
Our 3D computer models of a basal dinosaur and bird, showing methods and key differences in body shape. The numbers at the bottom are museum specimen numbers.
At about the moment I’m posting this, our Nature paper (our more formal page here, and the actual article here) embargo is ending, drawing a 14+ year gestation to a close. The paper is about how dinosaur 3D body shape changed during their evolution, and how that relates to changes in hindlimb posture from early dinosaurs/archosaurs to birds; “morpho-functional evolution” sums up the topic. We used the 3D whole-body computational modelling that I, Allen and Bates (among others) have developed to estimate evolutionary changes in body dimensions, rather than focusing on single specimens or (as in our last study) tyrannosaur ontogeny. We’ve strongly supported the notion (dating back to Gatesy’s seminal 1990 Paleobiology paper) that the centre of mass of dinosaurs shifted forwards during their evolution, and that this shift gradually led to the more crouched (flexed) hind leg posture that characterizes living birds. Here is a movie from our paper showing how we did the modelling:
And here is a summary of our 17 computer models of archosaur bodies, shown as one walks along the tips of the phylogeny shown in the video (the models are not considered to be ancestral to one another; we used a common computer algorithm called squared-change parsimony to estimate ancestral state changes of body dimensions between the 16 numbered nodes of the tree):
But we’ve done much more than just put numbers on conventional wisdom.
We’ve shown, to our own surprise, that the shift of the centre of mass was largely driven by evolutionary enlargements of the forelimbs (and the head and neck, and hindlimbs, to a less strong degree), not the tail as everyone including ourselves has assumed for almost 25 years. And the timing of this shift occurred inside the theropod dinosaur group that is called Maniraptora (or Maniraptoriformes, a slightly larger group), so the change began in animals that were not flying, but not long before flight evolved (depending on whom you ask, what taxonomy they favour and what evidence one accepts, either the smaller clade Eumaniraptora/Paraves or the bird clade Aves/Avialae).
Now, if you don’t like the cliche “rewriting the textbooks”, do have a look through texts on dinosaur/early avian palaeobiology and you probably will find a discussion of how the tail shortened, the centre of mass moved forwards as a consequence, the caudofemoral musculature diminished, and theropod dinosaurs (including birds) became more crouched as a result. We did that to confirm for ourselves that it’s a pretty well-accepted idea. Our study supports a large proportion of that idea’s reasoning, but modifies the emphasis to be on the forelimbs more than the tail for centre of mass effects, so the story gets more complex. The inference about caudofemoral muscles still seems quite sound, however, as is the general trend of increased limb crouching, but our paper approximates the timing of those changes.

Figure 3 from our paper, showing how the centre of mass moved forwards (up the y-axis) as one moves toward living birds (node 16); the funny dip at the end is an anomaly we discuss in the paper.
A final implication of our study is that, because the forelimbs’ size influenced the centre of mass position, and thus influenced the ways the hindlimbs functioned, the forelimbs and hindlimbs are more coupled (via their effects on the centre of mass) than anyone has typically considered. Thus bipedalism and flight in theropods still have some functional coupling– although this is a matter of degree and not black/white, so by no means should we do away with helpful concepts like locomotor modules.
And in addition to doing science that we feel is good, we’ve gone the extra mile and presented all our data (yes, 17 dinosaurs’ worth of 3D whole body graphics!) and the critical software tools needed to replicate our analysis, in the Dryad database (link now working!), which should have now gone live with the paper! It was my first time using that database and it was incredibly easy (about 1 hour of work once we had all the final analysis’s files properly organized)– I strongly recommend others to try it out.
That’s my usual general summary of the paper, but that’s not what this blog article is about. I’ll provide my usual set of links to media coverage of the paper below, too. But the focus here is on the story behind the paper, to put a more personal spin on what it means to me (and my coauthors too). I’ll take a historical approach to explain how the paper evolved.
This paper’s story, with bits from the story of my life:

Embarassing picture of me before I became a scientist. Hardee’s fast food restaurant cashier, my first “real job”, from ~1999- no, wait, more like 1986. The 1980s-style feathered (and non-receding) hair gives it away!
