Ho ho ho! The vagaries of the scientific publication system today brings forth TWO open access papers on crocodylian functional anatomy, evolution and biomechanics, from my team with others’; including our DAWNDINOS project in part. Get ready to bite down on the science! I’ve loved crocodylians throughout my life– “dacadile” was among my first words, for a beloved stuffed croc toy, and “Alligators All Around” was an early favourite song (it’s still GREAT).

One of the many large adult alligators in St. Augustine, Florida.
Stomach-Churning Rating: 1/10; bones and movies of awesome behaviours.
First, I am so relieved and pleased to finally publish an experimental study I began over 17 years ago. This is my most-delayed paper ever, due to my own perfectionism, overcommitment and failures at funding it more broadly. But published is published and I’m glad to see it out. We collected a large experimental dataset from 15 species of Crocodylia at the St Augustine Alligator Farm Zoological Park (a conservation/education centre) in Florida. (No matter how you species-ify them, that’s a good chunk of diversity; roughly half or more.) This was a non-invasive study of 42 individuals ranging from 0.5 to 43 kg in body mass (hatchlings to adults). Larger adults were too dangerous or too slow to work with. It took 3 years (2002, 2004, 2005) of data collection to assemble this, with some twists and turns (including a close brush with Hurricane Katrina), and then a lot of analysis and reanalysis; and I’d do it all very differently if I did it today but that’s a moot point. So what’s the paper about?

Adorable Siamese crocodile family “cuddling”. Crocs are great parents! IIRC, that is the father shown.
Some Crocodylia (the inclusive modern name for all crocs, caimans, gharials, gators) are known to use what we call asymmetrical gaits: “mammal-like” footfall patterns in which the left and right limbs do not move as mirror images of each other. In particular, these gaits include galloping (rotary or transverse; either way a “4-beat” pattern with left-right hind- followed by right/left forefoot contacts) and bounding or half-bounding (the former being the most extreme, with left-right hind- and then forefoot contacts as synchronous pairs). Often people just say that crocs can “gallop” but this confuses/conflates the issue and omits that they can use these faster bounding gaits. Regardless, we’ve known about these gaits at least since HB Cott’s 1961 photographic documentation of them in Nile crocodiles; and more detailed studies of Australian freshwater and saltwater crocodiles in the 1970s-2000s. But very often, scientists and popular natural history accounts ascribe the asymmetrical gaits to only a few species or young individuals.

“Freshie” croc bounding in the wilds of Australia; credit Kent Vliet.

Osteolaemus dwarf African crocodile getting marked up for study.
That’s where we came in. We had access to a huge collection of captive Crocodylia and a very supportive institution (with coauthors from there as a result). I wanted to know which Crocodylia do use asymmetrical gaits, having a very strong suspicion from the literature that Alligatoroidea, the alligator and caiman lineage, don’t use them, whereas their cousins the “true crocodiles” in Crocodyloidea do. And I wanted to test how body size interacted with this ability, as prior accounts hinted that asymmetrical gaits got lost with increasing size or in adults. Finally, I was interested in what the benefits of asymmetrical gaits were– did they give those that used them marked boosts in performance, especially maximal speed? Answering that would help understand why these gaits are used.

