating: -1/10; dinosaurs!
I handed these 3D model data off to my PhD student Vivian Allen in ~2007, charging him with the task of making more models to flesh out the phylogeny and finish what I’d started. And he sure did. He graduated, did a couple of postdocs with me, and we gradually massaged his thesis chapter on this topic into a draft paper. Easier said than done, though! That’s why 14 more years have passed.
Viv came up with some clever tools in MATLAB software code (from which he became a very competent programmer and went on to a successful career in that!) to boil complex data on muscle leverages (moment arms) across a wide range of joint motion for the hindlimbs for each taxon.
These data then were fed into further code that took the results from all models, ultimately 13 of them from an Australian freshwater crocodile to two living birds and 10 extinct dinosaurs plus close cousin Mara/Lagosuchus (Figure 1). The code expressed these leverages as changes in ancestral values along the main branch of the evolutionary tree from early (Triassic) “ruling reptile” Archosauria (represented here just by the croc as a proxy) to modern birds, and 9 main ancestral “nodes” in between. Our code tracked both how each of 35 hindlimb muscles we modelled evolved in its leverage, as well as overall “average” leverage of functional groups around the hip, knee and ankle joints.
So, back and forth we went for some 10 years playing with the models (see Video below), data and code, and the paper describing the whole thing, slowly closing in on a final version but also sometimes distracted by our other projects and Real Life Stuff like health and children, and concerns about how we conducted this study (i.e. a lot of fiddling).
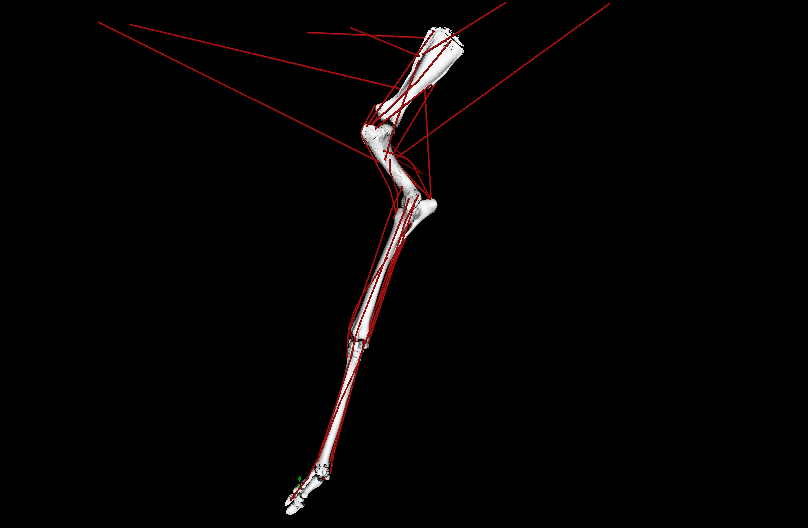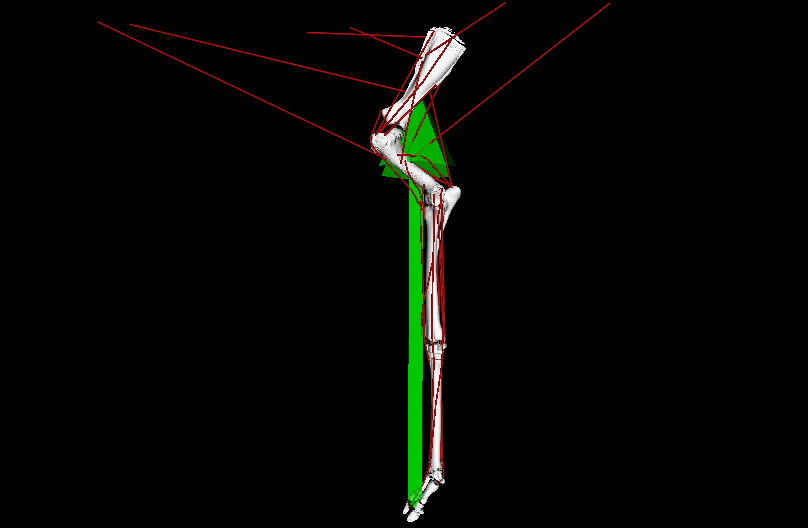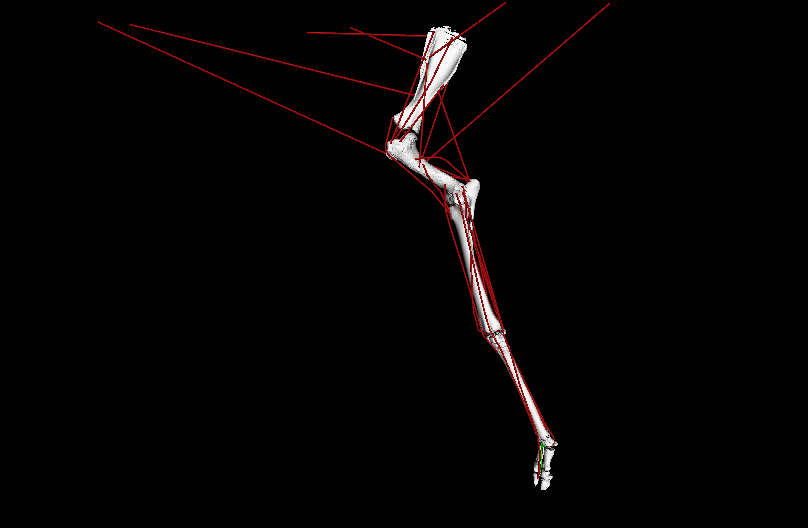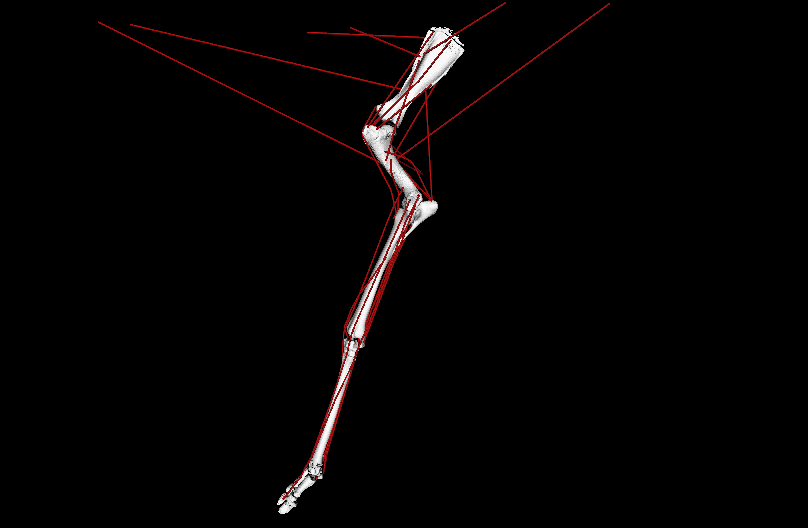
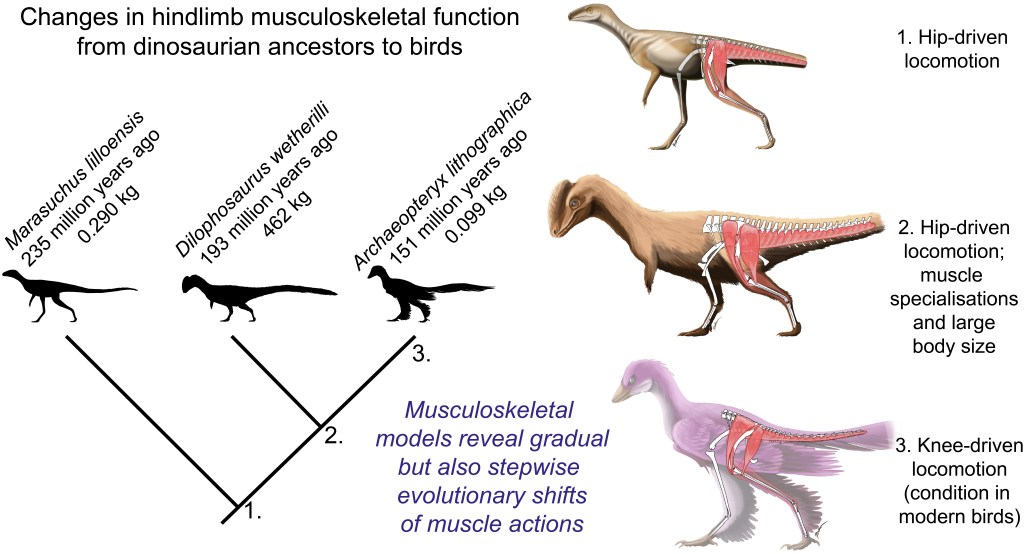
Figure 1: Evolutionary tree of dinosaurs and their relatives as used in the study, showing all 13 models, species names, and names of groups along the bottom (red nodes) of the tree. Averostra and Avetheropoda were ancestral groups of theropod dinosaurs that the study inferred had particular specialisations of the hindlimb muscles. Right hindlimbs in side view. The limbs are all straightened vertically into a baseline reference posture but the study investigated variation in muscle function across a wide range of limb poses.