Rewind to 1995. I started my PhD at Berkeley. I planned to use biomechanical methods and evidence to reconstruct how Tyrannosaurus rex moved, and started by synthesizing evidence on the anatomy and evolution of the hindlimb musculature in the whole archosaur group, with a focus on the lineage leading to Tyrannosaurus and to living birds. As my PhD project evolved, I became more interested and experienced in using 3D computational tools in biomechanics, which was my ultimate aim for T. rex.
In 1999, Don Henderson published his mathematical slicing approach to compute 3D body dimensions in extinct animals, which was a huge leap for the field forward beyond statistical estimates or physical toy models, because it represented dinosaurs-as-dinosaurs (not extrapolated reptiles/mammals/whatever) and gave you much more information than just body mass, with a lot of potential to do sensitivity analysis.
I struggled to upgrade my computer skills over the intervening years. I was developing the idea to reconstruct not only the biomechanics of T. rex, but also the evolutionary changes of biomechanics along the whole archosaur lineage to birds– because with a series of models of different species and a working phylogeny, you could do that. To me this was far more interesting than the morphology or function of any one taxon, BUT required you to be able to assess the latter. So Tyrannosaurus became a “case study” for me in how to reconstruct form and function in extinct animals, because it was interesting in its own right (mainly because of its giant size and bipedalism). (Much later, in 2007, I finally finished a collaboration to develop our own software package to do this 3D modelling, with Victor Ng-Thow-Hing and F. Clay Anderson at Honda and Stanford)

Me and a Mystery Scientist (then an undergrad; now a very successful palaeontologist!), measuring up a successful Cretaceous hypercarnivore at the UCMP; from my PhD days at Berkeley, ~2000 or so.
In all this research, I was inspired by not only my thesis committee and others at Berkeley, but also to a HUGE degree by Steve Gatesy, a very influential mentor and role model at Brown University. I owe a lot to him, and in a sense this paper is an homage to his trailblazing research; particularly his 1990 Paleobiology paper.
In 2001, I got the NSF bioinformatics postdoc I badly wanted, to go to the Neuromuscular Biomechanics lab at Stanford and learn the very latest 3D computational methods in biomechanics from Prof. Scott Delp’s team. This was a pivotal moment in my career; I became partly-an-engineer from that experience, and published some papers that I still look back fondly upon. Those papers, and many since (focused on validating and testing the accuracy/reliability of computer models of dinosaurs), set the stage for the present paper, which is one of the ones I’ve dreamed to do since the 1990s. So you may understand my excitement here…

Stanford’s Neuromuscular Biomechanics Lab, just before I left in 2003.
But the new paper is a team effort, and was driven by a very talented and fun then-PhD-student, now postdoc, Dr Vivian Allen. Viv’s PhD (2005-2009ish) was essentially intended to do all the things in biomechanics/evolution that I had run out of time/expertise to do in my PhD and postdoc, in regards to the evolution of dinosaur (especially theropod) locomotor biomechanics. And as I’d hoped, Viv put his own unique spin on the project, proving himself far better than me at writing software code and working with 3D graphics and biomechanical models. He’s now everything that I had hoped I’d become by the end of my postdoc, but didn’t really achieve, and more than that, too. So huge credit goes to Viv for this paper; it would never have happened without him.
We also got Karl Bates, another proven biomechanics/modelling expert, to contribute diverse ideas and data. Furthermore, Zhiheng Li (now at UT-Austin doing a PhD with Dr Julia Clarke) brought some awesome fossil birds (Pengornis and Yixianornis) from the IVPP in Beijing in order to microCT scan them in London. Zhiheng thus earned coauthorship on the paper — and I give big thanks to the Royal Society for funding this as an International Joint Project, with Dr Zhonghe Zhou at the IVPP.
That’s the team and the background, and I’ve already given you the punchlines for the paper; these are the primitive and the derived states of the paper. The rest of this post is about what happened behind the scenes. No huge drama or anything, but hard, cautious work and perseverance.

