I had a spare hour in Cambridge this weekend so I dared the crowds in the revamped UMZC’s upper floor. In my prior visit and post I’d experienced and described the lower floor, which is almost exclusively mammals. This “new” floor has everything else that is zoological (animal/Metazoa) and again is organized in an evolutionary context. And here is my photo tour as promised!

Inviting, soft lighting perfuses the exhibits from the entryway onwards.
All images can be clicked to mu-zoom in on them.
Stomach-Churning Rating: 5/10 for spirit animals, by which I mean dissected/ghostly pale whole specimens of animals in preservative fluids.

The exhibits are on a square balcony overlooking the lower floor, so you can get some nice views. It does make the balcony crowded when the museum is busy, so take that in mind if visiting. Strollers on this upper floor could be really difficult. But the ceiling is very tall so it is not cramped in a 3D sense. The lower floor is more spacious.

Like phylogenies? You got em! Tucked away at the beginning of each major group; not occupying huge valuable space or glaringly obvious like AMNH in NYC but still noticeable and useful. To me, it strikes a good balance; gives the necessary evolutionary context for the displayed specimens/taxa.

Introductory panels explain how names are given to specimens, how specimens are preserved and more.

The exhibits give due focus to research that the UMZC is doing or has been famous for. Hey I recognize that 3D tetrapod image in the lower left! 🙂

There is ample coverage of diversity throughout Metazoa but my camera tended to be drawn to the Vertebrata. Except in some instances like these.

Some larger chelicerates.

Some smaller, shadowy sea scorpion (eurypterid) fossils.

Watch here for more about ophiuroids (brittlestars) in not too long!

A BIG fish brain! Interesting!
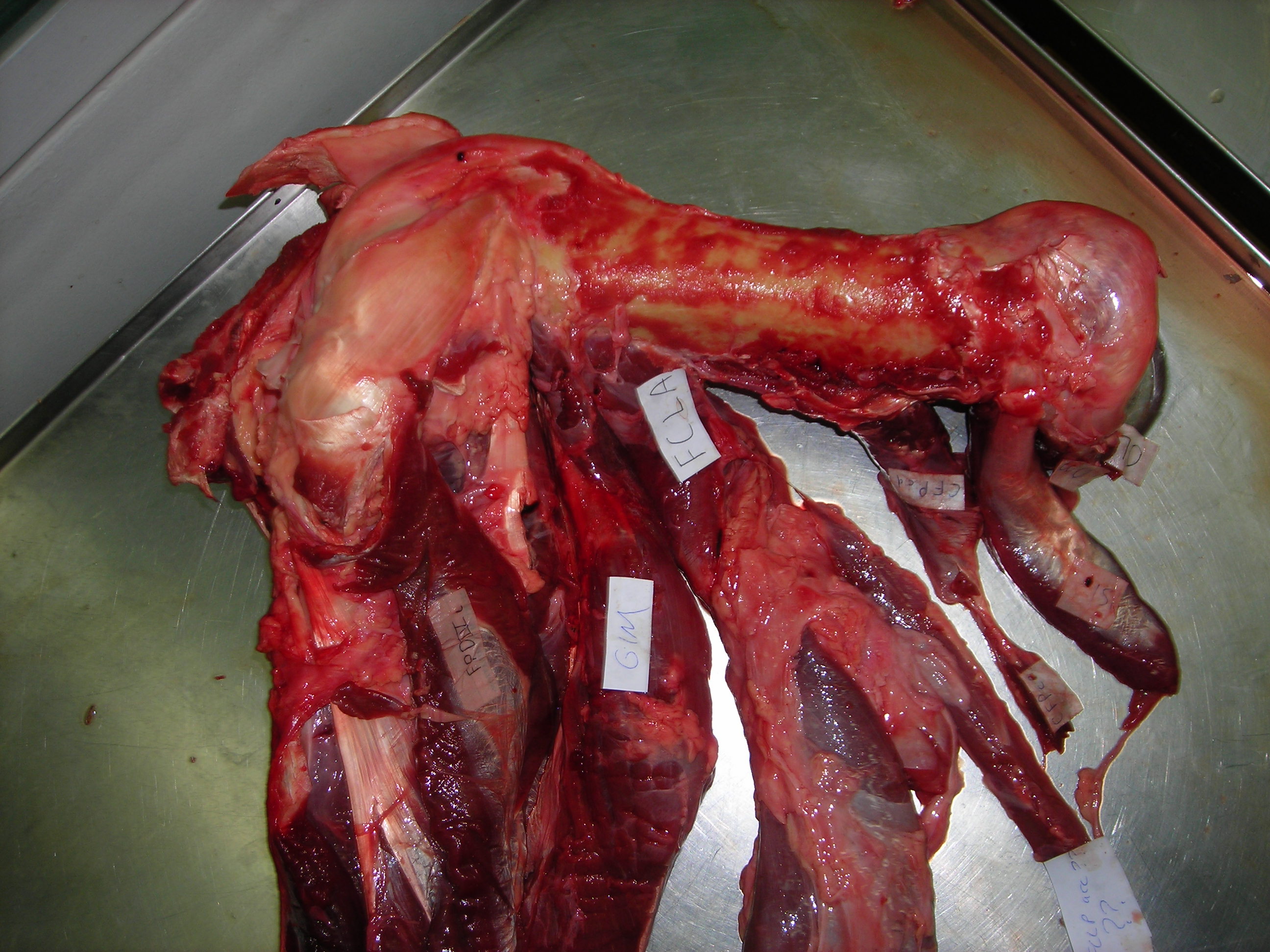
Before I go through specimens in evolutionary “sequence”, I will feature another thing i really liked: lots of dissected spirit-specimens that show off cool anatomy/evolution/adaptation (and technical skills in anatomical preparation). Mostly heads; mostly fish.