Cuban crocodile Crocodylus rhombifer in preparation. A gorgeous but aggressive species that we handled carefully.
So we walked and ran our subjects across some platforms past video cameras and collected about 184 useful trials or strides of gait across level ground at a wide range of speeds; and a LOT of not-so-useful data (mostly subjects just sitting and pouting). We found that, yes, most Crocodyloidea we studied could bound or gallop; and no Alligatoroidea did. In the latter case, we didn’t use as large a sample of subjects as we could have, partly because it already seemed evident that alligators did not use asymmetrical gaits, and partly because those alligatoroids we did try to coax to move quickly either only used symmetrical gaits (e.g. trotting) or would only sit and fight or hiss. And we found that bigger animals moved at least relatively more slowly and less athletically, and perhaps even more slowly in absolute terms (metres/second).
Most intriguingly to me, it didn’t matter what gait alligatoroids or crocodyloids used. They all could move at roughly similar top speeds if they wanted to; less than 5 m/s or 11 mph. It’s just that crocodyloids tended to use asymmetrical gaits, especially bounding, at top speeds– but not always: some even chose to trot at their top speeds. We don’t know why, and we still don’t know why asymmetrical gaits are chosen but they likely have other benefits such as acceleration and manoeuvrability.
It’s a thrill to finally be able to share the huge dataset, including a gigantic file of videos (with some highlights shown here), with the paper, closing this study at last. It should be very useful to anyone studying Crocodylia or wanting to educate people about locomotion. I’m a bit tired of hearing that galloping is a mammalian behaviour when we know so well that many species of animals do it, or something like it. And it was absolutely thrilling to see five species of Crocodylia bound or gallop when they hadn’t been properly documented to do it before– enough anecdotes, here’s cold hard facts from video on what happens. What remains is a mystery: did Crocodylia have this ability to use asymmetrical gaits as an ancestral trait, as almost everyone assumes (and thus alligators and caimans have lost or essentially never express the ability), or did crocodiles uniquely evolve this ability more recently? I would join most scientists in wagering on the former; and there are good reasons to suspect the ability goes deeper into extinct Crocodylomorpha.
(my favourite video is below!)
Want more cool videos? Try my Youtube channel— or if you want ALL of the videos, go here!
Next, Torsten Scheyer was kind enough to invite me to join his team in studying a fossil I’ve long been fascinated by: the “giant caiman” Purussaurus mirandai, from the Miocene (~6 million years ago?) of Venezuela, in the Urumaco Formation‘s very weird biota. Purussaurus has been known of for >125 years but Torsten’s team noticed that Purussaurus (mirandai) specimens tended to add one of their trunk vertebrae to their hip girdles (sacrum; normally only two vertebrae in Crocodylia but here three), and that the shoulder and hip girdles had unusual bone morphology (straighter, more vertical relative to the body). So they asked me to help interpret these features. And here’s the paper!

