To me, there is no question that the Galerie de Paléontologie et d’Anatomie comparée of Paris’s Muséum national d’Histoire naturelle (MNHN) is the mecca of organismal anatomy, as their homepage describes. Georges Cuvier got the morphological ball rolling there and numerous luminaries were in various ways associated with it too; Buffon and Lamarck and St Hiliaire to name but a few early ones. It is easy to think of other contenders such as the NHMUK in London (i.e., Owen), Jena in Germany, the MCZ at Harvard (e.g. Romer) and so forth. But they don’t quite cut the dijon.
As today is John’s Freezer’s 7th blogoversary, and I was just at the MNHN in Paris snapping photos of their mecca, it’s time for an overdue homage to the magnificent mustard of that maison du morphologie. The exhibits have little signage and are an eclectic mix of specimens, but this adds to its appeal and eccentricity for me. I’ve chosen some of my favourite things I saw on exhibit on this visit, with a focus on things that get less attention (NO MESOZOIC DINOSAURS! sorry), are just odd, or otherwise caught my fancy. It’s a photo blog post, so I shall shut up now, much as I could gush about this place. I could live here.
Need plus-grand images? Clic!
Stomach-Churning Rating: 7/10 for some potentially disturbing anatomical images such as viscera, preserved bits, models of naughty bits etc.
Greetings. Note the stomach-churning rating above, please.
Right. We’ll get the amazing first view as one steps into the gallery done first. Mucho mecca. Anatomy fans simply must go here at least once in their life to experience it, and one cannot ever truly absorb all the history and profound, abundant details of morphology on exhibit.
Less-often-seen views from the balcony; one more below.
Indian Rhinoceros from Versailles’s royal menagerie; came to the MNHN in 1792.
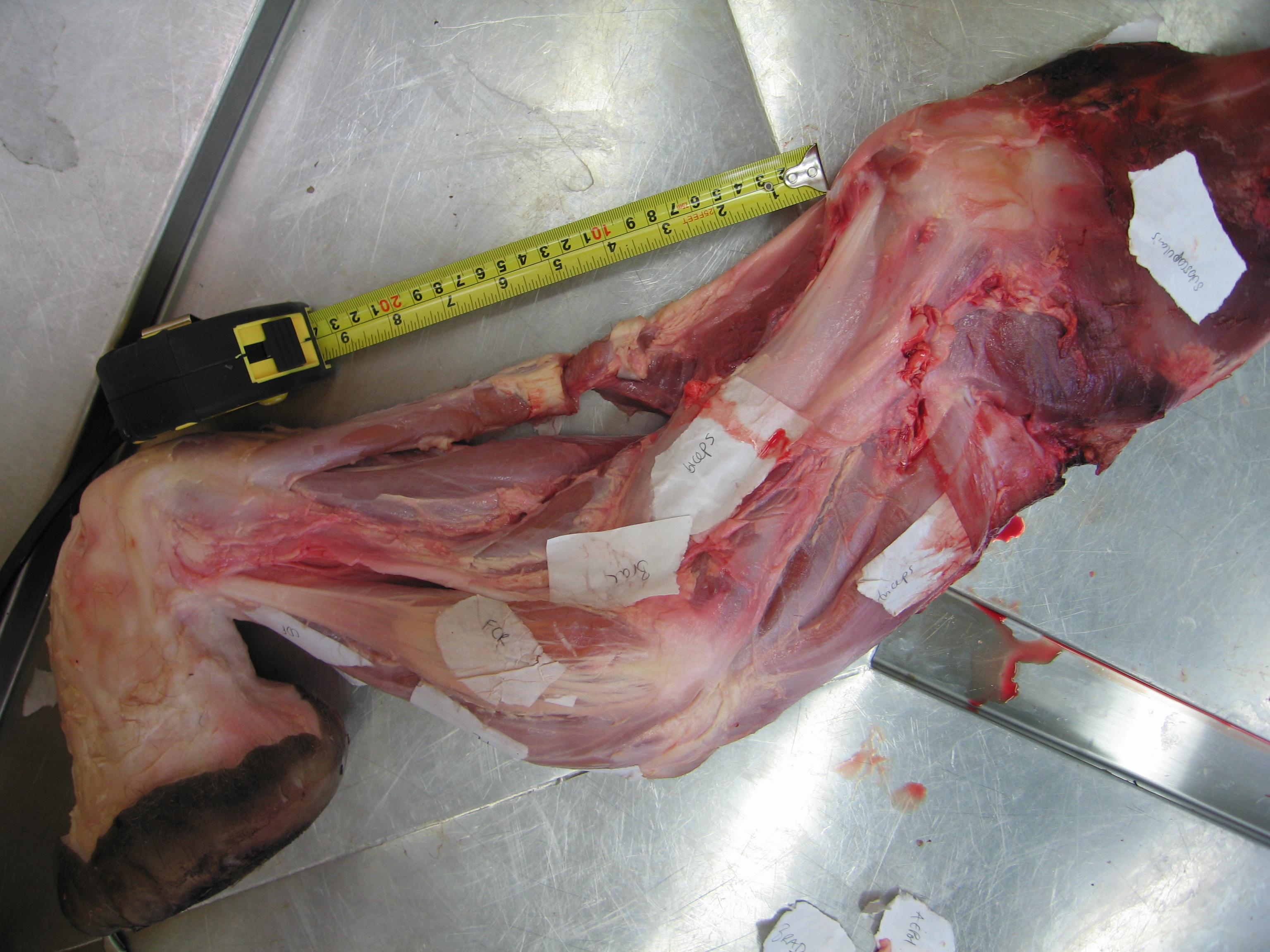
Brown bear hindlimb bones.
Brown bear forelimb bones and pelvis.
Two baby polar bears; part of the extensive display of ontogeny (too often missing in other museums’ exhibits).
Asian elephant from Sri Lanka.
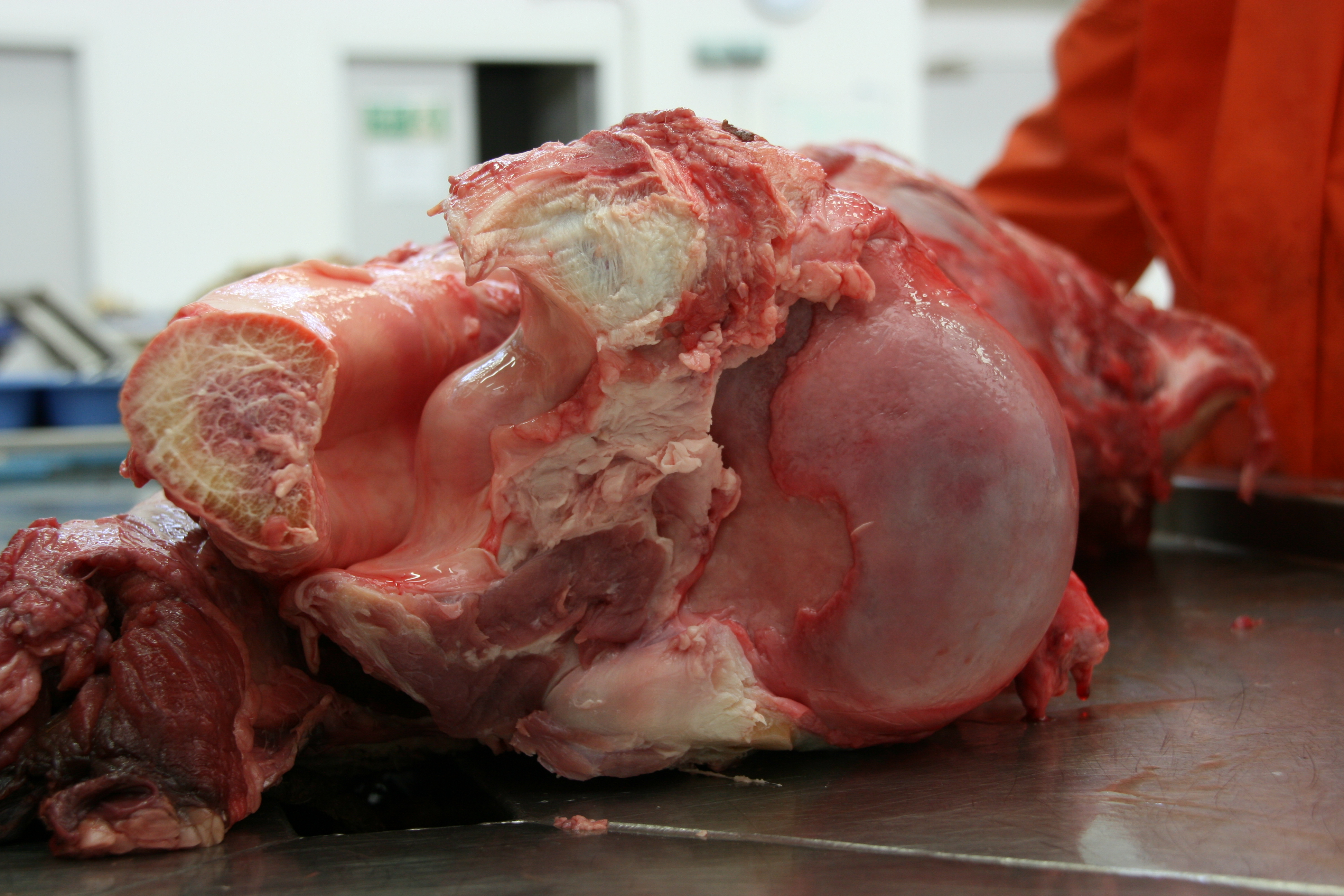
Lamb birth defect. Like ontogeny, pathology was a major research interest in the original MNHN days.
Wild boar birth defect.
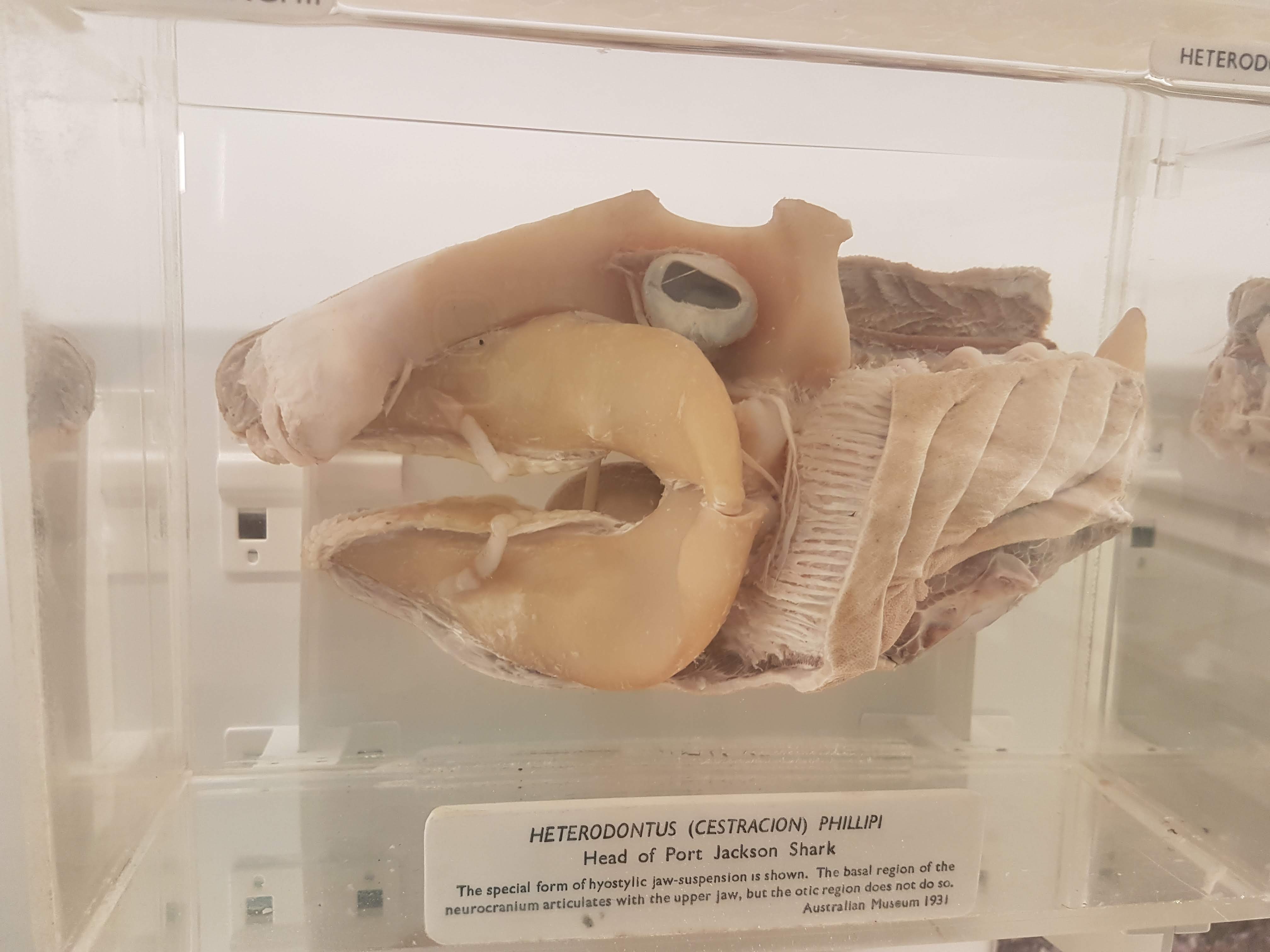
Fabulous large Indian gharial skull + skeleton.
“Exploded” Nile crocodile skull to show major bones.
Let’s play name-all-the-fish-skull-bones, shall we?
Rare sight of a well-prepared Mola mola ocean sunfish skeleton.
Diversity of large bird eggs.