Salps and other tunicates! Our closest non-vertebrate relatives- and some insight into how our head and gut came to be.

Salp-reflection.

Lamprey head: not hard to spot the commonalities with the salps; but now into Vertebrata.

Hagfish head: as a fellow cyclostome/agnathan, much like a lamprey but never forget the slime glands!
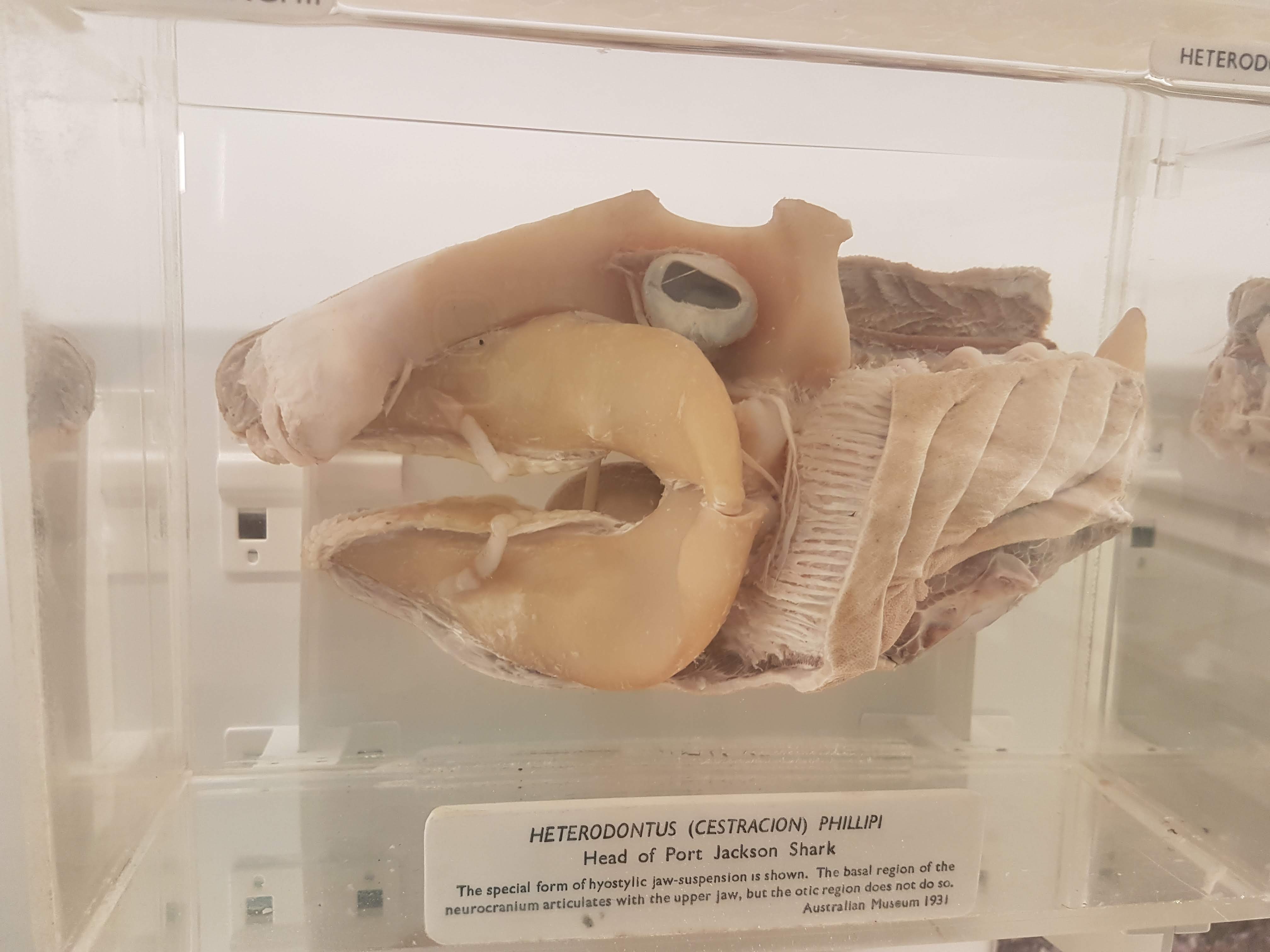
Shark head. Big fat jaws; all the better to bite prey with!