Me shortly after I moved to the RVC; video still frame from a dinosaur exhibit (c. 2004) I was featured in. Embarassingly goofy pic, but I like the blurb at the bottom. It’s all about the evolutionary polarity, baby!
The paper of course got started during Viv’s PhD thesis; it was one of his chapters. However, back then it was just a focus on how the centre of mass changed, and the results for those simple patterns weren’t very different from those we present in the paper. We did spot, as our Nature supplementary information notes, a strange trend in early theropods (like Dilophosaurus; to a lesser degree Coelophysis too) related to some unusual traits (e.g. a long torso) and suggested that there was a forward shift of centre of mass in these animals, but that wasn’t strongly upheld as we began to write the Nature paper.
On the urging of the PhD exam committee (and later the paper reviewers, too), Viv looked at the contributions of segment (i.e. head, neck, trunk, limbs, tail) mass and centre of mass to the overall whole body centre of mass. And I’m glad he did, since that uncovered the trend we did not expect to find: that the forelimb masses were far more important for moving the centre of mass forwards than the mass (or centre of mass) of the tail was– in other words, the statistical correlation of forelimb mass and centre of mass was strong, whereas changes of tail size didn’t correlate with the centre of mass nearly as much. We scrutinized those results quite carefully, even finding a very annoying bug in the 3D graphics files that required a major re-analysis during peer review (delaying the paper by ~6 months).
The paper was submitted to Nature first, passing a presubmission inquiry to check if the editor felt it fit the journal well enough. We had 3 anonymous peer reviewers; 1 gave extensive, detailed comments in the 3 rounds of review and was very fair and constructive, 1 gave helpful comments on writing style and other aspects of presentation as well as elements of the science, and 1 wasn’t that impressed by the paper’s novelty but wanted lots more species added, to investigate changes within different lineages of maniraptorans (e.g. therizinosaurs, oviraptorosaurs). That third reviewer only reviewed the paper for the first round (AFAIK), so I guess we won them over or else the editor overruled their concerns. We argued that 17 taxa were probably good enough to get the general evolutionary trends that we were after, and that number was ~16 more species than any prior studies had really done.
Above: CT scan reconstruction of the early extinct bird Yixianornis in slab conformation, and then Below: 3D skeletal reconstruction by Julia Molnar, missing just the final head (I find this very funny; Daffy Duck-esque) which we scaled to the fossil’s dimensions from the better data in our Archaeopteryx images.  There is also the concern, which the reviewers didn’t focus on but I could see other colleagues worrying about, that some of the specimens we used were either composites, sculpted, or otherwise not based on 100% complete, perfectly intact specimens. The latter are hard to come by for a diversity of extinct animals, especially in the maniraptoran/early bird group. We discussed some of these problems in our 3D Tyrannosaurus paper. The early dinosauromorph Marasuchus that we used was a cast/sculpted NHMUK specimen based on original material, as was our Coelophysis, Microraptor and Archaeopteryx; and our Carnegie
There is also the concern, which the reviewers didn’t focus on but I could see other colleagues worrying about, that some of the specimens we used were either composites, sculpted, or otherwise not based on 100% complete, perfectly intact specimens. The latter are hard to come by for a diversity of extinct animals, especially in the maniraptoran/early bird group. We discussed some of these problems in our 3D Tyrannosaurus paper. The early dinosauromorph Marasuchus that we used was a cast/sculpted NHMUK specimen based on original material, as was our Coelophysis, Microraptor and Archaeopteryx; and our Carnegie ??Caenagnathus??Anzu (now published) specimen was based more on measurements from 1 specimen than from direct scans, and there were a few other issues with our other specimens, all detailed in our paper’s Supplementary Information.