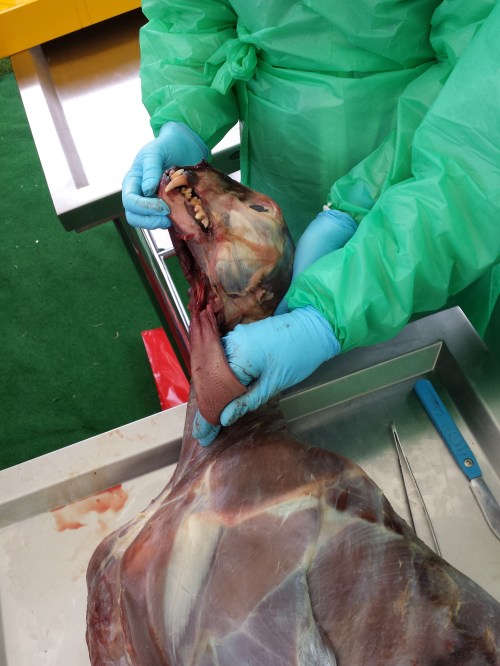
Asian musk deer (male), with tooth roots exposed.
Freaky gorilla is here to say that now the really odd specimens begin, including the squishy bits.
Freaky tamandua, to keep freaky gorilla company. Displaying salivary glands associated with the tongue/pharynx. These are examples of anatomical preparations using older analogues of plastination, such as papier-mâché modelling. I’m not completely sure how the preservation was done here.
Simpler stomach of a wolf. Not much room for Little Red Riding Hood, I’m afraid.
Expansive surface area of a hippo’s stomach; but not a multi-chambered ruminant gut.
Cervical air sacs of a Turquoise-fronted Amazon parrot.
Heart and rather complex pulmonary system of a varanid lizard.
It’s pharynx time: Keratinous spines of a sea turtle’s throat. All the better to grip squids or jellies!
Pharynx convergent evolution in a giraffe: keratinous spines to help grip food and protect the pharynx from spiny acacia thorns while it passes down the long throat.
Tongue/hyoid region of the pharynx of a varanid, showing the forked tongue mechanism.
Palaeontological awesomeness on the upper floor (the 2nd part of the gallery’s name). Here, the only Siberian woolly mammoth, I’m told, to have left Russia for permanent display like this. Frozen left side of face, here, and 2 more parts below.
Extinct rhino Diaceratherium, with a pathological ankle (degenerative joint disease). I love spotting pathologies in specimens- it makes them stand out more as individuals that lived a unique life.
Glyptodont butt and thagomizer, to begin our tour of this business-end weaponry.
Eutatus leg bones, from a large fossil armadillo; Argentina. Really odd morphology; Xenarthrans are so cool.
I had a spare hour in Cambridge this weekend so I dared the crowds in the revamped UMZC’s upper floor. In my prior visit and post I’d experienced and described the lower floor, which is almost exclusively mammals. This “new” floor has everything else that is zoological (animal/Metazoa) and again is organized in an evolutionary context. And here is my photo tour as promised!
Inviting, soft lighting perfuses the exhibits from the entryway onwards.
All images can be clicked to mu-zoom in on them.
Stomach-Churning Rating: 5/10 for spirit animals, by which I mean dissected/ghostly pale whole specimens of animals in preservative fluids.
The exhibits are on a square balcony overlooking the lower floor, so you can get some nice views. It does make the balcony crowded when the museum is busy, so take that in mind if visiting. Strollers on this upper floor could be really difficult. But the ceiling is very tall so it is not cramped in a 3D sense. The lower floor is more spacious.
Like phylogenies? You got em! Tucked away at the beginning of each major group; not occupying huge valuable space or glaringly obvious like AMNH in NYC but still noticeable and useful. To me, it strikes a good balance; gives the necessary evolutionary context for the displayed specimens/taxa.
Introductory panels explain how names are given to specimens, how specimens are preserved and more.
The exhibits give due focus to research that the UMZC is doing or has been famous for. Hey I recognize that 3D tetrapod image in the lower left! 🙂
There is ample coverage of diversity throughout Metazoa but my camera tended to be drawn to the Vertebrata. Except in some instances like these.
Some larger chelicerates.
Some smaller, shadowy sea scorpion (eurypterid) fossils.
Watch here for more about ophiuroids (brittlestars) in not too long!
A BIG fish brain! Interesting! Before I go through specimens in evolutionary “sequence”, I will feature another thing i really liked: lots of dissected spirit-specimens that show off cool anatomy/evolution/adaptation (and technical skills in anatomical preparation). Mostly heads; mostly fish.
Salps and other tunicates! Our closest non-vertebrate relatives- and some insight into how our head and gut came to be.
Salp-reflection.
Lamprey head: not hard to spot the commonalities with the salps; but now into Vertebrata.
Hagfish head: as a fellow cyclostome/agnathan, much like a lamprey but never forget the slime glands!
Shark head. Big fat jaws; all the better to bite prey with!
Lungfish (Protopterus) head showing the big crushing tooth plates (above).
Sturgeon vertebrae: tweak some agnathan/shark bits and here you are.
Worm (annelid) anatomy model, displaying some differences from/similarities to Vertebrata. (e.g. ventral vs. dorsal nerve cord; segmentation)
Dissected flipper from a small whale/other cetacean. Still five fingers, but other specializations make it work underwater.
Wonderful diversity of tooth and jaw forms in sharks, rays and relatives. I like this display a lot.
More of the above, but disparate fossil forms!
On with the evolutionary context! Woven throughout the displays of modern animals are numerous fossils, like these lovely placoderms (lineage interposed between agnathans, sharks and other jawed fish).
Goblin shark head.
I seem to always forget what ray-finned fish this is (I want to say wolffish? Quick Googling suggests maybe I am right), but see it often and like its impressive bitey-ness.
Bichir and snakefish; early ray-finned fish radiations.
Armoured and similar fish today.
Armoured fish of the past; some convergent evolution within ray-fins.
Convergence- and homology- of amphibious nature in fish is another evolutionary pattern exemplified here.
Gorgeous fossils of ray-finned fish lineages that arose after the Permian extinctions, then went extinct later in the Triassic.
Note the loooooong snout on this cornetfish but the actual jaws are just at the tip.
Flying fish– those ray-fins are versatile.
Diversity of unusual ray-finned fish, including deep-water and bottom-dwelling forms.
Can you find the low-slung jaws of a dory?
Recent and fossil perch lineage fish.
It’s hard to get far into talking about evolution without bringing up the adaptive radiation of east African cichlid fish, and UMZC researchers are keen on this topic too.
Lobe-fins! Everybody dance!
Rhizodonts & kin: reasons to get out of Devonian-Carboniferous waters.
A Cretaceous fossil coelacanth (skull); not extremely different from living ones’.
Let’s admire some fossil and modern lungfish skulls, shall we? Big platey things (here, mainly looking at the palate) with lots of fusions of tiny bones on the skull roof.
Eusthenopteron fossils aren’t that uncommon but they are still great to see; and very important, because…
OK let’s stop messing around. The UMZC has one of the best displays of fossil stem-tetrapods in the world! And it should.
Another look at the pretty Acanthostega models.
Acanthostega vs. primate forelimb: so like us.
Ichthyostega parts keep Acanthostega company.
A closer look at the “Mr. Magic” Ichthyostega specimen, which takes some unpacking but is incredibly informative and was a mainstay of our 2012 model. Back of skull, left forelimb, and thorax (from left to right here).
Eucritta, another stem-tetrapod.
Closer look at Eucritta‘s skull.
Weird stem-tetrapod Crassigyrinus, which we’re still trying to figure out. It’s a fabulous specimen in terms of completeness, but messy “roadkill” with too many damn bones.
The large skull of Crassigyrinus, in right side view.
Early temnospondyl (true amphibian-line) skulls and neck.
Nectrideans or the boomerangs of the Palaeozoic.
Cool fossil frogs.
Giant Japanese salamander!
Fire salamanders: not as colourful as the real thing, but here revealing their reproductive cycle in beautiful detail.
Closeup of oviduct in above.
Sexual dimorphism in Leptodactylus frogs: the males have bulging upper arms to (I am assuming) help them hold onto females during amplexus (grasping in mating competitions).
Did I forget that Leptodactylus has big flanges on the humerus in males, to support those muscles? Seems so.
An early stem-amniote, Limnoscelis (close to mammals/reptiles divergence); cast.
Grand sea turtle skeleton.
One of my faves on display: a real pareiasaurian reptile skeleton, and you can get a good 3D look around it.
Details on above pareiasaurian.
Mammals are downstairs, but we’re reminded that they fit into tetrapod/amniote evolution nonetheless.
Let there be reptiles! And it was good.
Herps so good. (slow worm, Gila monster, glass lizard)
A curator is Dr Jason Head so you bet Titanoboa is featured!
Crocodylia: impressive specimens chosen here.
It ain’t a museum without a statuesque ratite skeleton. (There are ~no non-avian dinosaurs here– for those, go to the Sedgwick Museum across the street, which has no shortage!)
Avian diversity takes off.
Glad to see a tinamou make an appearance. They get neglected too often in museums- uncommon and often seemingly unimpressive, but I’m a fan.
I still do not understand hoatzins; the “cuckoo” gone cuckoo.
Dodo parts (and Great Auk) near the entrance.
Wow. What an oilbird taxidermy display! :-O
There we have it. Phew! That’s a lot! And I left out a lot of inverts. This upper floor is stuffed with specimens; easier there because the specimens are smaller on average than on the lower floor. Little text-heavy signage is around. I give a thumbs-up to that– let people revel in the natural glory of what their eyes show them, and give them nuggets of info to leave them wanting more so they go find out.
Now it’s in your hands– go find out yourself how lovely this museum is! I’ve just given a taste.
As 2017 approaches its end, there have been a few papers I’ve been involved in that I thought I’d point out here while I have time. Our DAWNDINOS project has been taking up much of that time and you’ll see much more of that project’s work in 2018, but we just published our first paper from it! And since the other two recent papers involve a similar theme of muscles, appendages and computer models of biomechanics, they’ll feature here too.
Stomach-Churning Rating: 0/10; computer models and other abstractions.
Mussaurus patagonicus was an early sauropodomorph dinosaur from Argentina, and is now widely accepted to be a very close relative of the true (giant, quadrupedal) sauropods. Here is John Conway’s great reconstruction of it:
We have been working with Alejandro Otero and Diego Pol on Mussaurus for many years now, starting with Royal Society International Exchange funds and now supported by my ERC grant “DAWNDINOS”. It features in our grant because it is a decent example of a large sauropodomorph that was probably still bipedal and lived near the Triassic-Jurassic transition (~215mya).
In our new study, we applied one of my team’s typical methods, 3D musculoskeletal modelling, to an adult Mussaurus’s forelimbs. This is a change of topic from the hindlimbs that I’ve myopically focused on before with Tyrannosaurus and Velociraptor [in an obscure paper that I should never have published in a book! pdf link], among other critters my team has tackled (mouse, elephant [still to be finished…], ostrich, horse, Ichthyostega… dozens more to come!). But we also modelled the forelimbs of Crocodylus johnstoni (Australian “freshie”) for a key comparison with a living animal whose anatomy we actually knew, rather than reconstructed.
Mussaurus above; Crocodylus below; forelimb models in various views; muscles are red lines.
The methods for this biomechanical modelling are now standard (I learned them from their creator Prof. Scott Delp during my 2001-2003 postdoc at Stanford): scan bones, connect them with joints, add muscle paths around them, and then use the models to estimate joint ranges of motion and muscle moment arms (leverage) around joints. I have some mixed feelings about developing this approach in our 2005 paper that is now widely used by the few teams that study appendicular function in extinct animals. As a recent review paper noted and I’ve always cautioned, it has a lot of assumptions and problems and one must exercise extreme caution in its design and interpretation. Our new Mussaurus paper continues those ruminations, but I think we made some progress, too.
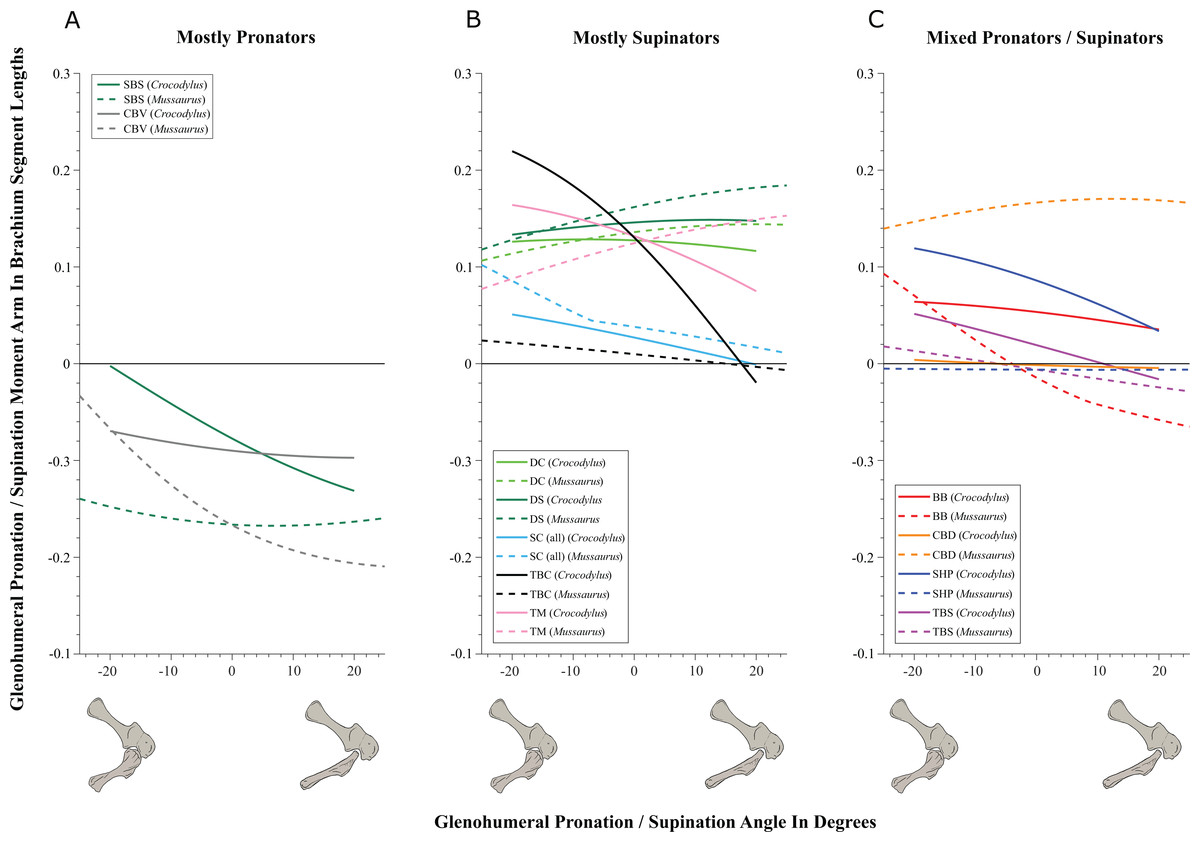
On to the nuts and bolts of the science (it’s a 60 page paper so this summary will omit a lot!): first, we wanted to know how the forelimb joint ranges of motion in Mussaurus compared with those in Crocodylus and whether our model of Mussaurus might be able to be placed in a quadrupedal pose, with the palms at least somewhat flat (“pronated”) on the ground. Even considering missing joint cartilage, this didn’t seem very plausible in Mussaurus unless one allowed the whole forearm to rotate around its long axis from the elbow joint, which is very speculative—but not impossible in Crocodylus, either. Furthermore, the model didn’t seem to have forelimbs fully adapted yet for a more graviportal, columnar posture. Here’s what the model’s mobility was like:
So Mussaurus, like other early sauropodomorphs such as Plateosaurus, probably wasn’t quadrupedal, and thus quadrupedalism must have evolved very close to in the Sauropoda common ancestor.