Infographic by Torsten Scheyer’s team– click to emcroccen!
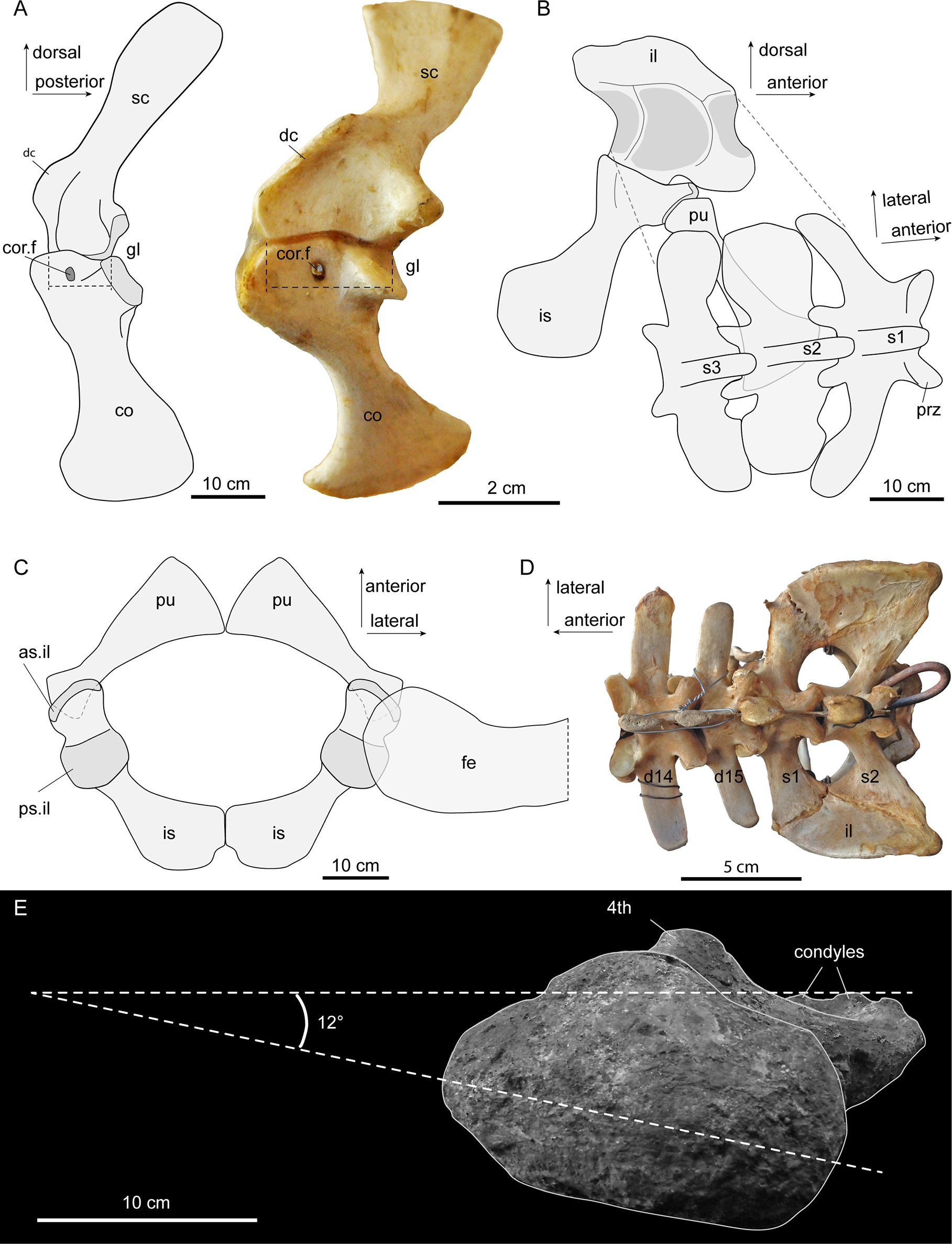
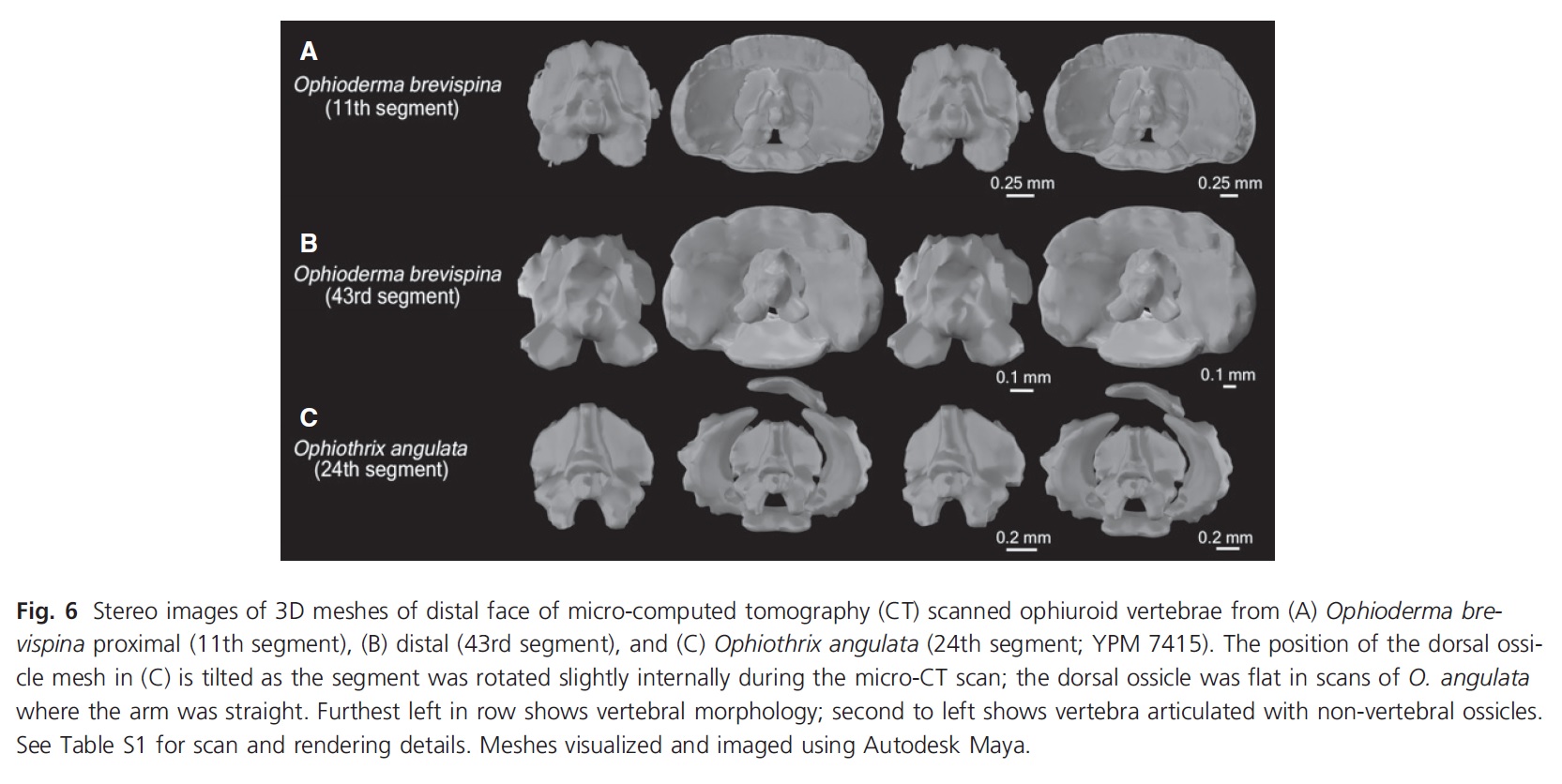
Three-vertebra sacrum and other traits of Purussaurus; with living caiman bones for comparison. E (bottom): inwards-facing femur head. (see paper for more info)
It became evident that, together, those odd traits conveyed a signal that the skeleton was transformed to aid in supporting the huge body against gravity. For example, I found it quite interesting how the head of the femur (thigh bone) was oriented more directly into the hip socket in multiple specimens, more like a dinosaur’s hip, and specialised for support and fore-aft motions. I used Haley O’Brien et al’s data to estimate just how big P. mirandai might have been and it came out as perhaps 3000 kg and 8 metres total length; as we’d thought, among the largest Crocodylia (and there are larger Purussaurus known, too).