But our intuition, based on a lot of time spent with these models and the analysis of their data, is that these anatomical imperfections matter far, far less than the statistical methods that we employed– because we add a lot of flesh (like real animals have!) outside of the skeleton in our method, the precise morphology of the skeleton doesn’t matter much. It’s not like you need the kind of quality of anatomical detail that you need to do systematic analyses or osteological descriptive papers. The broad dimensions can matter, but those tend to be covered by the (overly, we suspect) broad error bars that our study had (see graph above). Hence while anyone could quibble ad infinitum about the accuracy of our skeletal data, I doubt it’s that bad– and it’s still a huge leap beyond previous studies, which did not present quantitative data, did not do comparative studies of multiple species, or did not construct models based on actual 3D skeletons as opposed to artists’ 2D shrinkwrapped reconstructions (the “Greg Paul method”). We also did directly measure the bodies of two extant archosaurs in our paper: a freshwater crocodile and a junglefowl (CT scan of the latter is reconstructed below in 3D).
One thing we still need to do, in future studies, is to look more carefully inside of the bird clade (Aves/Avialae) to see what’s going on there, especially as one moves closer to the crown group (modern birds). We represented modern birds with simply 1 bird: the “wild-type chicken” Red junglefowl, which isn’t drastically different in body shape from other basal modern birds such as a tinamou. Our paper was not about how diversity of body shape and centre of mass evolved within modern birds. But inspecting trends within Palaeognathae would be super interesting, because a lot of locomotor, size and body shape changes evolved therein; ostriches are probably a very, very poor proxy for the size and shape of the most recent common ancestor of all extant birds, for example, even though they seem to be fairly basal within that whole lineage. And, naturally, our study opens up opportunities for anyone to add feathers to our models and investigate aerodynamics, or to apply our methods to other dinosaur/vertebrate/metazoan groups. If the funding gods are kind to us, later this year we will be looking more closely, in particular, at the base of Archosauria and what was happening to locomotor mechanics in Triassic archosaurs…
Clickum to embiggum:

Australian freshwater crocodile, Crocodylus johnstoni; we CT scanned it in 3 pieces while visiting the Witmer lab in Ohio.

A Red junglefowl cockerel, spotted in Lampang, Thailand during one of my elephant gait research excursions there. Svelte, muscular and fast as hell. This photo is here to remind me to TAKE BLOODY PICTURES OF MY ACTUAL RESEARCH SPECIMENS SO I CAN SHOW THEM!
I’d bore you with the statistical intricacies of the paper, but that’s not very fun and it’s not the style of this blog, which is not called “What’s in John’s Software Code?”. Viv really worked his butt off to get the stats right, and we did many rounds of revisions and checking together, in addition to consultations with statistics experts. So I feel we did a good job. See the paper if that kind of thing floats your boat. Someone could find a flaw or alternative method, and if that changed our major conclusions that would be a bummer– but that’s science. We took the plunge and put all of our data online, as noted above, so anyone can do that, and that optimizes the reproducibility of science.
What I hope people do, in particular, is to use the 3D graphics of our paper’s 17 specimen-based archosaur bodies for other things– new and original research, video games, animations, whatever. It has been very satisfying to finally, from fairly early in the paper-writing process onwards, present all of the complex data in an analysis like this so someone else can use it. My past modelling papers have not done this, but I aim to backtrack and bring them up to snuff like this. We couldn’t publish open access in Nature, but we achieved reasonably open data at least, and to me that’s as important. I am really excited at a personal level, and intrigued from a professional standpoint, to see how our data and tools get used. We’ll be posting refinements of our (Matlab software-based) tools, which we’re still finding ways to enhance, as we proceed with future research.