Second, we compared the muscle moment arms (individual 3D “muscleactions” for short) in different poses for all of the main forelimb muscles that extend (in various ways and extents) from the pectoral girdle to the thumb, for both animals, to see how muscle actions might differ in Crocodylus (which would be closer to the ancestral state) and Mussaurus. Did muscles transform their actions in relation to bipedalism (or reversal to quadrupedalism) in the latter? Well, it’s complicated but there are a lot of similarities and differences in how the muscles might have functioned; probably reflecting evolutionary ancestry and specialization. What I found most surprising about our results was that the forelimbs didn’t have muscles well-positioned to pronate the forearm/hand, and thus musculoskeletal modelling of those muscles reinforced the conclusions from the joints that quadrupedal locomotion was unlikely. I think that result is fairly robust to the uncertainties, but we’ll see in future work.
You like moment arms? We got moment arms! 15 figures of them, like this! And tables and explanatory text and comparisons with human data and, well, lots!
If you’re really a myology geek, you might find our other conclusions about individual muscle actions to be interesting—e.g. the scapulohumeralis seems to have been a shoulder pronator in Crocodylus vs. supinator in Mussaurus, owing to differences in humeral shape (specialization present in Mussaurus; which maybe originated in early dinosaurs?). Contrastingly, the deltoid muscles acted in the same basic way in both species; presumed to reflect evolutionary conservation. And muuuuuuch more!
Do you want to know more? You can play with our models (it takes some work in OpenSim free software but it’s do-able) by downloading them (Crocodylus; Mussaurus; also available: Tyrannosaurus, Velociraptor!). And there will be MUCH more about Mussaurus coming soon. What is awesome about this dinosaur is that we have essentially complete skeletons from tiny hatchlings (the “mouse lizard” etymology) to ~1 year old juveniles to >1000kg adults. So we can do more than arm-wave about forelimbs!
But that’s not all. Last week we published our third paper on mouse hindlimb biomechanics, using musculoskeletal modelling as well. This one was a collaboration that arose from past PhD student James Charles’s thesis: his model has been in much demand from mouse researchers, and in this case we were invited by University of Virginia biomechanical engineers to join them in using this model to test how muscle fibres (the truly muscle-y, contractile parts of “muscle-tendon units”) change length in walking mice vs. humans. It was a pleasure to re-unite in coauthorship with Prof. Silvia Blemker, who was a coauthor on that 2005 T. rex hindlimb modelling paper which set me on my current dark path.
Mouse and human legs in right side view, going through walking cycles in simulations. Too small? Click to embiggen.
We found that, because mice move their hindlimb joints through smaller arcs than humans do during walking and because human muscles have large moment arms, the hindlimb muscles of humans change length more—mouse muscles change length only about 48% of the amount that typical leg muscles do in humans! This is cool not only from an evolutionary (mouse muscles are probably closer to the ancestral mammalian state) and scaling (smaller animals may use less muscle excursions, to a point, in comparable gaits?) perspective, but it also has clinical relevance.
Simulated stride for mouse and human; with muscles either almost inactive (Act=0.05) or fully active (Act=1). Red curve goes through much bigger excursions (along y-axis) than blue curve), so humans should use bigger % of their muscle fibre lengths in walking. Too small? Click to embiggen.
My coauthors study muscular dystrophy and similar diseases that can involve muscle stiffness and similar biomechanical or neural control problems. Mice are often used as “models” (both in the sense of analogues/study systems for animal trials in developing treatments, and in the sense of computational abstractions) for human diseases. But because mouse muscles don’t work the same as human muscles, especially in regards to length changes in walking, there are concerns that overreliance on mice as human models might cause erroneous conclusions about what treatments work best to reduce muscle stiffness (or response to muscle stretching that causes progressive damage), for example. Thus either mouse model studies need some rethinking sometimes, or other models such as canines might be more effective. Regardless, it was exciting to be involved in a study that seems to deliver the goods on translating basic science to clinical relevance.
Muscle-by-muscle data; most mouse muscles go through smaller excursions; a few go through greater; some are the same as humans’.
Finally, a third recent paper of ours was led by Julia Molnar and Stephanie Pierce (of prior RVC “Team Tetrapod” affiliation), with myself and Rui Diogo. This study tied together a bunch of disparate research strands of our different teams, including musculature and its homologies, the early tetrapod fossil record, muscle reconstruction in fossils, and biomechanics. And again the focus was on forelimbs, or front-appendages anyway; but turning back the clock to the very early history of fishes, especially lobe-finned forms, and trying to piece together how the few pectoral fin muscles of those fish evolved into the many forelimb muscles of true tetrapods from >400mya to much more recent times.
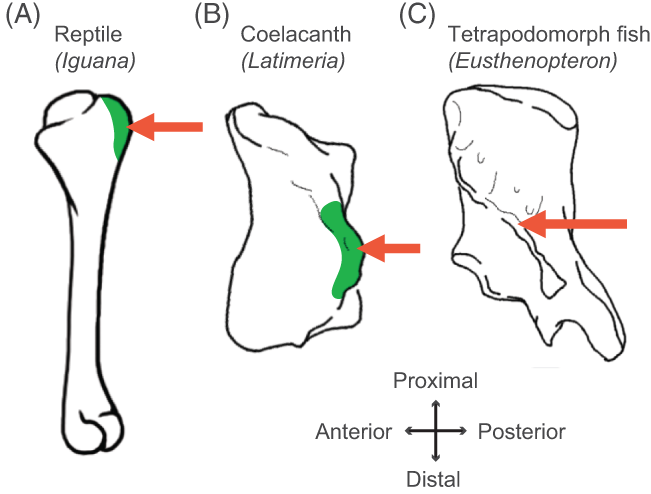
Humerus in ventral view, showing muscle attachments. Extent (green) is unknown in the fossil but the muscle position is clear (arrow).
We considered the homologies for those muscles in extant forms, hypothesized by Diogo, Molnar et al., in light of the fossil record that reveals where those muscles attach(ed), using that reciprocal illumination to reconstruct how forelimb musculature evolved. This parallels almost-as-ancient (well, year 2000) work that I’d done in my PhD on reconstructing hindlimb muscle evolution in early reptiles/archosaurs/dinosaurs/birds. Along the way, we could reconstruct estimates of pectoral muscles in various representative extinct tetrapod(omorph)s.
Disparity of skeletal pectoral appendages to work with from lobe-fins to tetrapods.
Again, it’s a lengthy, detailed study (31 pages) but designed as a review and meta-analysis that introduces readers to the data and ideas and then builds on them in new ways. I feel that this was a synthesis that was badly needed to tie together disparate observations and speculations on what the many, many obvious bumps, squiggles, crests and tuberosities on fossil tetrapods/cousins “mean” in terms of soft tissues. The figures here tell the basic story; Julia, as usual, rocked it with some lovely scientific illustration! Short message: the large number of pectoral limb muscles in living tetrapods probably didn’t evolve until limbs with digits evolved, but that number might go back to the common ancestor of all tetrapods, rather than more recently. BUT there are strong hints that earlier tetrapodomorph “fishapods” had some of those novel muscles already, so it was a more stepwise/gradual pattern of evolution than a simple punctuated event or two.
Colour maps of reconstructed right fin/limb muscles in tetrapodomorph sarcopterygian (~”fishapod”) and tetrapod most recent common ancestors. Some are less ambiguous than others.
That study opens the way to do proper biomechanical studies (like the Mussaurus study) of muscle actions, functions… even locomotor dynamics (like the mouse study)– and ooh, I’ve now tied all three studies together, tidily wrapped up with a scientific bow! There you have it. I’m looking forward to sharing more new science in 2018. We have some big, big plans!
We’d been wanting to do a family holiday in Ireland for years and so we finally did. I’d been to Dublin twice before for work visits and we wanted a more rural experience. On others’ recommendations, we started in the city of Cork. With some sleuthing and asking around, I realized that we weren’t far then from gorgeous Killarney National Park, and then it wasn’t far west from there to get to Valentia Island, where incidentally there is something amazing for palaeontology-lovers. There was no deterring me at that point from visiting what I’d only read about. I’ll mainly let the images tell the story.
Stomach-Churning Rating: 0/10; fossils and scenery. Kick back and enjoy.
Island map- it really is that simple to get around! The harbour town of Portmagee is damned adorable.
Driving in (no I am the passenger; not taking photo while at the wheel!)- excitement level = 8 and building… “Tetrapod carpark” sign ratcheted up the excitement and was amusing.
Headed to the trail; excitement level = 9…
Looking down onto the site (on the right); excitement level = 9.5; beauty level = 9.5 too!
Now, the site of what is broadly accepted by experts as a ~Middle Devonian tetrapod(omorph)’s fossil trackway(s) was originally described by Stössel in 1995. To me, that feels like a recent discovery but it is 22 years ago. The only other well-preserved, widely-accepted, probably-terrestrial, Late Devonian tetrapod(omorph) trackways are from the Genoa River site in Australia; described by Warren et al. in 1972. Those trackways even reveal some details of the fingers and toes; these do not. Other tracks are either isolated footprints of minimal scientific value/clarity, subaerial (i.e. underwater), not clearly stem-tetrapod (or now argued to be arthropod or other origin), not Devonian, or controversial for reasons I won’t get into here. The famous Zachelmie tracks in Poland are strong contenders but remain controversial to more than a few researchers in terms of who made them and in what environmental/substrate context; but their Middle Devonian age seems robustly agreed. Clack and Lucas have reviewed the relevant evidence recently. So there are essentially at least two, and arguably three, places in the world that you can visit to view tracks like these and it was a joy to go visit one set. (Easter Ross, Scotland may be a fourth site but it is reasonably disputed in age and maker)
There is a “however,” however- Falkingham and Horner showed how lungfish can produce tracks (with fins and heads together) that look like these, to some viewers (but not to others) and in some substrates (mud; not sand as at the Valentia site)– so there is still uncertainty for some tracks although the lungfish-feeding claims have also been vehemently disputed too. Without finger and toe impressions, claims of discrete tetrapod tracks are risky, and it would be wrong to say that the Valentia Island footprints are uncontroversially or 100% certainly tetrapod in origin, although they are (late-Middle) Devonian and made by some sort of animal, and very likely a tetrapodomorph at least.
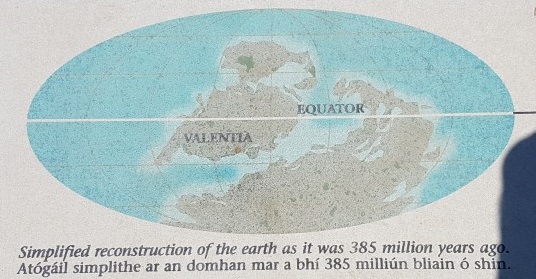
Stössel et al. also published a recent update on these Valentia Island tracks with more information. I wish I’d come across that before I visited (oops!). That study reports on a total of nine(!) trackways from the area, adding to the 1995’s first one (the “Dohilla locality, Do 1”– see diagrams below), and describes them as Middle Devonian (with a radiometric dating of 385 million years old). I’m not enough of a geologist to evaluate that; prior reports had focused on Late Devonian or so.
Rippled sandstone example; preservation characteristic of the trackway area/Valentia Slate Formation. It’s an alluvial deposit (freshwater floodplain), interpreted to lie inland from the coastal marine deposits. Raindrop impressions above the plane of the tracks raise the possibilities that the tracks were made on (moist) land.
The island has plenty of signs advertising the tracks as a tourist destination but happily(?) there are no knick-knack shops stocked with plush tetrapods, or other developments at or near the site. One simply winds down a very narrow road near a radio station and old lighthouse, and parks then walks to see the tracks. No fancy crap; just AWESOME sights to take in, and some good educational information.
Explanatory plaque at the viewing area. Pretty good!
Nice image of where Valentia Island was; although the 385 My age may be exaggerated. It’s not clear how old the tracks are but “Mid-to-Late Devonian” might suffice depending on how you view the evidence. The tracks were the “oldest known” at the time of discovery and remain close to that, but challenged by the Zachelmie trackways (see references above).
Explanatory signs on the peak above the shore. Given the likely tetrapod(omorph) trackmakers like Acanthostega-style critters, the adult animal may have been able to breathe air with lungs and underwater with gills.
