The pandemic goes on; my life goes on; but it has been another rough 1+ years. I have hardly done any hands-on anatomy as I’m hardly on campus at all, and my team’s work has mostly shifted into digital modelling for now (more about that below; it is not a bad thing though). My main news for 2021-2022 falls into the categories of Life Stuff, and Work Stuff to summarise here. “WTF” sums it up as it has been a… strange time; very challenging at a personal level, due to the Life Stuff.
Stomach-Churning Rating: stuff is weird, but nothing truly stomach-churning is here
Life Stuff: It’s been about the same as 2020-2021; summarised here. Thankfully, no major grief from losing people/pets close to me, this time. But my heath has been really awful instead — my epilepsy returned in May 2021, much to our surprise, after 2 years of remission. I suspect dehydration was a cause, as I later found out that I’d been chronically dehydrated, which came as a shock. I’ve since learned to step up my hydration routine, and I feel better. Right now I’m >5 months seizure-free, after a very hard time of monthly seizures for ~5 months in a row, including a scary one after a flight from Phoenix-London, in which I woke up in a toilet stall at Heathrow baggage claim, very disoriented and alone, eerily with no one in that large men’s loo area. My taxi driver was wondering why I was so late… I am glad I didn’t fall and hurt myself with no one around. Fingers crossed that doesn’t happen again. But on top of that, I’ve been fighting a longstanding chronic illness (details are not necessary) at the same time, and that got very bad in April 2022, sending me to hospital with severe internal infection; very life-threatening, painful and frightening. Again, right now I feel that I’m in recovery, and grateful for some good (overall) care from the NHS. Owing to these health problems though, plus the pandemic and financial challenges, I’ve not been travelling and don’t foresee much of that for my near future. Which also means not enough real holiday; “staycations” in my house just aren’t enough, as I’ve been here for ~2.5 years. I’m starting to do more fun things, finally, again, and that led to this blog post (first one in almost one year). I feel I have some energy to do things that I enjoy again.
Work Stuff: Mostly that has been pretty good, with a caveat. The DAWNDINOS project still dominated my work life, much to my pleasure. Indeed, just this week I tied the final ribbon on that, formally, with submission of my final grant report to the EU/ERC. The grant ended on March 31, 2022 and I was VERY, VERY sad to have to bid farewell to my team, who I hugely enjoyed working with for those 5.5 years. Now comes the caveat to “work is good”, which is that suddenly I have no funding (feast-to-famine) and “just” one PhD student (Vittorio LaBarbera; reinforcement learning simulations of locomotion); MRes student Georgia Wells just finished; and a Research Fellow (Dr. Masaya Iijima). It looks like I’ll be doing more undergrad research projects than postgrad for awhile, but we’ll see. The grant funding lottery can be hard to predict. Regardless, there’s a lot of fun science going on! With DAWNDINOS, since last summary we’ve cranked out a bunch of cool papers on archosaur locomotor biomechanics — find them here. #25-31 are the newer ones I haven’t blogged about anywhere yet; #25, 28 and 30 are blogged about by Dr. Ashleigh Wiseman here; and #30 (which is, in part, a summary of DAWNDINOS to date) got SICB conference coverage here.

Muscle-bound Euparkeria hindlimbs from our DAWNDINOS paper #28; picture by Oliver Demuth.
DAWNDINOS paper #26 with DAWNDINOS postdoc Dr. Delyle Polet was a serendipitous one inspired by him giving a seminar to our lab when he first came to the UK, and it struck me that his method for using biomechanical simulations with the “Murphy number” (related to pitch moment of inertia; MOI) to test how animals move would work really well with a long-bodied, hefty Triassic pseudosuchian (= large pitch MOI) such as Batrachotomus, whose results we could compare with known fossil trackways of similar archosaurs (e.g., Isocheirotherium). We found evidence for it using at least two running gaits, which was pretty surprising.