Then I got a new grant “DAWNDINOS” that changed the scene for me, refocusing my team’s energies onto the Triassic (and early Jurassic) and the evolutionary biomechanics of diverse archosaurs’ locomotion, assessed with both LOTS of experimental studies of living crocs and birds, and LOTS of predictive simulations of locomotion. Stay tuned for much more on that from our team, but we’ve already published some key steps here. Most notably, we developed an improved protocol for modelling and simulating our animals, as shown by Bishop et al.’s 2021 study of the early theropod Coelophysis bauri (also appearing in the current paper). Awkwardly for me, that new method rendered our old models and methods a bit obsolete (although still fine), so I pushed to publish this current paper with Viv, and brought collaborator Dr. Brandon Kilbourne on board to aid in some final stats, figures and more. That finally did it, and now we’ve published the paper in Science Advances. Deep breath.
Video: Rotating movies of 3 musculoskeletal models from this study. Models have been posed into representative limb orientations illustrating a gradual or stepwise transformation from more upright to more crouched.
Well what’s the paper about, then? We used our 13 models and processed evolutionary functional patterns to test three main questions (hypotheses) about muscle leverage, making educated guesses at what might prevail from early Archosauria to Aves:
Hip extensor / flexor (i.e. femur retractor/protractor) moment arm ratios remained constant. We weren’t sure what to expect, as these antagonists both seem to change a lot on the whole lineage, so we went with this prediction.
Knee flexor / extensor ratios decreased; i.e. the flexors (“hamstrings” etc.) weakened and/or extensors (equivalent of our quadriceps) strengthened their leverage. Anatomy of the knee joint and muscles around it suggests this, plus since Gatesy’s 1990-onwards studies we’ve expected archosaurs to shift from more ‘hip-based’ to more ‘knee-based’ locomotion as we get closer to avian ancestry.
Hip medial (internal) long-axis rotator / abductor (i.e. pronators of the limb vs. those that draw the leg away from the body) ratios increased. This idea comes right from my paper w/Gatesy in 2000, where we surmised that archosaurs shifted from relying on hip adductors (in crocs/other quadrupeds) to abductors (in bipedal dinosaurs; like humans) to medial rotators (‘torsional control’ as in birds today) during weight support.
Moreover, we reconstructed the evolution of 35 muscles’ actions across ~250 million years, which was a new step.
Here’s a summary of what we found (Figure 2):
Figure 2: Short visualization/explanation of the study’s main insights. Pictures by palaeoartist Jaime Headden: https://qilong.wordpress.com/about/ in left side view, including “muscled” and silhouette images. Right side images include representative hip, knee and ankle muscles from the study. Changes such as the enlargement of muscles in front of the hip that straighten the knee, and reduction of the caudofemoralis longus muscle that runs from the tail to the back of the thigh, are evident.