Enough exposition– let’s expose those tracks! (images can be clicked to enlarge)
My first close-up look at the tracks. Whoa! Small tracks are presumably hand (manus) impressions; larger ones are foot (pes). The tracks go in an alternating fashion (like a salamander’s walk) and the animal was probably going from the bottom-right toward the top-left. Moss and moisture obscured some of the prints that day, sadly. The tracks are oval, with the long axes perpendicular to the direction of travel. There are some pesky geological deformations of the trackway, faults, and other distortions. 145 footprints in total are reported from this one trackway!
Trackway as it turns to the left and gets harder to follow. John-shadow for ~scale. Frustratingly for me, a little rivulet was coming down the hill across the left side of the trackway and hiding much of the detail of the end.
Alternative view of the majority of the tracks; turned ~90 degrees from above two views.
Zoomed-in view of the tracks from head-on (opposite the view in other photos); i.e. western position looking east (ish). I added red and blue dots to roughly outline the right side of the main trackway (red) and the second one (blue), which crosses it and may have been made after it.
Even these nice trackways, viewed by an expert, take some unpacking. Here is some:
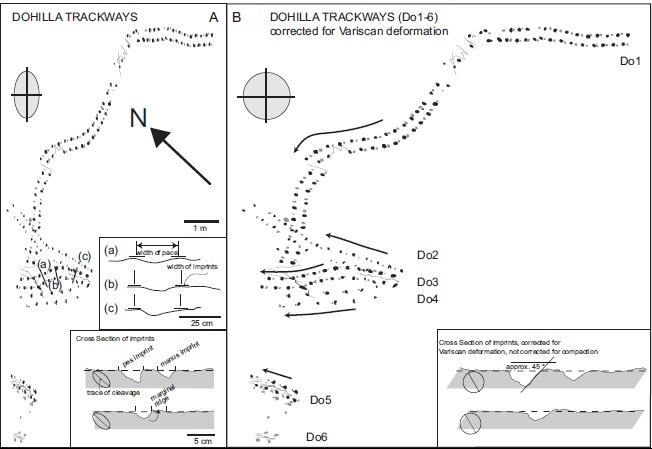
Diagram of known tracks at the site by Stössel et al. 2016.
Diagram at view site with extra tail (or body) drag trail crossing the main tracks; described later by Stössel et al. 2016.
I’m not at all a religious person and I don’t really like the term “spiritual” either, but this experience was emotional for me. Awe is certainly the best word to describe what I felt on viewing these tracks. The literature just doesn’t do them justice; nothing beats a first-person experience like this. We were lucky with excellent weather, too, and we were almost alone during the visit so there was pleasant silence in which to contemplate the tracks. I brought my copies of three papers on the trackways and, struggling with the wind, compared them with the visible tracks to understand what other scientists had seen. That amplified the experience enormously for me.
Even if they turn out to be non-tetrapod or younger or something less exciting (“sham-rock”?), it was thrilling to see the Valentia Island tracks and think about what happened ~385 million years ago when they were made by our very distant cousins, along the land-water interface of space and time.
(I also feel bad for online reviewers that were disappointed with the site- it’s hard to grasp the scientific importance and/or accept the evidence, even with the decent information available on-site. Even if people know the nice fossil record of dinosaurs, they may not know how good the fossil record of early tetrapods is and how confidently we can figure out what happened in the Devonian emergence of tetrapod(omorph)s onto land. But some visitors clearly got it.)
And, looking at the site myself, I realized how many more tracks might be buried under the cliffs of the site- the first trackway emerges from under a cliff and thus must still be preserved for some distance underground, awaiting future exposure. What more might we learn about that single animal and others that made tracks around the same time? I hope to live to find out. I feel a personal connection now to these tracks, left pondering what story they preserve– and hide. I’m glad I’m able to share my own story with you, and encourage you to make the visit yourself!
Last year on Darwin Day I debuted “Better Know A Muscle” (BKAM), which was intended to be a series of posts focusing on one cool muscle at a time, and its anatomical, functional and evolutionary diversity and history. A year later, it’s another post on another muscle! Several dozen more muscles to go, so I’ve got my work cut out for me… But today: get ready to FLEX your myology knowledge! Our subject is Musculus biceps brachii; the “biceps” (“two-headed muscle of the arm”). Beloved of Arnie and anatomists alike, the biceps brachii is. Let’s get pumped up!
Stomach-Churning Rating: 7/10. Lots of meaty elbow flexion!
While the previous BKAM’s topic was a hindlimb muscle with a somewhat complex history (and some uncertainties), the biceps brachii is a forelimb muscle with a simpler, clearer history. Fish lack a biceps, just having simple fin ab/adductor muscles with little differentiation. Between fish and tetrapods (limb-bearing vertebrates), there was an explosion in the number of muscles; part of transforming fins into limbs; and the biceps is thenceforth evident in all known tetrapods in a readily identifiable anatomical form. In salamanders and their amphibian kin, there is a muscle usually called “humeroantebrachialis” that seems to be an undivided mass corresponding to the biceps brachii plus the brachialis (shorter humerus-to-elbow) muscle:
Most of the humerobrachialis muscle (purplish colour), in dorsal (top) view of the right forelimb of the fire salamander Salamandra salamandra (draft from unpublished work by my team).
In all other tetrapods; the amniote group (reptiles, mammals, etc.); there is a separate biceps and brachialis, so these muscles split up from the ancestrally single “humeroantebrachialis” muscle sometime after the amphibian lineage diverged from the amniotes. And not much changed after then– the biceps is a relatively conservative muscle, in an evolutionary (not political!) sense. In amniote tetrapods that have a biceps, it develops as part of the ventral mass of the embryonic forelimb along with other muscles such as the shorter, humerus-originating brachialis, from which it diverges late in development (reinforcing that these two muscles are more recent evolutionary divergences, too).
Biceps brachialis or humerobrachialis, the “biceps group” tends to originate just in front of the shoulder (from the scapula/coracoid/pectoral girdle), running in front of (parallel to) the humerus. It usually forms of two closely linked heads (hence the “two heads” name), most obviously in mammals; one head is longer and comes from higher/deeper on the pectoral girdle, whereas the other is closer to the shoulder joint and thus is shorter. The two heads fuse as they cross the shoulder joint and we can then refer to them collectively as “the biceps”. It can be harder to see the longer vs. shorter heads of the biceps in non-mammals such as crocodiles, or they may be more or less fused/undifferentiated, but that’s just details of relatively minor evolutionary variation.
The biceps muscle then crosses in front of the elbow to insert mainly onto the radius (bone that connects your elbow to your wrist/thumb region) and somewhat to the ulna (“funny bone”) via various extra tendons, fascia and/or aponeuroses. The origin from the shoulder region tends to have a strong mark or bony process that identifies it, such as the coracoid process in most mammals (I know this well as I had my coracoid process surgically moved!). The insertion onto the radius tends to have a marked muscle scar (the radial tuberosity or a similar name), shared with the brachialis to some degree. A nice thing about the biceps is that, because it may leave clear tendinous marks on the skeleton, we sometimes can reconstruct how its attachments and path evolved (and any obvious specializations; even perhaps changes of functions if/when they happened).
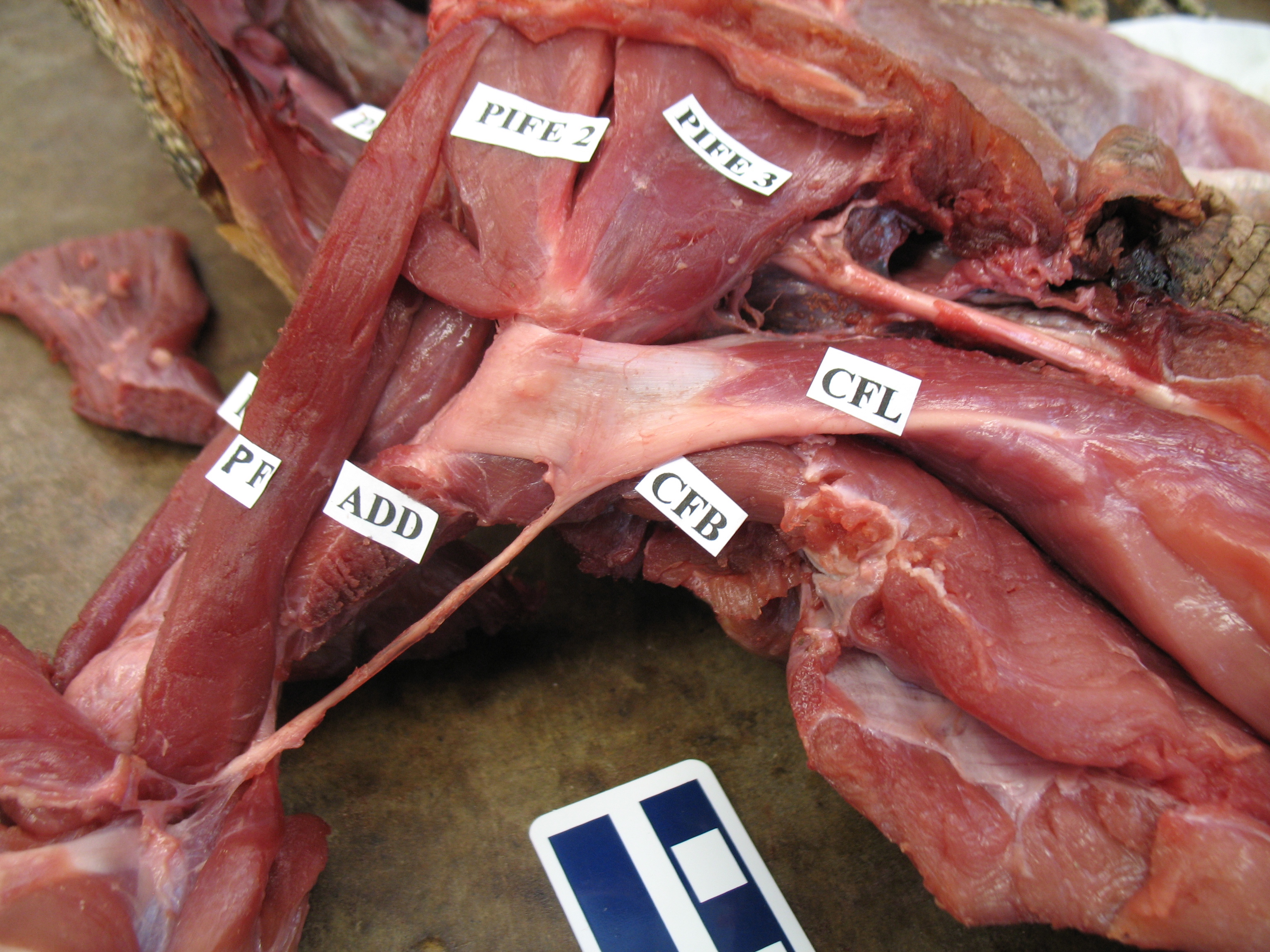
Here are some biceps examples from the world of crocodiles:
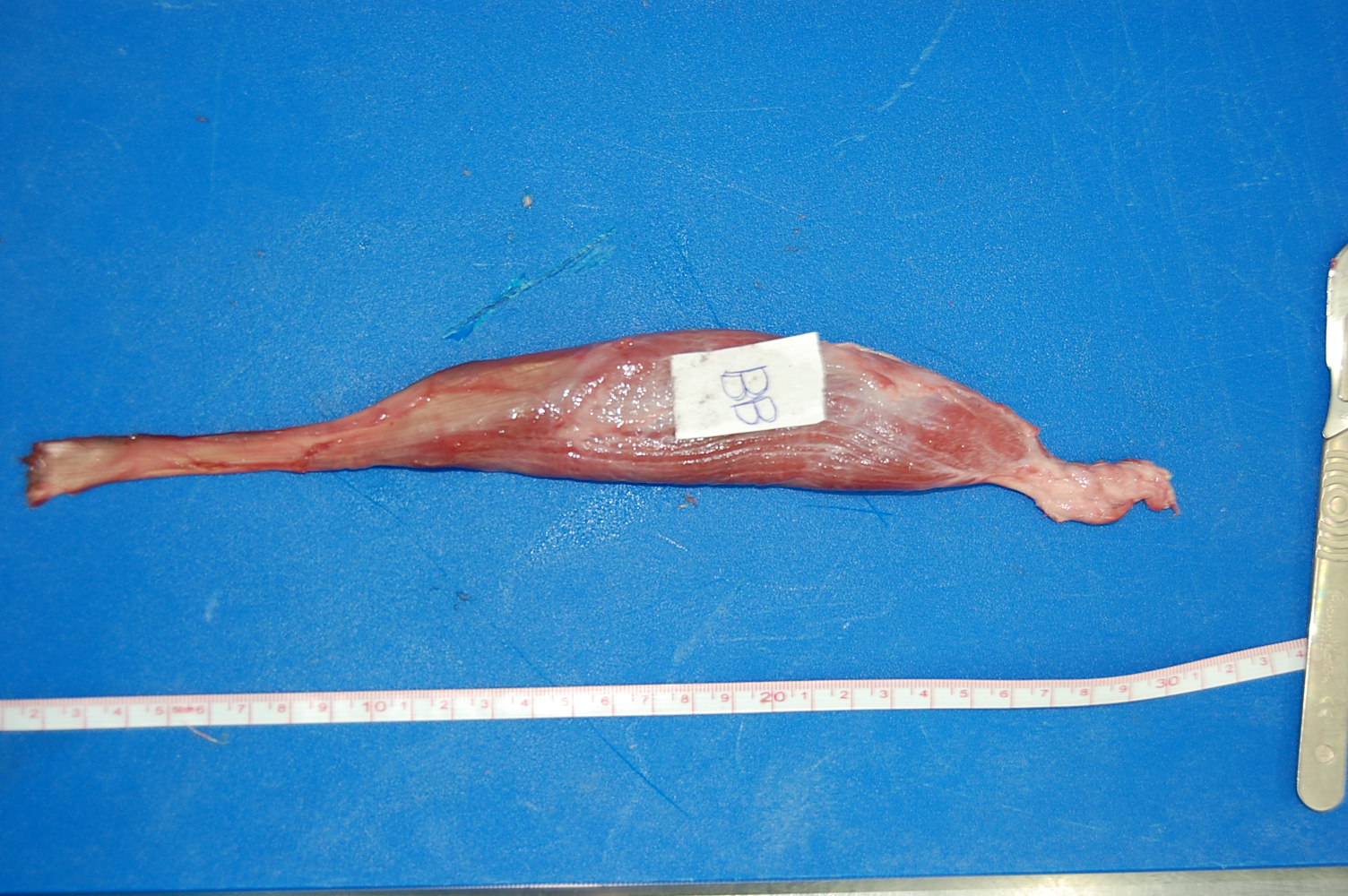
Crocodile’s right forelimb showing the huge pectoralis, and the biceps underlying it; on the bottom right (“BB”- click to embiceps it).