Lungfish (Protopterus) head showing the big crushing tooth plates (above).

Sturgeon vertebrae: tweak some agnathan/shark bits and here you are.

Carp head, with explainer on the Weberian apparatus linking the ear to the swim bladder.

Worm (annelid) anatomy model, displaying some differences from/similarities to Vertebrata. (e.g. ventral vs. dorsal nerve cord; segmentation)

Dissected flipper from a small whale/other cetacean. Still five fingers, but other specializations make it work underwater.

Wonderful diversity of tooth and jaw forms in sharks, rays and relatives. I like this display a lot.

More of the above, but disparate fossil forms!

On with the evolutionary context! Woven throughout the displays of modern animals are numerous fossils, like these lovely placoderms (lineage interposed between agnathans, sharks and other jawed fish).

Goblin shark head.

I seem to always forget what ray-finned fish this is (I want to say wolffish? Quick Googling suggests maybe I am right), but see it often and like its impressive bitey-ness.

Bichir and snakefish; early ray-finned fish radiations.

Armoured and similar fish today.

Armoured fish of the past; some convergent evolution within ray-fins.

Convergence- and homology- of amphibious nature in fish is another evolutionary pattern exemplified here.

Gorgeous fossils of ray-finned fish lineages that arose after the Permian extinctions, then went extinct later in the Triassic.

Note the loooooong snout on this cornetfish but the actual jaws are just at the tip.

Flying fish– those ray-fins are versatile.

Diversity of unusual ray-finned fish, including deep-water and bottom-dwelling forms.

Can you find the low-slung jaws of a dory?

Recent and fossil perch lineage fish.

It’s hard to get far into talking about evolution without bringing up the adaptive radiation of east African cichlid fish, and UMZC researchers are keen on this topic too.

Lobe-fins! Everybody dance!

Rhizodonts & kin: reasons to get out of Devonian-Carboniferous waters.

A Cretaceous fossil coelacanth (skull); not extremely different from living ones’.

Let’s admire some fossil and modern lungfish skulls, shall we? Big platey things (here, mainly looking at the palate) with lots of fusions of tiny bones on the skull roof.

Eusthenopteron fossils aren’t that uncommon but they are still great to see; and very important, because…

OK let’s stop messing around. The UMZC has one of the best displays of fossil stem-tetrapods in the world! And it should.

Another look at the pretty Acanthostega models.

Acanthostega vs. primate forelimb: so like us.

Ichthyostega parts keep Acanthostega company.

A closer look at the “Mr. Magic” Ichthyostega specimen, which takes some unpacking but is incredibly informative and was a mainstay of our 2012 model. Back of skull, left forelimb, and thorax (from left to right here).

Eucritta, another stem-tetrapod.

Closer look at Eucritta‘s skull.

Weird stem-tetrapod Crassigyrinus, which we’re still trying to figure out. It’s a fabulous specimen in terms of completeness, but messy “roadkill” with too many damn bones.

The large skull of Crassigyrinus, in right side view.

Early temnospondyl (true amphibian-line) skulls and neck.

Nectrideans or the boomerangs of the Palaeozoic.

Cool fossil frogs.

Giant Japanese salamander!

Fire salamanders: not as colourful as the real thing, but here revealing their reproductive cycle in beautiful detail.

Closeup of oviduct in above.

Sexual dimorphism in Leptodactylus frogs: the males have bulging upper arms to (I am assuming) help them hold onto females during amplexus (grasping in mating competitions).

Did I forget that Leptodactylus has big flanges on the humerus in males, to support those muscles? Seems so.

An early stem-amniote, Limnoscelis (close to mammals/reptiles divergence); cast.