And just this week we published another “spin-off” paper (also see van Beesel primate shoulder-modelling studies #17,#29) adapting our 3D digital modelling methods to another taxon. This one came out of left field for me (I’d never expected I’d work on sharks!) but actually fits very well with my research interests in giant animals, biomechanics and palaeontology. We reconstructed the giant shark Otodus megalodon from the best fossils available (including a Belgian vertebral column somewhat neglected since the 1860s), finding that it was ~16m long and >60,000 kg; but this is not the largest it could get, as a vertebra ~50% larger is known! This paper got a LOT of nice press attention, and the video below is perhaps the best science communication release I’ve been involved with (all kudos go to Catalina Pimiento and the animation team she commissioned). Very importantly, the key data are free to use.
LATE ADDITIONS: But it wasn’t all #DAWNDINOS-related research! I was very pleased to have Dr. Chris Basu’s PhD work with me on giraffid locomotor biomechanics published in PNAS. We showed, with experiments and computer simulations, that Giraffa has unusually low overall leverage (“effective mechanical advantage”; EMA) for its forelimbs during walking (and presumably all gaits/speeds); and even its cousin Okapia does, to a degree; and the extinct giant giraffid Sivatherium too. This is because of its long limbs, which one might look at and call it “cursorial adaptation” but our analysis reveals the tradeoffs of that; as limb length goes up, EMA goes down, and that negatively impacts athletic abilities. All the more reason to be wary of simplistic length-speed conclusions from extinct animals (calling T. rex!). This, with the similar paper on elephant EMA we published in 2010, is one of my papers I’m proudest of; even though neither (curiously) got much (if any) media/other attention. So it goes.
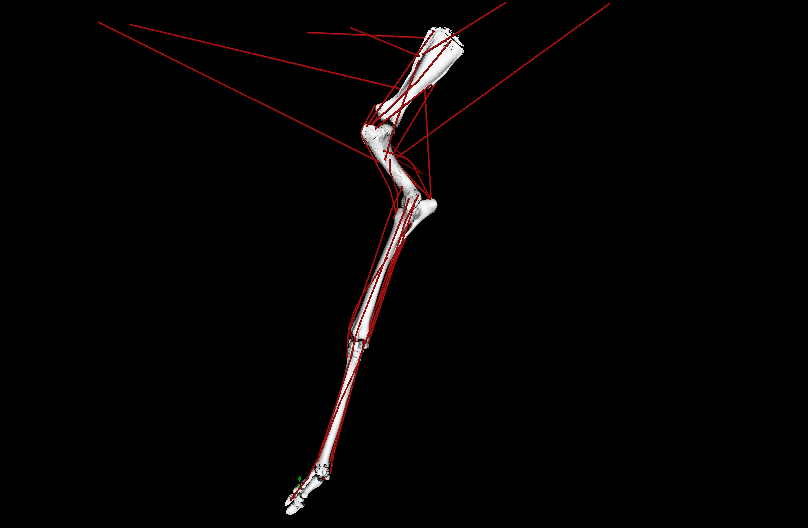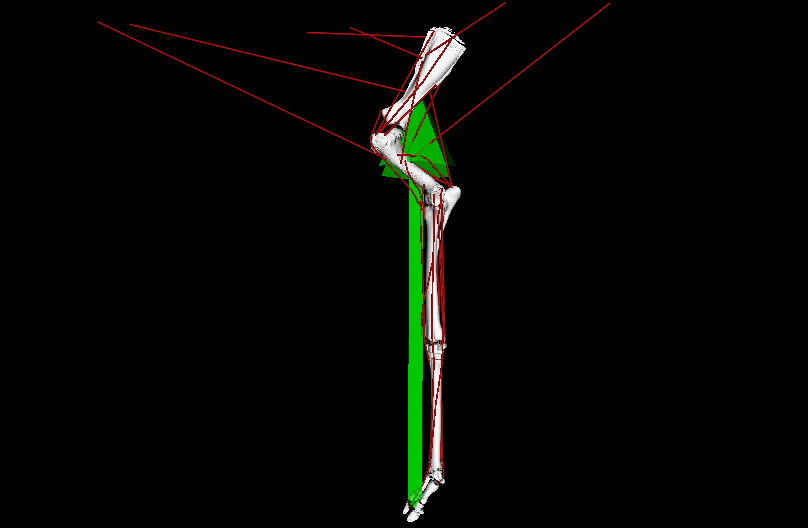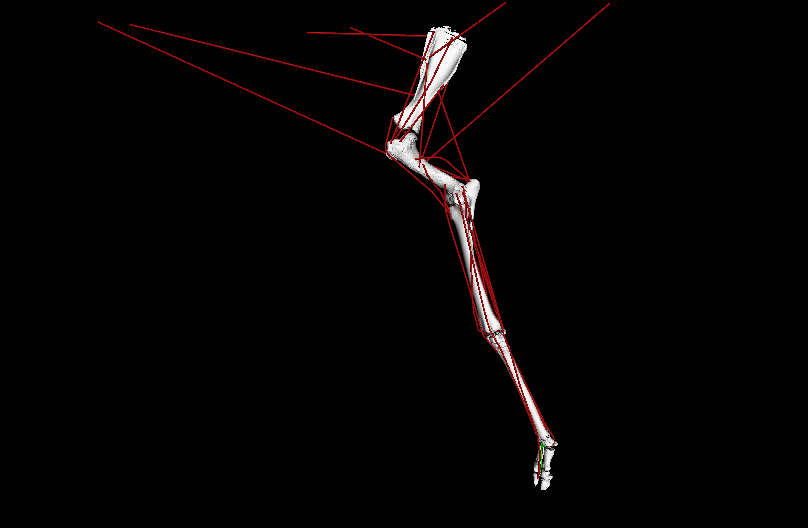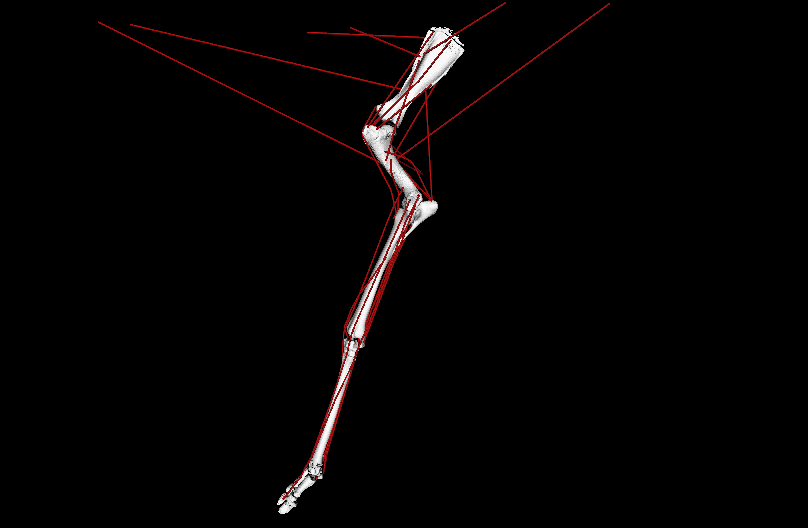
Above: OpenSim simulation of left forelimb of Giraffa during walking; in ~real-time, representing one ground-contact (stance) phase. Green arrow = ground-reaction force (GRF); red lines = major muscle lines of actions (the simulation activated/deactivated them, producing forces to counter the GRF). EMA is the ratio of the muscles’ leverage vs. GRF leverage around joints; it is ~0.3 in a giraffe vs. ~1.0 in a horse. EMA tends to be larger in larger mammals, up to horse-sized, then it gets weird in really big animals.