Crocodile left forelimb with biceps visible (“BB”) on the left.
Crocodile biceps muscle cut off, showing the proximal (to right) and distal (to left) tendons (and long parallel muscle fibres) for a typical amniote vertebrate.
What does the biceps muscle do? It flexes (draws forward) the shoulder joint/humerus, and does the same for the elbow/forearm while supinating it (i.e. rotating the radius around the ulna so that the palm faces upwards, in animals like us who can rotate those two bones around each other). In humans, which have had their biceps muscles studied by far the most extensively, we know for example that the biceps is most effective at flexing the elbow (e.g. lifting a dumbbell weight) when the elbow is moderately straight. These same general functions (shoulder and elbow flexion; with some supination) prevail across the biceps muscle of [almost; I am sure there are exceptions] all tetrapods, because the attachments and path of the biceps brachii are so conservative.
And this flexor function of the biceps brachii stands in contrast to our first BKAM muscle, the caudofemoralis (longus): that muscle acts mainly during weight support (stance phase) as an antigravity/extensor muscle, whereas the flexor action of biceps makes it more useful as a limb protractor or “swing phase” muscle used to collapse the limb and draw it forwards during weight support. However, mammals add some complexity to that non-supportive function of the biceps…
Hey mammals! Show us your biceps!
Jaguar forelimb with biceps peeking out from the other superficial muscles, and its cousin brachialis nicely visible, running along the front of the forearm for a bit.
Elephant’s left forelimb with the biceps labelled.
Longitudinal slice thru the biceps of an elephant, showing the internal tendon that helps identify where the two bellies of the biceps fuse.
In certain mammals; the phylogenetic distribution of which is still not clear; the biceps brachii forms a key part of a passive “stay apparatus” that helps keep the forelimb upright against gravity while standing (even sleeping). The classic example is in horses but plenty of other quadrupedal mammals, especially ungulate herbivores, show evidence of similar traits:
Giraffe biceps cut away proximally to show the “stay apparatus” around the shoulder joint (upper right).
Zooming in on the “stay apparatus”; now in proximal view, with the biceps tendon on the left and the humeral head (showing some arthritic damage) on the right, with the groove for the biceps in between.
Hippo’s humerus (upper left) and biceps muscle cut away proximally, displaying the same sort of “stay apparatus” as in the giraffe. Again, note the stout proximal and distal tendons of the biceps. The proximal tendon fits into the groove of the humerus on the far left side of the image; becoming constrained into a narrow circular “tunnel” there. It’s neat to dissect that region because of its fascinating relationships between bone and soft tissues.
The biceps brachii, in those mammals with a stay apparatus, seems to me to have a larger tendon overall, especially around the shoulder, and that helps brace the shoulder joint from extending (retracting) too far backward, whilst also transmitting passive tension down the arm to the forearm, and bracing the elbow (as well as distal joints via other muscles and ligaments). It’s a neat adaptation whose evolution still needs to be further inspected.
Otherwise, I shouldn’t say this but the biceps is sort of boring, anatomically. Whether you’re a lizard, croc, bird or mammal, a biceps is a biceps is a biceps; more or less-ceps. But the biceps still has a clear evolutionary history and Darwin would gladly flex his biceps to raise a pint in toast to it.
So now we know a muscle better. That’s two muscles now. And that is good; be you predator or prey. Let’s shake on it!
Who needs “Ice Road Truckers” when you have the “John’s Freezer” team on the road with fossils, amphibians, felids and 3D phenotype fun? No one, that’s who. We’re rocking the Cheltenham Science Festival for our first time (as a group), and pulling out all the stops by presenting two events! Here’s the skinny on them, with updates as the week proceeds.
Stomach-Churning Rating: 2/10 for now (just bones), but it could change once the cheetah dissection is under way… 8/10 bloody cheetah bits but only at the end (updated)
Right now, Lauren Sumner-Rooney (of “Anatomy To You” and other fame) is on-site with a rotating team of others from our lab, in the “Free Activity Tents” area of the Imperial Gardens/Square, inside a marquee where we’ll be showing off our NERC-funded tetrapod research all week. This “First Steps” event features not only our past and present work with Jenny Clack, Stephanie Pierce, Julia Molnar and others on Ichthyostega & its “fishapod” mates, but also our “scampering salamanders” research in Spain, Germany and England. I’ve blogged a lot about all that, and won’t repeat it here, but you can see a 3D-printed Ichthyostega skeleton, view the skeleton in a virtual reality 3D environment, see related specimens and engage in kid-friendly activities, and talk to our team about this and other related research.
Ichthyostega 3D printed backbone is born!
The central themes of that event are how bone structure relates to function and how we can use such information, along with experimental measurements and computer models of real salamanders, to reconstruct how extinct animals might have behaved as well as how swimming animals became walking ones. How did fins transform into limbs and what did that mean for how vertebrates made the evolutionary transition onto land? If you know my team’s work, that encapsulates our general approach to many other problems in evolutionary biomechanics (e.g. how did avian bipedalism evolve?). Added benefits are that you too can explore this theme in a hands-on way, and you can talk with us about it in person. That continues all week (i.e. until Saturday evening); I’ll be around from Thursday afternoon onwards, too. Kids of all ages are welcome!
Ichthyostega 3D print taking shape!
Then, on Saturday for our second free event we join forces with Ben Garrod (master of primate evolution, the secrets of bones, and “Attenborough and the Giant Dinosaur”) and RVC’s forensic pathologist Alexander Stoll as well as Sophie Regnault (“sesamoid street” PhD student w/me). As the “Large Animal Dissection” title hints, it’s not the right kind of gig to bring small kids to. There will be blood and stuff— we’ll be dissecting a cheetah together from 10am-4pm. This will involve walking through all the major organ systems, giving evolutionary anecdotes, and plenty more, with an aim to understand how the magnificent adaptations of cheetahs evolved—but also to investigate what problem(s) this animal faced that led to its sad demise. By the day’s end, there will just be a skeleton left. Get a front row seat early for this event, which serendipitously ties into “Team Cat”’s Leverhulme Trust-funded research project (we wanted a big animal and it just happened to be a cheetah; I had hoped for a giant croc or a shark or something but can’t complain!).
Ichthyostega 3D print is ready!
If you miss these events, please do cry bitter tears of regret. But don’t despair, there will be another “big cat dissection” in the London area in ~November (watch here for details), and plenty more fossil tetrapod stuff to come, plus a LOT more dinosaurs on the horizon!
Guess the bones! (photo by Zoe Self)
And please come back to this blog post for more pics and stories as the week carries on… For hashtag afficionados, you can follow the fun on Twitter etc. at #firststepsCSF16. What a world we live in!
Update 1: While you’re here, check out our Youtube playlists of tetrapod-related videos:
Update 2: Photos of our main stand (about tetrapod evolution)
Our poster/banner display looks nice.
Our tent brings in some punters.
Our bones excite people young and old, sighted and blind.
Fun with stickers and lab t-shirts.
And…
Update 3: Cheetah meat & greet
Ben, Alex, Sophie and I tackled the cheetah dissection today and it went GREAT! Much better than I’d optimistically expected. Rain didn’t scare the crowds off and neither did the gore, which there was some of (gelatinous spinal cords, lumpy tumors and at least one flying tiny bit of cheetah flesh that landed on a good-natured audience member!). Photos will tell the tale:
Peek-a-boo!
Sophie and Alex help us get set up in our tent.
Our initial rough schedule- although we ended up improvising more after lunch.
Dissectors assemble!
The beast revealed. It was skinned by the museum that loaned it to us.
Alex showing his talent: removing the viscera in one piece from end to end, starting with the tongue.
Impressive finding of a surgical fixture (plate and wires) on the tibia, which had been used to hold the shattered bone back together long enough for it to heal. Added to the kidney disease and liver-spleen-lung cancer, this cheetah was in the sorriest shape of any cadaver I’ve seen yet.
Cheetah coming to pieces: (from bottom) lumbar/pelvic region, hindlimb, thorax, forelimb and other bits.
Dr Adam Rutherford, an eye expert, did a nice dissection of the cheetah’s eye, here showing the tapetum lucidum (reflective membrane), which shows up as light blue colour. Its small size befits the not-very-nocturnal habits of cheetahs.
The lens of the cheetah’s eye. Now cloudy because of dehydration and crystalization, but still fascinating to see.
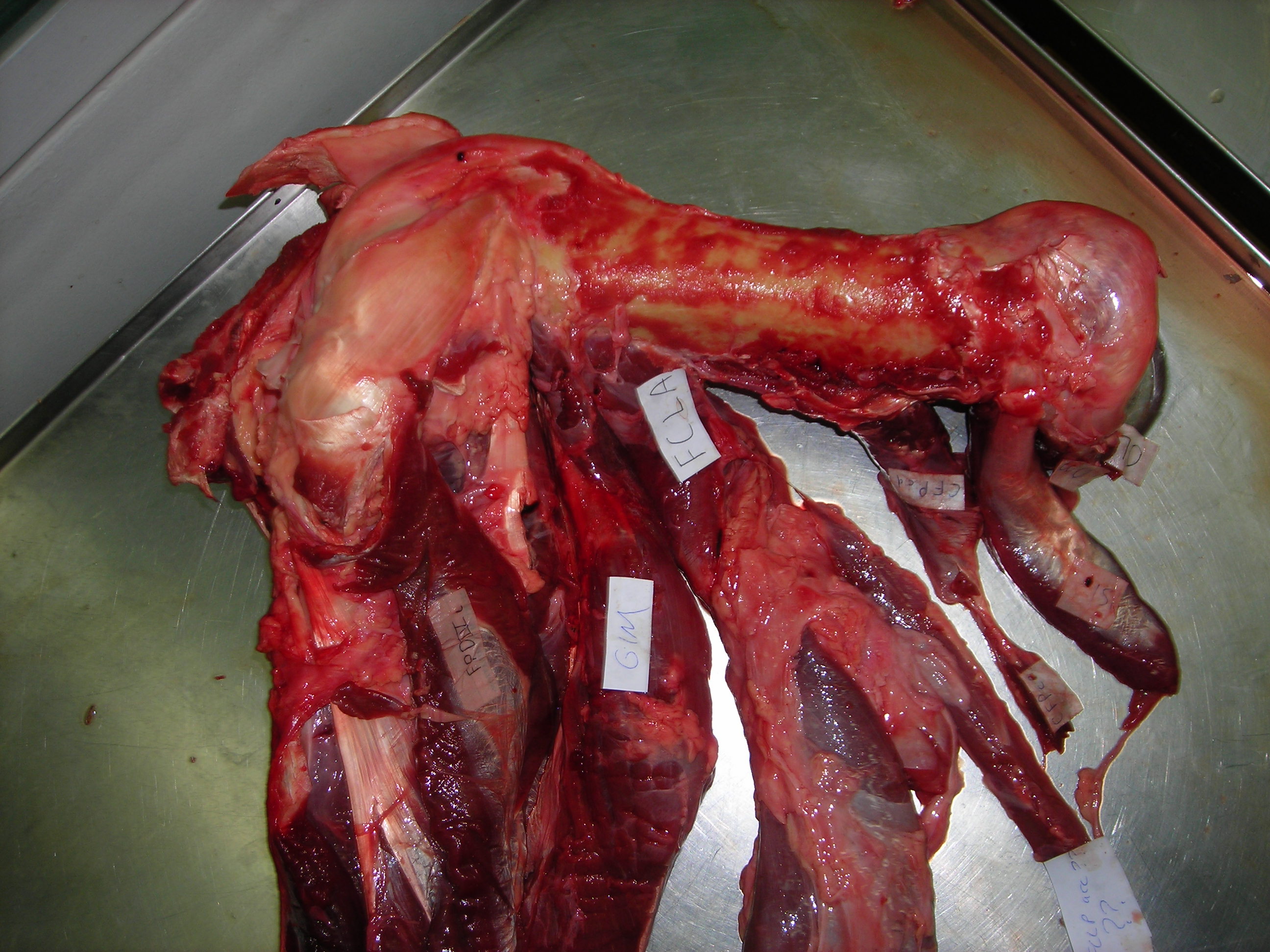
Happy Darwin Day from the frozen tundra sunny but muddy, frosty lands of England! I bring you limb muscles as peace offerings on this auspicious day. Lots of limb muscles. And a new theme for future blog posts to follow up on: starting off my “Better Know A Muscle” (nod to Stephen Colbert; alternative link) series. My BKAM series intends to walk through the evolutionary history of the coolest (skeletal/striated) muscles. Chuck Darwin would not enjoy the inevitable blood in this photo-tour, but hopefully he’d like the evolution. Off we go, in search of better knowledge via an evolutionary perspective!