So, overall hypothesis 1 about hip extensors/flexors ended up complicated; rejected because hip flexor leverage actually increased. Furthermore, we found that around the ancestral nodes for early theropod dinosaurs (Neotheropda through Avetheropoda; around 200 Mya), there were peaks in muscle leverage (size-normalized) that surprised us, and persisted despite many different analyses we threw at them over the years. As far as we could tell, these peaks that kept appearing for various muscles’ actions were “real” (estimates). Which meant these ancestors may have had specialised high leverage relative to both their own ancestors and descendants; the peaks got reversed in evolution. These ancestors had some other weird anatomical and functional traits, such as tightly articulated hip joints early on (which they lost later), increased body size in the later forms, more ‘macropredatory’ ecology (e.g. eating sauropods?), and a centre of mass of the body that was shifted forwards (due to big arms and heads/necks). This weirdness is a cool unexpected finding that showed up for the other hypotheses too, and it needs some more investigating. A ‘failed’ hypothesis test led to neat insights.
Figure 3. From the paper– showing our main results for changes in moment arm ratios across archosaurian ancestors. Hip extensors/flexors decreased then increased; knee flexors/extensors decreased; and hip medial rotators/abductors decreased then had a series of increases.
Hypotheses 2 and 3 found good support, on the contrary, overall (Figure 3). We seem to have been able to quantify the shifts from hip-based to knee-based, and abductor-based to medial-rotator-based, muscle actions. I find that very satisfying. Ankle weight support (extension) capacity also increased, which fits morphological changes fairly well. If you’re into archosaur limb muscle form and function, there’s a lot more food for thought in the paper.
Funnily enough, ~20 years has been sufficient time that we could have had plenty more models in this study if we’d delayed it even longer and re-re-re-analysed our data. But we had to draw the line somewhere and not infinitely revise with every new model we’ve been creating. With the current state of musculoskeletal modelling in my group, we could have more than doubled our sample size and fleshed out the most important gaps such as in the crocodile-lineage (extinct Pseudosuchia) and other Triassic forms plus elsewhere. A big challenge remains having some nice 3D-preserved early fossil birds beyond Archaeopteryx; e.g. so many nice Chinese ones are too flat (e.g. joints we need) to reliably model here. It’s something that can still be done and is worth doing, but I suspect the general trends we’ve found along the dinosaur lineage are “correct”.
What’s personally important to me about this paper is (1) how it not only bridges a huge morphofunctional gap across archosaur evolution in scientific terms, and (2) how we’ve completed a long-delayed project with stubbornness (and during a pandemic!), but also (3) how it bridges my past career from my PhD and postdoc to the present work with DAWNDINOS. We’re now forging well beyond what this new paper has done in terms of truly testing, as best we can (estimate) so far, how limb muscles of archosaurs functioned and evolved, and how these contributed to particular behaviours and performance (maybe even palaeoecology and evolutionary success/extinction?). The current paper is just simple modelling of muscle leverage, but leverage is only one (very important!) piece of muscle function and performance. With fully dynamic, anatomically integrative, physiologically and physically representative biomechanical computer simulations that predict what living and extinct archosaurs could or could not do, we can do even better. So watch for that! Hopefully it won’t all take 20 years, or 250 million.
Our special guest post this week comes from Dr. Liz Clark of Yale University in New Haven, Connecticut, USA. She continues to bring biomechanics-fu to echinoderms– the weird marine critters like seastars and sea urchins. Including fossils, as you’ll see today! You may remember her from blog posts such as “Guest Post: Brittle Star Arms Are Weird“.
Stomach-Churning Rating: 1/10; echinoderms are inoffensive.
Imagine that you’re stuck in a cardboard box on the beach, holding a small stick. Could you use the stick to move yourself forward? What would you do? You could try digging into the sediment ahead of you to pull yourself along. You could try rowing side to side, as if you were in a rowboat. Or maybe it’s not possible and you’d give up, decide to stay put, and wave your stick in the air for help.
Believe it or not, this is a strange-but-important dilemma that some paleobiologists- like me!- have been wrestling with for generations. My research specialty is in the biomechanics of locomotion– how organisms use their bodies to get from one place to the next (through walking or swimming, for instance). We can learn a lot about an animal by studying their locomotion, such as why their body is shaped the way that it is, or what role they occupy in their ecosystem. Animal motion is a major inspiration for robotic design, and I work with engineers to apply the novel insights on animal locomotion from my research to create new kinds of devices.