Reconstruction of Purussaurus and morphology of the girdles. (see paper for more info)
The team also put a cool “evo-devo-biomechanics” spin on the study. It is well known that the regional identities of vertebrae (e.g. neck, trunk, sacrum, tail) are largely determined by Hox (homeobox) regulatory genes, early in development. So changes of vertebral identity intimate changes of genetic controls. Crocodylia don’t normally add a trunk vertebra to their sacrum, and only a few fossil crocodyliforms (extinct cousins) ever did either, but we noticed that some specimens of Crocodylia would at least partially make this transformation in pathological states (below). Hence the controls to make these changes exist and sometimes manifest in living crocs, but it’s probably not an “easy” transformation to achieve. One could speculate that under intense selection, such as that imposed by giant body size and some degree of activity on land, that transformation could more easily get permanently “fixed” in a species.

Palaeosuchus palpebrosus (Cuvier’s dwarf caiman) with pathological partial-three-vertebra-sacrum; and lots more morphology. (see paper for more info)
As a nice tie-in to the asymmetrical gait study above, we can safely infer that the giant Purussaurus wasn’t a fast animal on land, by any means. But its skeleton is consistent with it having found novel ways to maintain the ability to stand and move on land, even if slowly.
Happy holidays! Santa Jaws is watching you– be good!