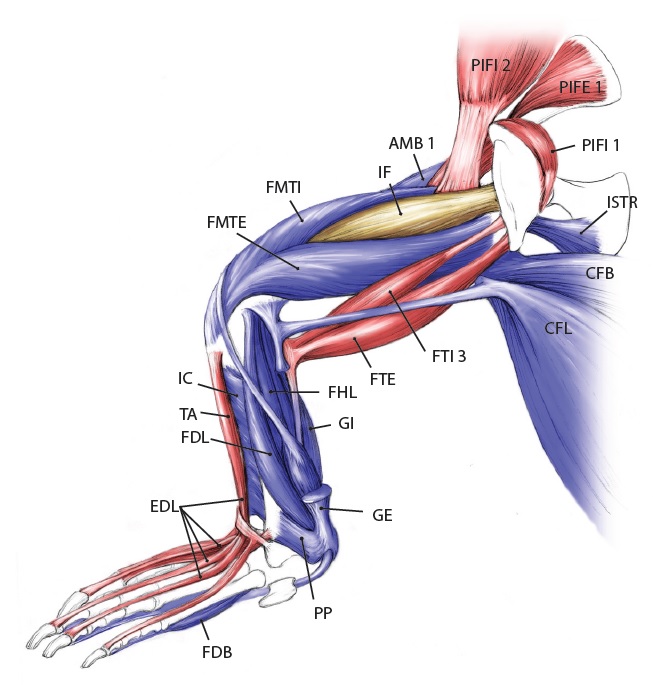
There is, inarguably, no cooler muscle than M. caudofemoralis longus, or CFL for short. It includes the largest limb muscles of any land animal, and it’s a strange muscle that confused anatomists for many years– was it a muscle of the body (an axial or “extrinsic” limb muscle, directly related to the segmented vertebral column) or of the limbs (an “abaxial” muscle, developing with the other limb muscles from specific regions of the paraxial mesoderm/myotome, not branching off from the axial muscles)? Developmental biologists and anatomists answered that conclusively over the past century: the CFL is a limb muscle, not some muscle that lost its way from the vertebral column and ended up stranded on the hindlimb.
The CFL is also a muscle that we know a fair amount about in terms of its fossil record and function, as you may know if you’re a dinosaur fan, and as I will quickly review later. We know enough about it that we can even dare to speculate if organisms on other planets would have it. Well, sort of…
Stomach-Churning Rating: 8/10. Lots of meaty, bloody, gooey goodness, on and on, for numerous species. This is an anatomy post for those with an appetite for raw morphology.
Let’s start from a strong (and non-gooey) vantage point, to which we shall return. The CFL in crocodiles and most other groups is (and long was) a large muscle extending from much of the front half or so of the tail to the back of the femur (thigh bone), as shown here:
Julia Molnar’s fabulous illustration of Alligator‘s limb muscles, from our 2014 paper in Journal of Anatomy. Note the CFL in blue at the bottom right.
As the drawing shows, the CFL has a friend: the CFB. The CFB is a shorter, stumpier version of the CFL restricted to the tail’s base, near the hip. The “B” in its name means “brevis”, or runty. It gets much less respect than its friend the CFL. Pity the poor CFB.
But look closer at the CFL in the drawing above and you’ll see a thin blue tendon extending past the knee to the outer side of the lower leg. This is the famed(?) “tendon of Sutton“, or secondary tendon of the CFL. So the CFL has two insertions, one on the femur and one (indirectly) onto the shank. More about that later.
Together, we can talk about these two muscles (CFL and CFB) as the caudofemoralis (CF) group, and the name is nice because it describes how they run from the tail (“caudo”) to the femur (“femoralis”). Mammal anatomists were late to this party and gave mammal muscles stupidly unhelpful names like “gluteus” or “vastus” or “babalooey”. Thanks.
But enough abstract drawings, even if they rock, and enough nomenclature. Here is the whopping big CFL muscle of a real crocodile:
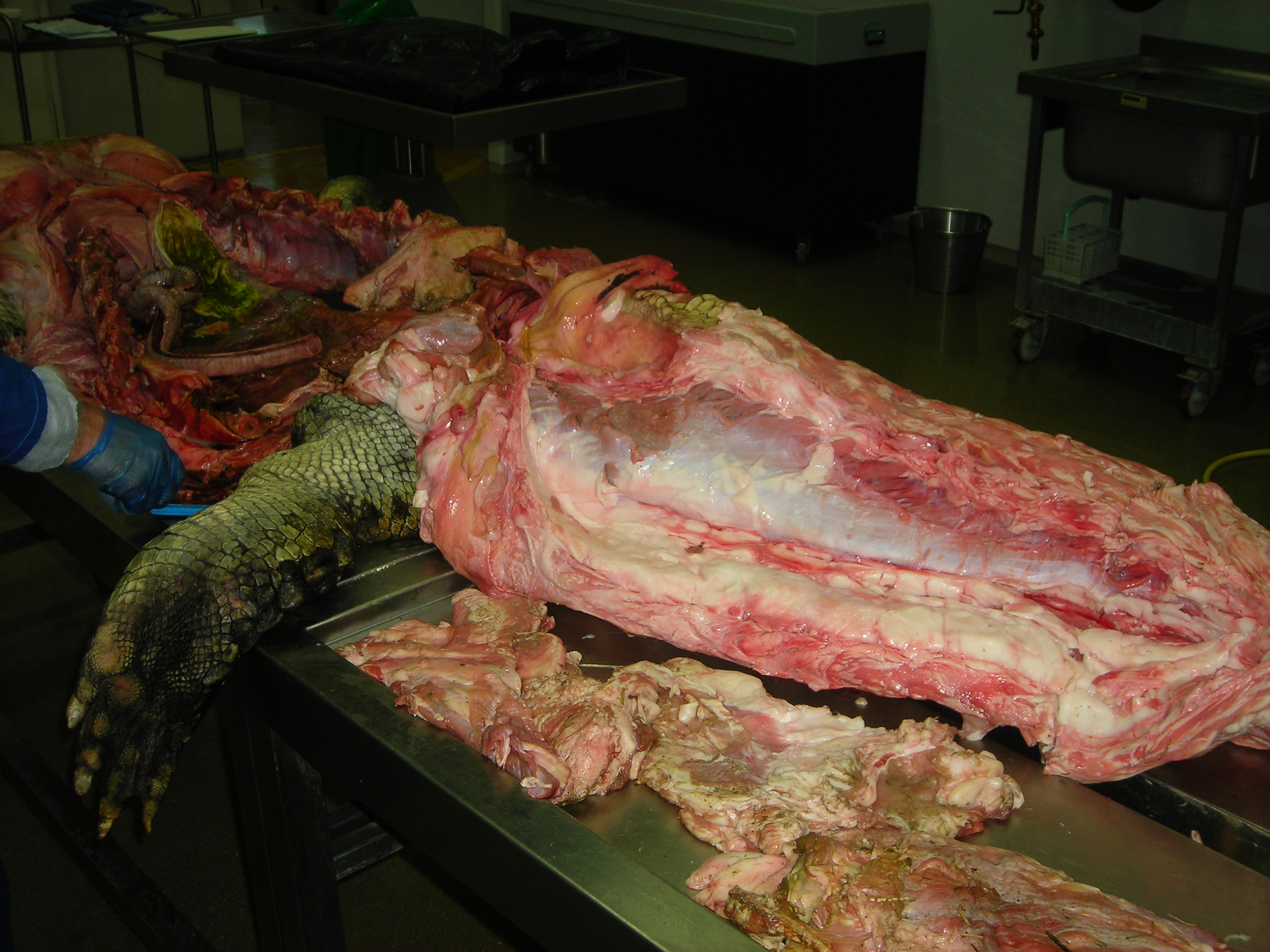
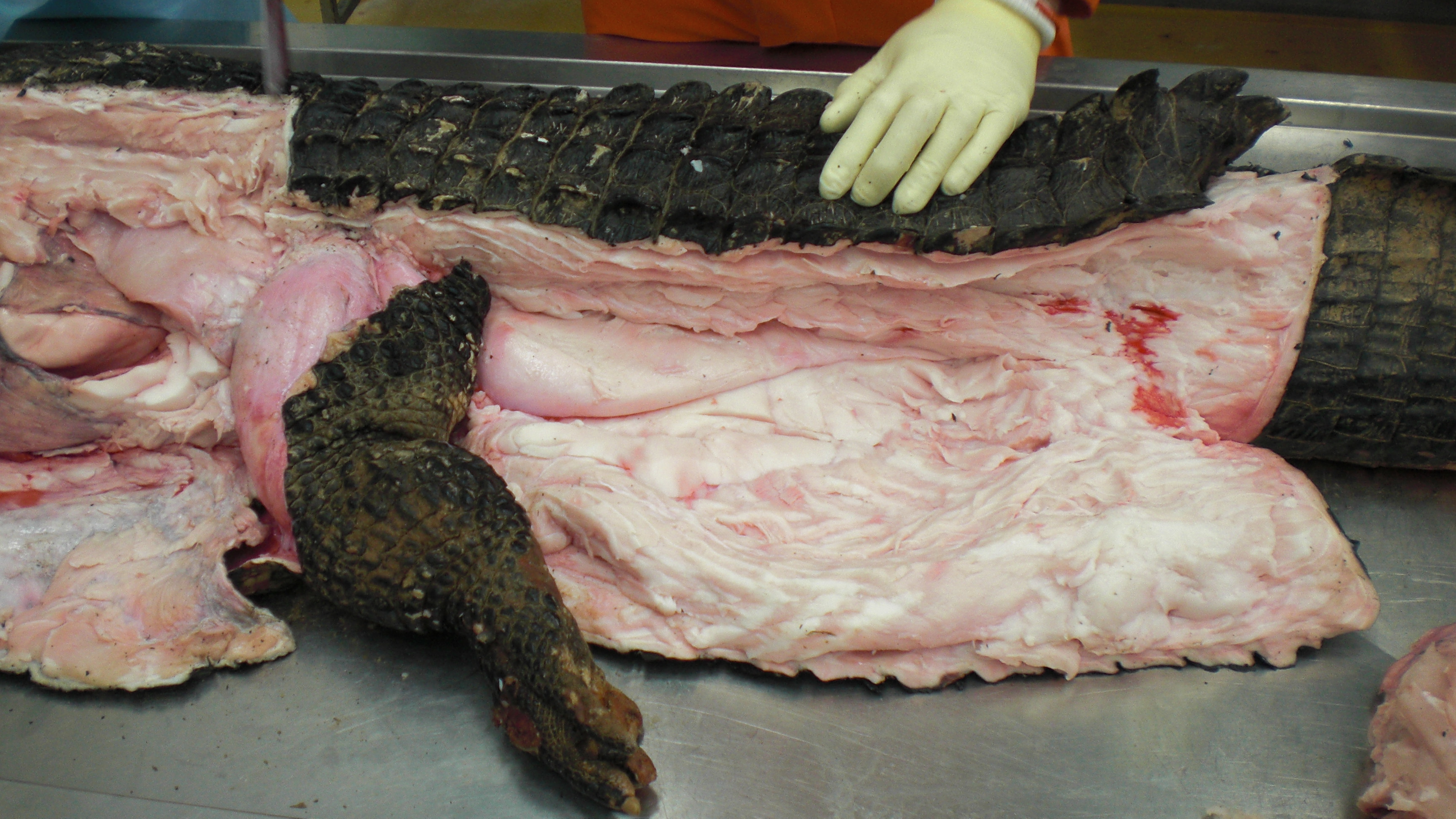
Huge Nile crocodile, but a relatively small CFL.
Bigger crocs have smaller legs and thus smaller leg muscles, relatively speaking. CFL at the top, curving to the left.
The giant Nile croc’s CFL muscle removed for measurements. 2.35 kg of muscle! Not shabby for a 278 kg animal.
However, maybe crocodile and other archosaur CFL muscles are not “average” for leggy vertebrates? We can’t tell unless we take an evolutionary tack to the question.
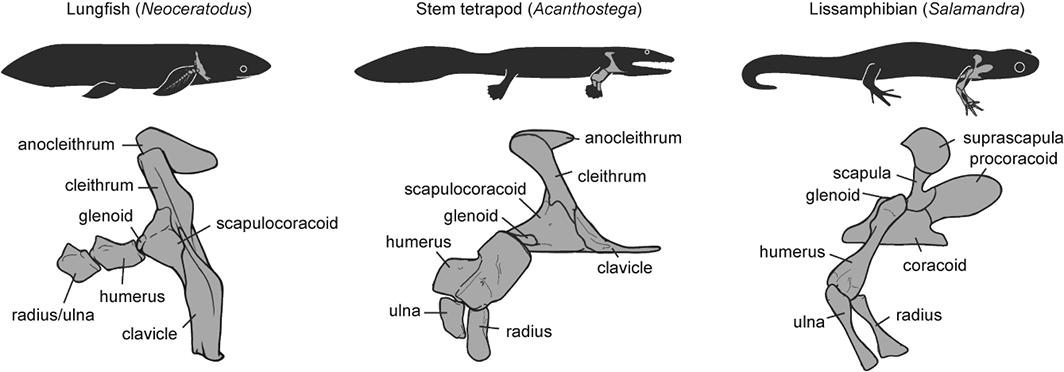
Where did the CFL come from, you may ask? Ahh, that is shrouded in the fin-limb transition‘s mysteries. Living amphibians such as salamanders have at least one CF muscle, so a clear predecessor to the CFL (and maybe CFB) was present before reptiles scampered onto the scene.
But going further back through the CF muscles’ history, into lobe-finned fish, becomes very hard because those fish (today) have so few fin muscles that, in our distant fishy ancestors, would have given rise eventually to the CF and other muscle groups. With many land animals having 30+ hindlimb muscles, and fish having 2-8 or so, there obviously was an increase in the number of muscles as limbs evolved from fins. And because a limb has to do lots of difficult three-dimensional things on land while coping with gravity, more muscles to enable that complex control surely were needed.
OK, so there were CF muscles early in tetrapod history, presumably, anchored on that big, round fleshy tail that they evolved from their thin, finned fishy one — but what happened next? Lizards give us some clues, and their CFL muscles aren’t all that different from crocodiles, so the CFL’s massive size and secondary “tendon of Sutton” seems to be a reptile thing, at least.
Courtesy of Emma Schachner, a large varanid lizard’s very freshly preserved CFL and other hindlimb muscles.
Courtesy of Emma Schachner, zoomed in on the tendons and insertions of the CFL muscle and others. Beautiful anatomy there!