We also scienced the hell out of salamanders. Four papers, all involving Fire salamanders Salamandra salamandra! Three stemmed from my past PhD student Eva Herbst’s work: one explaining a new method to measure joint mobility; another applying that to walking salamanders in vivo and ex vivo; and the third comparing similar data to the Permian ‘giant’ salamander-relative Eryops, showing that its hip and knee joints were about as mobile as a Fire salamander’s. The fourth paper used video analyses of Fire salamanders in a theoretical model and simulation (with other animals) to demonstrate how multi-legged locomotion is controlled. It’s great to have these studies (partly from my old NERC grant on tetrapod locomotor evolution) out after ?5+ years; now Fire salamanders are among the salamanders whose locomotion we understand best. And we have more data still…
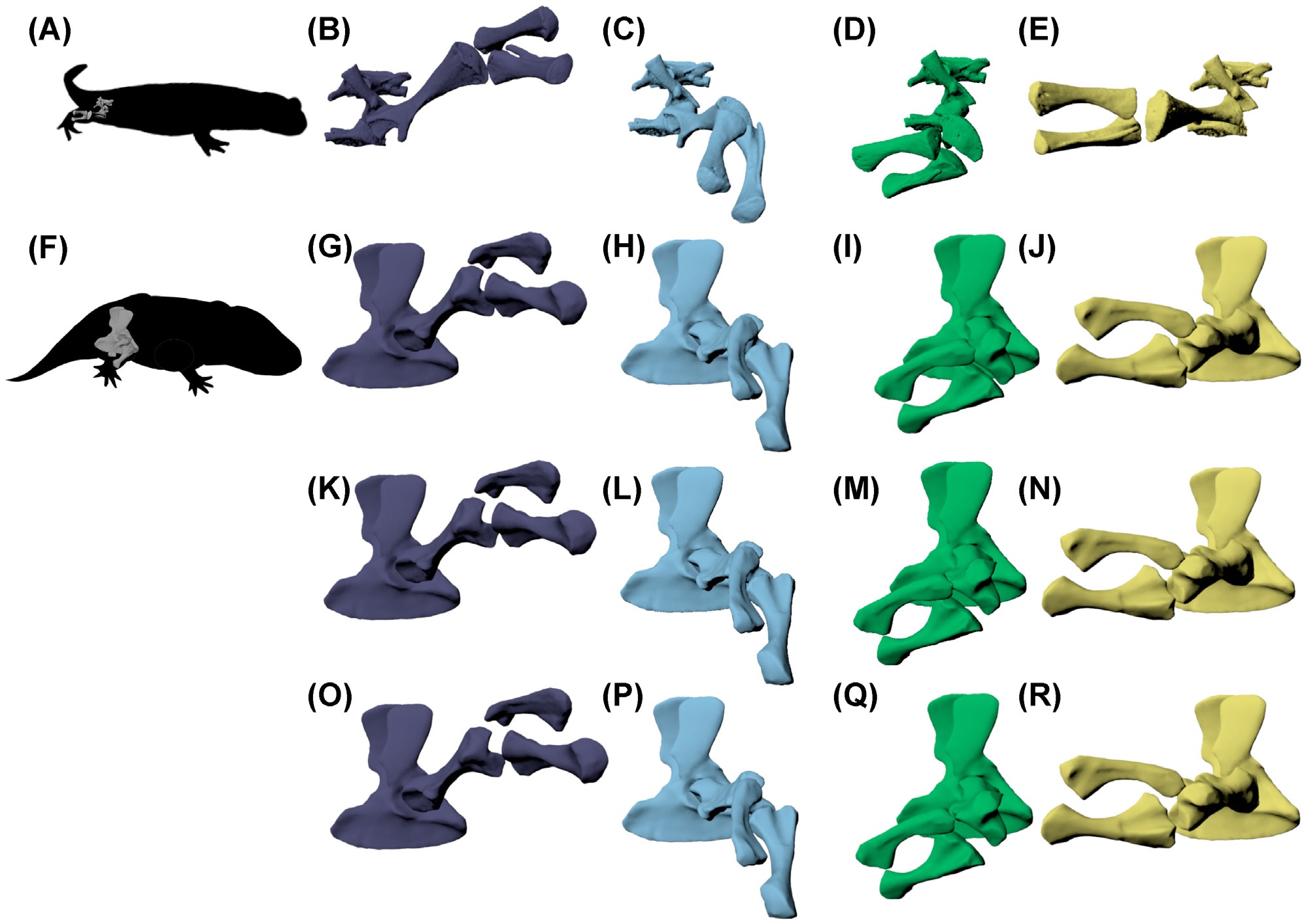
Above: Hindlimb configurations in S. salamandra (A) from rotoscoping of in vivo walking, during (B) mid-swing, (C) toe-on,(D) mid-stance, and (E) just before toe-off. These limb configurations were recreated in E. megacephalus (F) with three different knee spacing options: (G–J) tight knee spacing; (K–N) intermediate knee spacing; and (O–R) larger knee spacing, based on the amount of knee spacing present in the rotoscoped salamander at the null pose. S. salamandra configurations in (B–E) were scaled to E. megacephalus knee B.