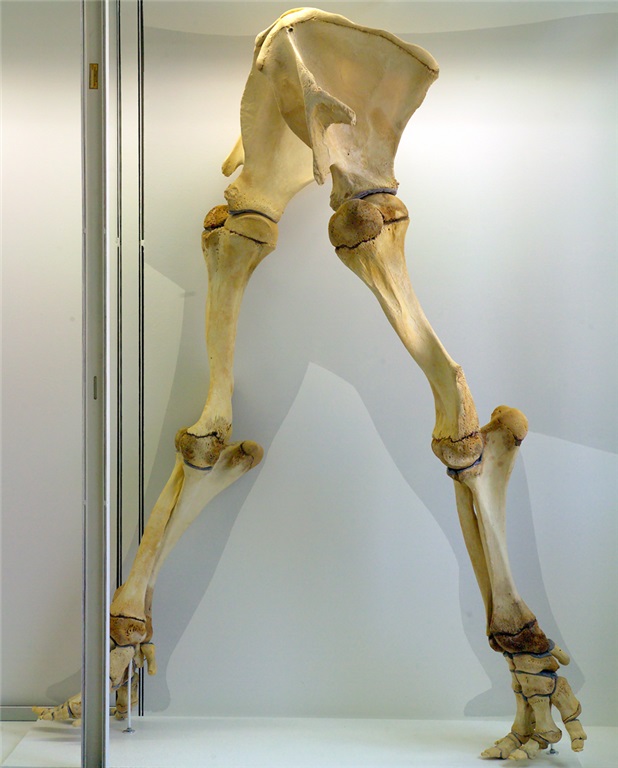
Grand sea turtle skeleton.

One of my faves on display: a real pareiasaurian reptile skeleton, and you can get a good 3D look around it.

Details on above pareiasaurian.

Mammals are downstairs, but we’re reminded that they fit into tetrapod/amniote evolution nonetheless.

Let there be reptiles! And it was good.

Herps so good. (slow worm, Gila monster, glass lizard)

A curator is Dr Jason Head so you bet Titanoboa is featured!

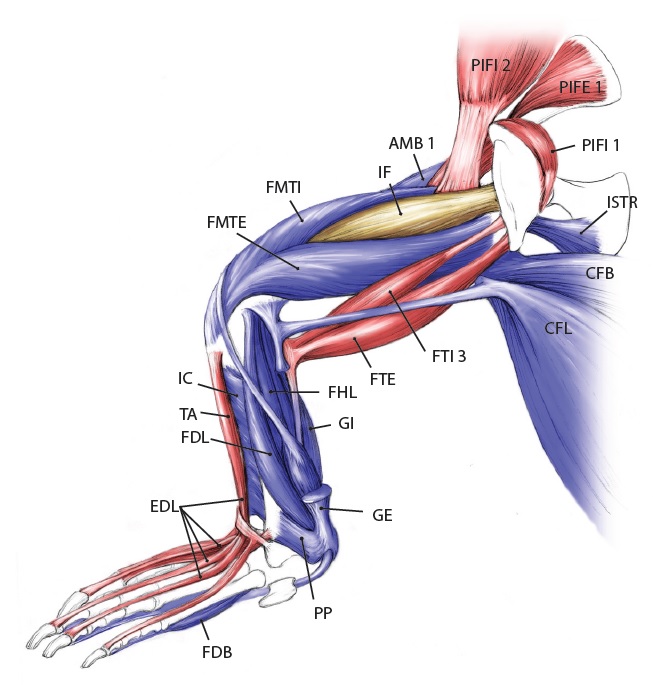
Crocodylia: impressive specimens chosen here.

It ain’t a museum without a statuesque ratite skeleton. (There are ~no non-avian dinosaurs here– for those, go to the Sedgwick Museum across the street, which has no shortage!)

Avian diversity takes off.

Glad to see a tinamou make an appearance. They get neglected too often in museums- uncommon and often seemingly unimpressive, but I’m a fan.

I still do not understand hoatzins; the “cuckoo” gone cuckoo.

Dodo parts (and Great Auk) near the entrance.

Wow. What an oilbird taxidermy display! :-O
There we have it. Phew! That’s a lot! And I left out a lot of inverts. This upper floor is stuffed with specimens; easier there because the specimens are smaller on average than on the lower floor. Little text-heavy signage is around. I give a thumbs-up to that– let people revel in the natural glory of what their eyes show them, and give them nuggets of info to leave them wanting more so they go find out.
Now it’s in your hands– go find out yourself how lovely this museum is! I’ve just given a taste.