Looking up at the belly of a basilisk lizard and its dissected right leg, with the end of the CFL labelled. It’s not ideally dissected here, but it is present.
An unspecified iguanid(?) lizard, probably a juvenile Iguana iguana, dissected to reveal its CFL muscle near its attachment to the femur. The muscle would extend further, about halfway down the tail, though.
Let’s return to crocodiles, for one because they are so flippin’ cool, and for another because they give a segue into archosaurs, especially dinosaurs, and thence birds:
A moderate-sized (45kg) Nile crocodile with its CFL muscle proudly displayed. Note the healthy sheath of fat (cut here) around the CFL.
American alligator’s CFL dominates the photo [by Vivian Allen].
Black caiman, Melanosuchus, showing off its CFL muscle (pink “steak” in the middle of the tail near the leg), underneath all that dark armour and fatty superficial musculature.
A closer look at the black caiman’s thigh and CFL muscle.
Like I hinted above, crocodiles (and the anatomy of the CFL they share with lizards and some other tetrapods) open a window into the evolution of unusual tail-to-thigh muscles and locomotor behaviours in tetrapod vertebrates.
Thanks in large part to Steve Gatesy’s groundbreaking work in the 1990s on the CFL muscle, we understand now how it works in living reptiles like crocodiles. It mainly serves to retract the femur (extend the hip joint), drawing the leg backwards. This also helps support the weight of the animal while the foot is on the ground, and power the animal forwards. So we call the CFL a “stance phase muscle”, referring to how it mainly plays a role during ground contact and resisting gravity, rather than swinging the leg forwards (protracting the limb; i.e. as a “swing phase muscle”).
The “tendon of Sutton” probably helps to begin retracting the shank once the thigh has moved forward enough, facilitating the switch from stance to swing phase, but someone really needs to study that question more someday.
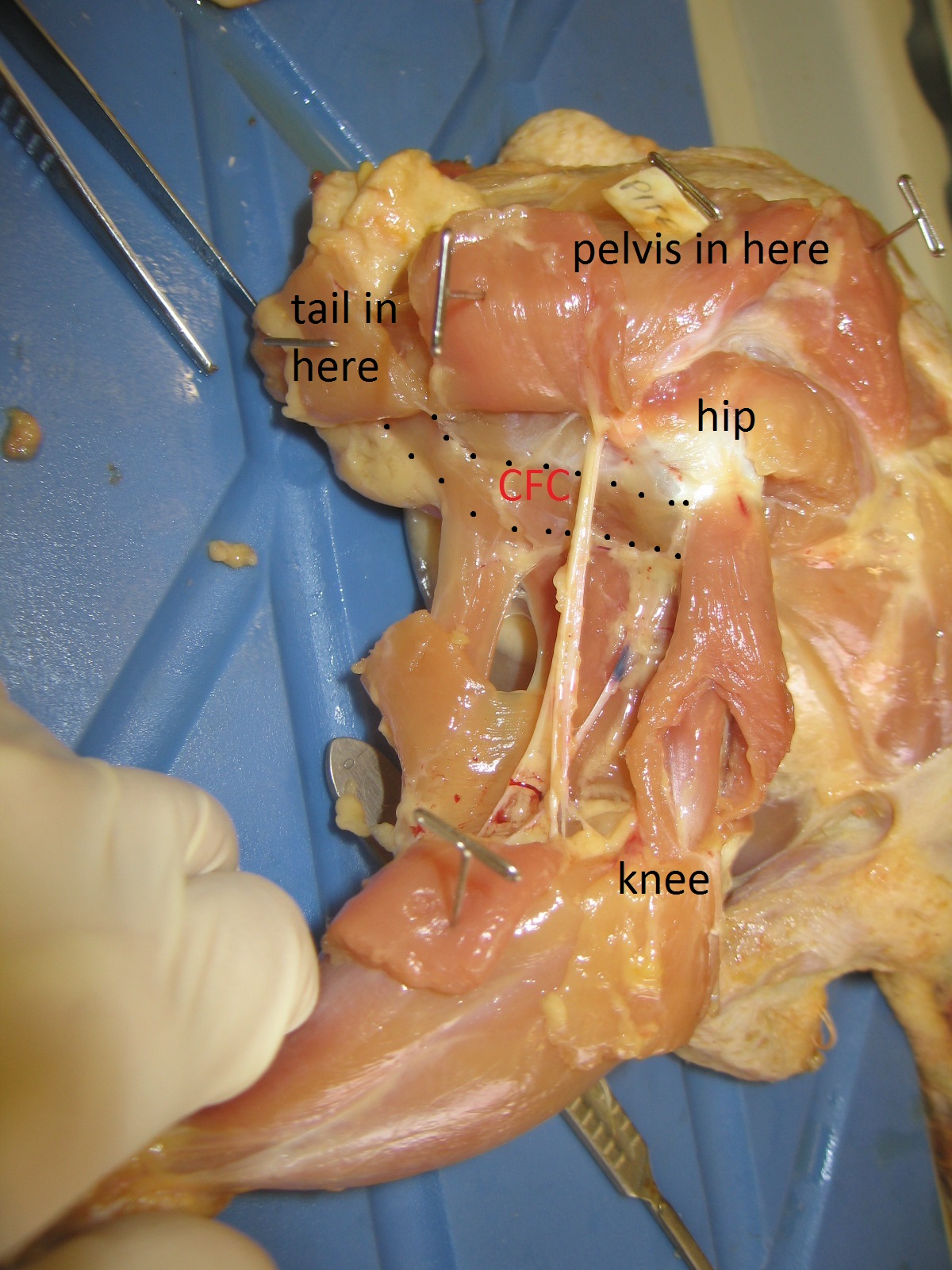
And thanks again to that same body of work by Gatesy (and some others too), we also understand how the CFL’s anatomy relates to the underlying anatomy of the skeleton. There is a large space for the CFL to originate from on the bottom of the tail vertebrae, and a honking big crest (the fourth trochanter) on the femur in most reptiles that serves as the major attachment point, from which the thin “tendon of Sutton” extends down past the knee.
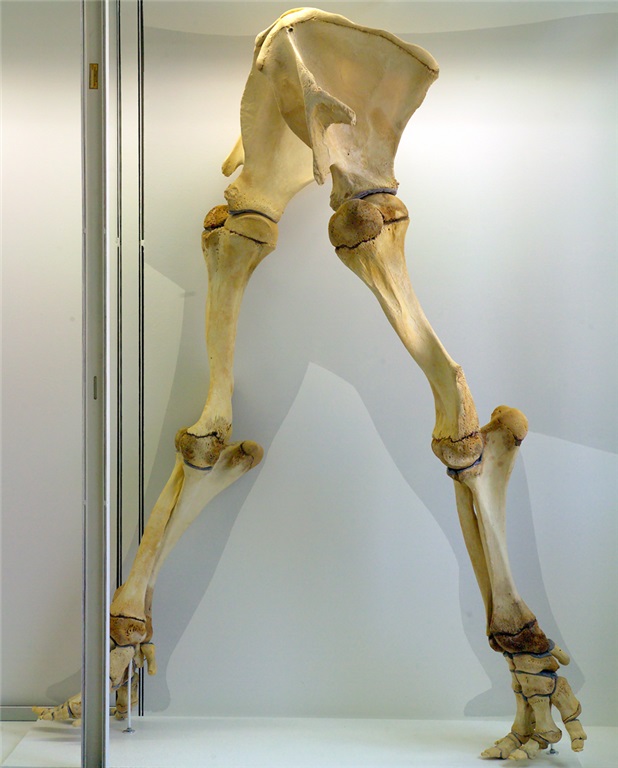
Femur bones (left side; rear view) from an adult ostrich (left) and Nile crocodile (right). Appropriate scale bar is appropriate. The fourth trochanter for the CFL is visible in the crocodile almost midway down the femur. Little is left of it in the ostrich but there is a bumpy little muscle scar in almost the same region as the fourth trochanter, and this is where the same muscle (often called the CFC; but it is basically just a small CFL) attaches.
That relationship of the CFL’s muscular anatomy and the underlying skeleton’s anatomy helps us a lot! Now we can begin to look at extinct relatives of crocodiles; members of the archosaur group that includes dinosaurs (which today we consider to include birds, too), and things get even more interesting! The “tendon of Sutton”, hinted at by a “pendant” part of the fourth trochanter that points down toward the knee, seems to go away multiple times within dinosaurs. Bye bye! Then plenty more happens:
A large duckbill dinosaur’s left leg, with a red line drawn in showing roughly where the CFL would be running, to end up at the fourth trochanter. Many Mesozoic dinosaurs have skeletal anatomy that indicates a similar CFL muscle.
We can even go so far as to reconstruct the 3D anatomy of the CFL in a dinosaur such as T. rex (“Sue” specimen here; from Julia Molnar’s awesome illustration as part of our 2011 paper), with a fair degree of confidence. >180kg steak, anyone?
As we approach birds along the dinosaur lineage, the tail gets smaller and so does the fourth trochanter and thus so must the CFL muscle, until we’re left with just a little flap of muscle, at best. In concert, the hindlimbs get more crouched, the forelimbs get larger, flight evolves and voila! An explosion of modern bird species!
Left femur of an ostrich in side view (hip is toward the right side) showing many muscles that attach around the knee (on the left), then the thin strap of CF muscle (barely visible; 2nd from the right) clinging near the midshaft of the femur.
Another adult ostrich’s CF muscle complex, removed for study. Not enough ostrich myology for you yet? Plenty more in this old post!Or this one!Or this one… hey maybe I need to write less about ostriches? The CF muscle complex looks beefy but it’s no bigger than any other of the main hindlimb muscles, unlike the CFL in a crocodile or lizard, which puts everything else to shame!
STILL not enough ostrich for you yet? Take a tour of the major hindlimb muscles in this video:
And check out the limited mobility of the hip joint/femur here. No need for much femur motion when you’re not using your hip muscles as much to drive you forwards:
But I must move on… to the remainder of avian diversity! In just a few photos… Although the CF muscles are lost in numerous bird species, they tend to hang around and just remain a long, thin, unprepossessing muscle:
Chicken’s right leg in side view. CFC muscle (equivalent of CFL; the ancestral CFB is confusingly called the CFP in birds, as it entirely resides on the pelvis) outlined and labelled.
A jay (species? I forget) dissected to show some of the major leg muscles, including the CFL-equivalent muscle; again, smallish. [Photo by Vivian Allen]
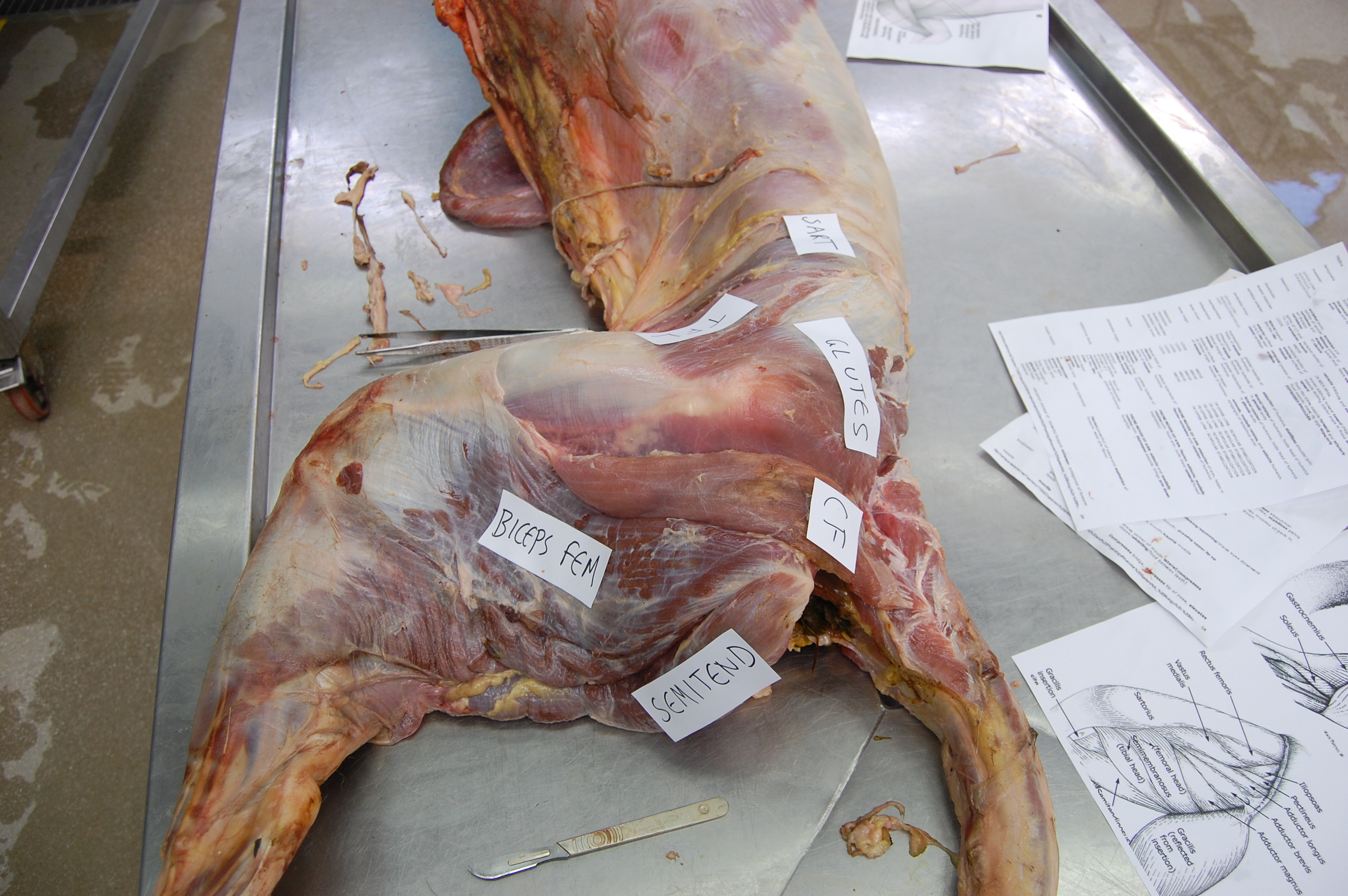
Finally, what’s up with mammals‘ tail-to-thigh CF-y muscles? Not much. Again, as in birds: smaller tail and/or femur, smaller CF muscles. Mammals instead depend more on their hamstring and gluteal muscles to support and propel themselves forward.