Oh and I did some science consulting! “Prehistoric Planet” rocked the casbah; glad to see it out ~3 years after I began offering some critiques on the animations. I hope one scene I commented on eventually sees the light of day, as it wasn’t in the final programme. Similarly, “Dinosaurs: The Final Day” did well, and I gave the same kind of input. My experiences with these shows have inspired me to blog someday about how to become, and do, science consulting for documentaries, so watch for that. I may work in some commentary on what it means to be an invisible minority in that context, as I have thoughts.
There’s a lot of fun science to come, and that keeps me going. We’ve finished initial biomechanical models of 13 extinct archosaurs for DAWNDINOS, and those will become papers on modelling and simulating locomotor function, ultimately testing how performance differed between Pseudosuchia and Dinosauriformes/Dinosauria; and how locomotion evolved (e.g., bipedalism). Some examples in progress are below; these don’t show the muscles or external dimensions reconstructed. Stay tuned in 2022 and beyond for all that! Beyond this, time will tell what I’ll be doing, but DAWNDINOS is going to keep me very busy for plenty of years, and that is good fun for me.


See you in 2023!?


[…] « Year 10.5 of John’s Freezer: WTF? […]
Hi john
Me and a couple of friends have recently discovered the coolest place on the internet, your freezer. Complete laymen and wing over our heads, we’ve being jointing our mind over your ostrich research paper and blog posts trying to figure out bird/dino legs
How do ostrich ankle and knees balance themselves side to side? Is it just the shape of the joint? are the muscles involved at all? How does it work? What do they do to balance themselves if they stumble or get kicked from the side?
What about other bird joints, like wing elbows? Do they work the same? How do they keep straight during flight?
p.s.
Thanks for all the awesomeness
Well, you know the way to my heart, Jarl! Although this site is mostly dormant, I keep it up as it still represents how I feel about the awesomeness of anatomy. And I want to share that with others!
So, for bird legs: the ankles are very passive (ligaments and joints support) from left-to-right and in twisting, but like pretty much all land vertebrates they have big tendons (like our Achilles; and connected to muscles) behind the joint and smaller ones in front, for fore-aft control and springyness.
For the knees, it’s a lot of musculature packed around there, especially for fore-aft control but also in the other directions. So it’s a very 3D mechanism. There are also plenty of ligaments involved, menisci (pads), and the joint surfaces constrain the knee, too.
Balancing side-to-side is super complex and still not well understood, scientifically! It is thought that most active control (i.e. by the brain) is from the hips, and this is very 3D control by big muscles etc. There are some intriguing nervous system organs inside the hip bones/sacrum that seem to act as a sensory organ related to stability and reflexes. Then further down the legs, control probably gets more passive/automated (e.g. stretch reflexes by tendons), which is faster than anything regulating via the brain. And once down to the ankle/toes, it becomes more and more passive, so the nervous system could be entirely uninvolved, allowing quick adjustments. This is a gross generalisation.
Wings follow the same principle essentially- concentrate the more active muscular control closer to the body, reduce it further down the limb. Once at the wrist, it’s mostly passive; like the ankle. BUT the big game-changer is the feathers, connected to forearms and hands, which are mostly very passive control (but the feather bases do have tiny muscles controlling them). The wings are kept straight by muscle and by ligaments/bone-bone contacts. The wrist/hand has a remarkably nice mechanism of keeping it straight. Wings are suggested to have a “4-bar linkage” (or 6 bars? https://royalsocietypublishing.org/doi/10.1098/rsif.2017.0224 or more bars?) connecting elbow-hand motion and simplifying control.
Thank you for your awesomeness in commenting!