Studying the biomechanics of motion in living organisms is (relatively) straightforward. We can use high-speed cameras, motion capture software, and 3D imaging tools to visualize and understand how organisms move in real-time, informing our inferences about how they perform certain tasks. Inferring locomotion in fossil organisms, on the other hand, is tricky since we can’t observe the organism’s behavior like we could if the organism were alive. Instead of being able to watch the organism move, we’re left with a snapshot of the animal frozen in place in a rock. We’re also missing a lot of physical information: locomotion in most animals requires soft tissue and hard skeletal structures, but typically with fossils, only the hard structures get preserved.
However, we can often garner some insights from living organisms to determine the locomotion strategies that fossil organisms use. Most organisms in the fossil record look at least somewhat similar to organisms alive today. If our fossil has four legs, for instance, we can study locomotion in living tetrapods (four-legged animals) to help us create a framework for deriving inferences about locomotion in our extinct tetrapod fossil animal. But for some really strange-looking animals- ones without obvious modern analogues- we’re not so lucky. For me, this is where the fun begins.
Figure 1: Stylophorans! Here are four fossilized stylophorans from the Helderberg Group of the Early Devonian (YPM 036413)
So getting back to the cardboard box and the stick. These are metaphorical examples of the different locomotion strategies that have been proposed for a group of fossil animals known as stylophorans (Figure 1). Stylophorans are extinct organisms related to sea stars and sea urchins, but with a body structure unlike any organism on the planet today. They have a large, relatively flat body called a theca (i.e., the cardboard box), and a long, thin segmented tail known as the aulacophore (i.e. the stick) (Figure 2). They’re known in the paleontological community as some of “the strangest-looking animals of all time.”
Figure 2: Stylophoran anatomy. The “theca” is the body cavity, and the “aulacophore” comprises of the proximal aulacophore, the stylocone, and the distal aulacophore.
By reconstructing stylophoran locomotion, we can unlock the mechanics of a unique system for motion and its potential applications to engineering. We can also understand more about how this organism lived and functioned in its ancient ecosystem. And, by developing a new approach to understand locomotion in stylophorans, we can apply this strategy to analyze locomotion and movement in other unusual fossil animals as well!
For years, scientists have been documenting the incredible array of stylophoran diversity in the fossil record and making their best predictions about how they would have been able to move (or not!). These predictions are based on their morphology– the structure of an organism’s body. For stylophorans, that means the shape and structure of the theca and aulacophore. There are a variety of stylophoran thecal shapes, ranging from ovoid in Enopleura to trapezoidal in Ceratocystis to almost crescent-shaped in Cortnurnocystis. There’s a similarly wide array of aulacophore morphologies as well.
Figure 3: Left: One half of the concretion within which the stylophoran fossil we analyzed is preserved. Right: The 3D digital image of the stylophoran fossil, created by micro-CT scanning the fossil specimen.
We developed a new approach using 3D imaging (Figure 3) to create a digital model of a stylophoran specimen. We used the model to test if several different locomotion strategies that had been proposed before were physically possible or impossible for a stylophoran to actually perform.
First, we used a micro-CT scanner to image a fossil stylophoran. This outputs a digital 3D picture of the stylophoran fossil that we can look at and analyze on a computer. Next, we developed a program to calculate the joint centers- the point at which one skeletal structure rotates relative to another-within the digitized stylophoran’s aulacophore (Figure 4). This created a digital marionette– a rig of our stylophoran fossil that flexes at the junctures between aulacophore segments as it would have in life. We then rotated each segment at the joint center to calculate the aulacophore’s total range of motion– a reconstruction of how far the aulacophore could flex in each direction (Figure 5).
Figure 4: A look into some of the nuts and bolts of the 3D model we created. Tri-colored axes demarcate where the joint centers are in the proximal aulacophore.
We used this 3D range of motion model to evaluate several locomotion strategies that had been previously hypothesized for this group of stylophorans. One hypothesis suggested that these stylophorans dug their aulacophores into the substrate– sediment on the ocean floor- to pull themselves forward. Another suggested that they moved the aulacophore side to side in order to push themselves along. We found that the first hypothesis would have been impossible to conduct based on the range of motion we calculated, but the second strategy was theoretically possible! We’ll need to do more work to see how likely it was that stylophorans would have actually used this technique. Nevertheless, through this investigation, our team produced the first objective, data-driven methodology for analyzing locomotion in fossil invertebrates, which is a big step in the right direction for the study of fossil invertebrate biomechanics! Our technique can be applied to study other organisms with rigid skeletons as well, like crabs, insects, or sea stars, for instance, and we’re looking forward to seeing our technique used to uncover more interesting locomotion strategies!