But many mammals do still have something that is either called the M. caudofemoralis or is likely the same thing, albeit almost always fairly modest in size. This evolutionary reduction of the CF muscle along the mammal (synapsid) lineage hasn’t gotten nearly as much attention as that given to the dinosaur/bird lineage’s CFL. Somebody should give it a thoroughly modern phylogenetic what-for! Science the shit outta that caudofemoralis…
Yet, oddly, to give one apparent counter-example, cats (felids) have, probably secondarily, beefed up their CF muscle a bit:
Cats have a pretty large CF muscle in general, and this jaguar is no exception! But mammals still tend to have fairly wimpy tails and thus CF muscles, or they even lose them (e.g. us?). [photo by Andrew Cuff, I think]
In summary, here’s what happened (click to embeefen):
Better Know A Muscle: the evolution of M. caudofemoralis (longus).
I hope you enjoyed the first BKAM episode!
I am willing to hear requests for future ones… M. pectoralis (major/profundus) is a serious contender.
P.S. It was Freezermas this week! I forgot to mention that. But this post counts as my Freezermas post for 2016; it’s all I can manage. Old Freezermas posts are here.
Ambitious experimental and morphological studies of a modern fish show how a flexible phenotype may have helped early “fishapods” to make the long transition from finned aquatic animals into tetrapods able to walk on land.
Stomach-Churning Rating: 1/10. Cute fish. Good science. Happy stomachs!
Photo by Antoine Morin, showing Polypterus on land.
Napoleon Bonaparte’s military excursions into Egypt in 1798-1799 led a young French naturalist, Ètienne Geoffroy Saint-Hilaire, to cross paths with a strange fish that had paired lungs and could “walk” across land on its stubby, lobelike fins. In 1802, he dubbed this fish “Polyptère bichir”1, today known as the Nile bichir, Polypterus bichir La Cepède 1803. The bichir’s mélange of primitive and advanced traits helped to catapult Geoffroy into scholarly conflict with the reigning naturalist Georges Cuvier back in France and to establish Ètienne as a leading anatomist, embryologist and early evolutionary researcher of repute even today2. Now, on their own excursion under the very “evo-devo” flag that the discoverer of Polypterus helped raise, Canadian scientists Standen et al.3 suggest how the remarkable plasticity of the skeleton of Polypterus (the smaller west African relative of P. bichir, P. senegalus or “Cuvier’s bichir”) reveals a key part of the mechanism that might have facilitated the gradual transition from water to land and thus from “fishapods” to tetrapods (four-limbed vertebrates).
In a bold experiment, the authors raised 149 young bichirs on land and in water for eight months, then studied how they moved on land vs. in water, and also how the ultimate shape of the skeletal elements of the paired front fin bases differed between the land- and water-raised bichirs. Standen et al.3 discovered that both the form and function of the fins’ foundations transformed to better satisfy the constraints of moving on land. Land-acclimated bichirs took faster steps on land, their fins slipped across the substrate less, they held their fins closer to their body, their noses stayed more aloft and their tails undulated less, with less variable motions overall—behaviours that the authors had predicted should appear to enhance walking abilities on land. In turn, the bones of the neck and shoulder region altered their shape to produce a more mobile fin base with greater independence of fin from neck motion, along with improved bracing of the ventral “collarbone” region. These environmentally-induced traits should have fostered the locomotor changes observed in “terrestrialized” fish and aided the animals in resisting gravity, and they represent a common biological phenomenon termed developmental plasticity4,5. Interestingly, the land-reared fish could still swim about as well as the wholly aquatic cohort, so there was not a clear trade-off between being a good swimmer and a good walker, which is surprising.
Considered alone, the developmental plasticity of bichir form and function shows how impressive these amphibious fish are. But Standen et al.’s study3 ventured further, to apply the lessons learned from bichir ontogeny to a phylogenetic context and macroevolutionary question. The phenotypic plasticity during bichir development, they infer, could have been harnessed during the evolutionary transformation of fins for swimming into limbs for walking, in the “fishapod” ancestors of tetrapods. Indeed, bichirs are close to the base of the family tree of fishes6, and other living relatives of tetrapods have reduced or lost their fins (lungfishes) or adapted to strange deep-sea swimming lifestyles, never walking on land (coelacanths). Thus perhaps bichirs and the “fishapod” lineage share what Geoffroy would have called “unity of type”, today termed homology, of their developmental plasticity in response to a land environment. Surveying the fossil record of early “fishapods” and tetrapods, Standen et al.3 found that the macroevolutionary changes of neck and shoulder anatomy in these gradually more land-adapted animals parallel those they observed in terrestrialized Polypterus, providing ancillary support for their hypothesis.
A further test of the application of Polypterus’s plasticity to fossil tetrapods is naturally difficult. However, the “fishapod” lineage has some exceptional examples of fossil preservation. With sufficient sample sizes (e.g. fossil beds that reveal growth series, such as the Late Devonian Miguasha site in Canada7) and palaeoenvironmental gradients in fish or tetrapods, one could imagine performing a rigorous indirect test. Even small samples could be helpful– for example, the early tetrapod Ichthyostega exhibits some developmental changes in its forelimb suggesting that it became more terrestrial as it grew, whereas the related Acanthostega does not evidence such changes8— this hints at some developmental plasticity in the former animal.
During the Devonian period (~360-420 million years ago), were the “fishapod” ancestors of tetrapods floundering about on land now and then, gradually shifting from anatomy and behaviours that were more developmentally plastic (as in bichirs) to ones that were more canalized into the terrestrialized forms and functions that more land-adapted tetrapods retained? An attractive possibility is that the developmental plasticity could have led to fixation (reduction of plasticity), an evolutionary phenomenon called genetic assimilation, which another intellectual descendant of Geoffroy, Conrad Hal Waddington, promoted from the 1950s onwards9, a concept that now enjoys numerous cases of empirical support10 that this one may eventually join.
The nature of the genetic and developmental mechanism that bichirs use to achieve the observed developmental plasticity is still unclear. If it has a high enough degree of heritability, then it could be selected for in cross-generational experiments with bichirs. With sufficient time and luck raising these unusual fish, the hypothesis that their plastic response to a terrestrial environment can become genetically assimilated could be directly tested. This study could thus become an epic exemplar of how genetic assimilation can contribute not only to microevolutionary change but also to major macroevolutionary events, as was presciently suggested in a seminal review of developmental plasticity4.
This genetic assimilation is the Polypterus study’s reasonable speculation, and one that Geoffroy likely would have applauded, all the more for involving his beloved bichirs. Much as Napoleon’s landfall in Egypt was not a lasting success, bichirs never left wholly terrestrial descendants despite their malleable locomotor system. But the same type of plastic developmental mechanism that bichirs use today to make tentative, floppy incursions of the terrestrial realm might have been harnessed by our own “fishapod” forebears, leaving a far more revolutionary dynasty upon the Earth.
References
Geoffroy, E. (1802). Histoire naturelle et description anatomique d’un nouveau genre de poisson du Nil, nommé polyptère. Annales du Muséum d’Histoire Naturelle 1:57-68.
Le Guyader, H., & Grene, M. (2004) Geoffroy Saint-Hilaire: A Visionary Naturalist. Univ. Chicago Press.
Standen, E. M., Du, T. Y., & Larsson, H. C. E. (2014). Developmental plasticity and the origin of tetrapods. Nature, published online.
West-Eberhard, M. J. (1989). Phenotypic plasticity and the origins of diversity. Annual Review of Ecology and Systematics 20:249-278.
Pigliucci, M., Murren, C. J., & Schlichting, C. D. (2006). Phenotypic plasticity and evolution by genetic assimilation. Journal of Experimental Biology 209(12):2362-2367.
Near, T. J., Dornburg, A., Tokita, M., Suzuki, D., Brandley, M. C., & Friedman, M. (2014). Boom and bust: ancient and recent diversification in bichirs (Polypteridae: Actinopterygii), a relictual lineage of ray‐finned fishes. Evolution68:1014-1026.
Cloutier, R. (2013). Great Canadian Lagerstätten 4. The Devonian Miguasha Biota (Québec): UNESCO World Heritage Site and a Time Capsule in the Early History of Vertebrates.Geoscience Canada40:149-163.
Callier, V., Clack, J. A., & Ahlberg, P. E. (2009). Contrasting developmental trajectories in the earliest known tetrapod forelimbs.Science324:364-367.
Waddington, C. H. (1953). Genetic assimilation of an acquired character. Evolution 7:118-126.
Crispo, E. (2007). The Baldwin effect and genetic assimilation: revisiting two mechanisms of evolutionary change mediated by phenotypic plasticity. Evolution 61:2469-2479.
Construction of the Phyletisches Museum in Jena, Germany began on Goethe’s birthday on August 28, 1907. The Art Nouveau-styled museum was devised by the great evolutionary biologist, embryologist and artist/howthefuckdoyousummarizehowcoolhewas Ernst Haeckel, who by that time had earned fame in many areas of research (and art), including coining the terms ontogeny (the pattern of development of an organism during its lifetime) and phylogeny (the pattern of evolution of lineages of organisms through time) which feature prominently in the building’s design and exhibits (notice them intertwined in the tree motif below, on the front of the museum). Ontogeny and phylogeny, and the flamboyant artistic sensibility that Haeckel’s work exuded, persist as themes in the museum exhibits themselves. Haeckel also came up with other popular words such as Darwinism and ecology, stem cell, and so on… yeah the dude kept busy.
Cavorting frogs from Haeckel’s masterpiece Kunstformen der Natur (1904).
I first visited the Phyletisches Museum about 10 years ago, then again this August. Here are the sights from my latest visit: a whirlwind ~20 minute tour of the museum before we had to drive off to far-flung Wetzlar. All images are click-tastic for embiggenness.
Stomach-Churning Rating: 3/10 for some preserved specimens. And art nouveau.
Willkommen!
Frog ontogeny, illustrated with gorgeous handmade ?resin? models.
Phylogeny of Deuterostomia (various wormy things, echinoderms, fish and us), illustrated with lovely artistry.
Phylogeny of fish and tetrapods.
Slice of fossil fish diversity.
Plenty of chondrichthyan jaws/chondrocrania, teeth and bodies.
Awesome model of a Gulper Eel — or, evocatively, “Sackmaul” auf Deutsch (Saccopharyngiformes).
Lobe-finned fishes (Sarcopterygii)- great assortment including a fossil coelacanth.
Lungfish body and skeleton.
Coelacanth!
Coelacanth staredown!
On into tetrapods– a Fire Salamander (Salamandra salamandra)! We love ’em, and the museum had several on display- given that we were studying them with x-rays, seeing the skeleton and body together here in this nice display was a pleasant surprise.
A tortoise shell and skeleton, with a goofball inspecting it.
In a subtle nod to recurrent themes in evolution, the streamlined bodies of an ichthyosaur and cetacean shown in the main stairwell of the museum, illustrating convergent evolution to swimming adaptations.
Phylogeny of reptiles, including archosaurs (crocs+birds).
Gnarly model of an Archaeopteryx looks over a cast of the Berlin specimen, and a fellow archosaur (crocodile). The only extinct dinosaur on exhibit!
Kiwi considers the differences in modern bird palates: palaeognathous like it and fellow ratites/tinamous (left), and neognathous like most living birds.
Echidna skeleton. I can’t get enough of these!
Skulls of dugong (above) and manatee (below), Sirenia (seacows) closely related to elephants.
Hyraxes, which Prof. Martin Fischer, longtime curator of the Phyletisches Museum, has studied for many years. Rodent-like elephant cousins.
Old exhibit at the Phyletisches Museum, now gone: Forelimbs of an elephant posed in the same postures actually measured in African elephants, for the instant of foot touchdown (left pic) and liftoff (right pic). Involving data that we published in 2008!
Eek, primates! Gorilla see, gorilla do. Notice the primitive “bent hip, bent knee” vs. the advanced “upright modern human” hindlimb postures in the two non-skeletal hominids.
Phylogeny of artiodactyl (even-toed) mammals, including the hippo-whale clade.
Hand (manus) of the early stem-whale Ambulocetus.
Carved shoulderblade (scapula) of a bowhead whale (Balaena mysticetus), which apparently Goethe owned (click to emwhalen and read the fine print). Quite a relic!
One of Haeckel’s residences, across the street from the museum. There is also a well-preserved house of his that one can visit, but I didn’t make it there. I heard it’s pretty cool.
Jena is tucked away in a valley in former East Germany, with no local airport for easy access- but get to Leipzig and take a 1.25 hour train ride and you’re there. Worth a trip! This is where not just ontogeny and phylogeny were “born”, but also morphology as a modern, rigorous discipline. Huge respect is due to Jena, and to Haeckel, whose quotable quotes and influential research still resonate today, in science as well as in art.
Short and sweet post here; it’s sunny outside and I want to be there BBQing!
I had a buried folder of CT files labelled as a species of fish, but on digging them out and segmenting them I realize it is not what I expected (inner fish or not!), as you will see.
Stomach-Churning Rating: 2/10; simple CT scan of a body.
Identify the animal in the CT scout/pilot image below, as specifically as you can. But… (READ THE SENTENCE BELOW FIRST BEFORE ANSWERING!)
Today’s special rule: Summertime is coming and that means superhero films! Your answer must be in the form of a dialogue between a superhero(ine) and a supervillain(ess)!
Difficulty: Even I am not 100% sure what this is but I have a decent idea. Not super hard, but not a super good segmentation.
Pow! Bam! Biff! Go forth and conquer! Then invite the Human Torch to your BBQ.