Figure 5: A snapshot of the 3D model where we can observe how dorsal and ventral range of motion compare to the originally preserved orientation of the aulacophore (highlighted in green).
We live in a weird future. In the Coronavirus pandemic anything seems possible; entropy has been set loose from its cage. Within this higgledy-piggledy universe, I realize that I have forgotten to write annual summaries for this blog for the past 2 years (2018-present). Physical distancing means that I can now make amends — will this pandemic mean the rebirth of the blog, as a means of social un-distancing? If only. Yet in this post, at least, I can take a nostalgic look back at different times. Everything was blissful in 2018 and 2019, right? Oh…
From the Bone Church/Kutna Hora near Prague, Czech Republic. One of those posts I haven’t made. It was an amazeballs sight.
Stomach-Churning Rating: 4/10 bones (+ bird muscles and bone pathology below), and the years 2018+2019 to boot. May 2021 be more sane.
I’ve been distracted. The DAWNDINOS project has taken an increasingly tremendous amount of my time, energy and concentration, and while it has been fun at many times it also has been the hardest thing I’ve ever done as a scientist. I knew it was a HUGE task when I got the funding notice, I cussed profusely, and then we dove in… and it was huger than huge. It will keep me busy for the rest of my career, no joking. But in good ways, too.
What has happened on the blog? (For year 6 go here) In 2 years, about 14 posts have happened, which surprises me (as it always seems to when I write these summaries). I guess this blog is not dead. Good!
“A day in my life” still roughly applies. I have learned to be even more focused, organised and efficient. Because work/life forced me to. I’ve also learned to enjoy that feeling of intense focus. When I am in that groove, I let it rip; when I’m not, I let it slide.
Much as the “day in John’s life” post noted, my career has shifted to encompass new roles, differing balances of duties, and important lessons. I wrote about “learning by serving” to give some views on service and administration as a rewarding side of being a scientist.
The new UMZC in Cambridge became a theme on this blog: my first visit to the renovated exhibits and building were followed by a second, and then a third! Click those links to see the new-new-new UMZC. We’re lucky to have it in England.
Museum blog posts were popular for me (not so much for readers, it seemed?) over the past 2 years, including the venerable and still-awesome MNHN in Paris. I’ve also hit a few Bodyworlds-type exhibits for the blog over the years, and the big London/Piccadilly Circus one became a must-see, so take a peek here. Then I had the joy of seeing a short-lived Ray Harryhausen exhibit in London. Childhood me would have exploded with glee. Very messily, but a happy end nonetheless. And I happened to walk into a palace-museum in Lausanne — and wow! I will try to remember not to skip those opportunities. Not-so-famous museums can still floor you.
Silly chimeric specimens at Museum of the Weird in Austin, Texas. Worth the ticket and time.
Post-publication peer review is great, but what about pre-submission publication planning for scientific papers? I gave some advice from my experience as author, reviewer, editor and mentor.
Chonky superficial muscles around the left thigh of a tinamou, from DAWNDINOS research-to-come-soon.
Finally, quite a few of the past years have had the quiet undertone for me of trying to make a change of location (i.e. moving back to the USA for another science job). Repeated failures were not fun, as for anyone in the academic job rat race. That long felt like a forbidden, lonely topic but I broke the silence as a form of self-therapy and it helped. I really appreciate the kind feedback I got on that post, too. It was a risky, vulnerable move to make but I am happy I did it. It’s one of my favourite posts on this blog.
Peer back into the forbidden past of thrash metal and you’ll find screamy classics like this one.
Hmm. I notice in writing this that there haven’t been as many classic “hey here’s something cool I dissected, brace yourselves for the awesome anatomy!” posts on the blog as there used to be. That activity has shifted more to Twitter or else just not been shared, either because I am just dissecting the same old thing (groan, ANOTHER crocodile, another ostrich, another elephant foot!) or I am just out of time. Which is what I am now. Here, consider this pathological (osteomyelitic) ostrich fourth toe while I tiptoe away. Ouch!