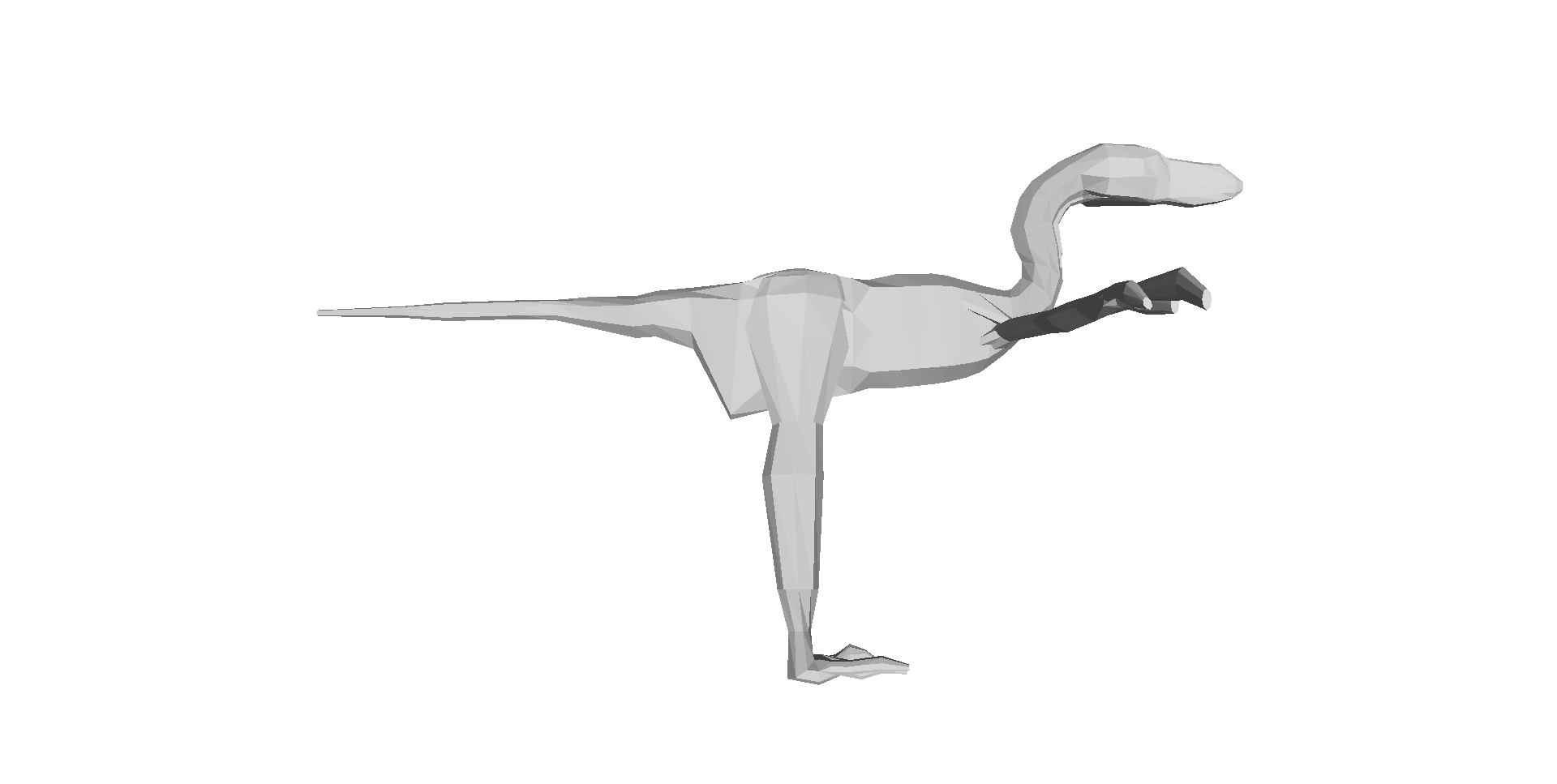
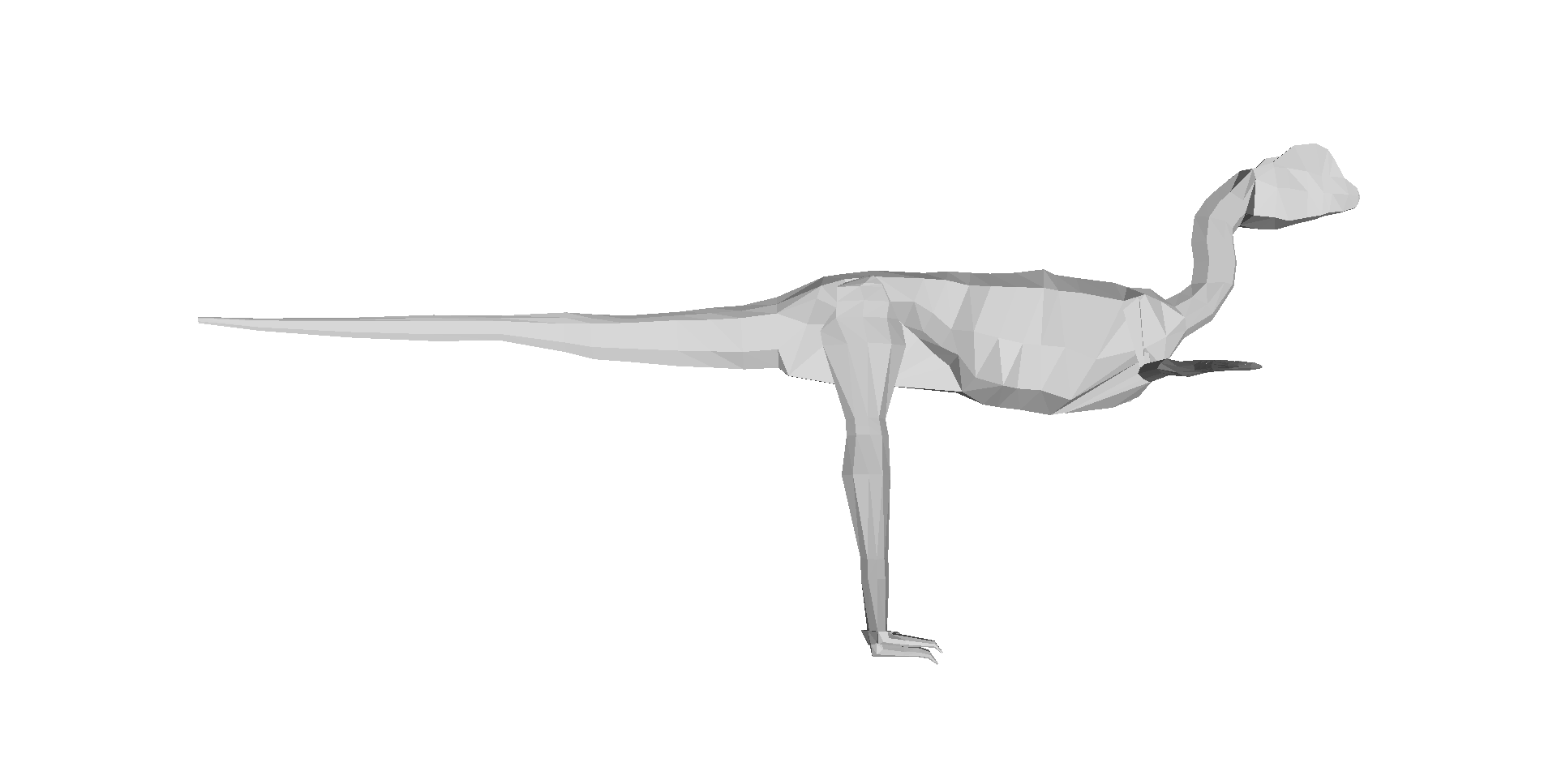
Above: Two of the 17 archosaur 3D models (the skinny “mininal” models; shrinkwrapped for your protection) that you can download and examine and do stuff with! Dilophosaurus on the left; Velociraptor on the right. Maybe you can use these to make a Jurassic Park 4 film that is better, or at least more scientifically accurate, than Hollywood’s version! 😉 Just download free software like Meshlab, drop the OBJ files in and go!
Now, to bring the story full circle, the paper is out at last! A 4 year journey from Viv’s PhD thesis to the journal, and for me a ~14 year journey from my mind’s eye to realization. Phew! The real fun begins now, as we see how the paper is received! I hope you like it, and if you work in this area I hope you like the big dataset that comes with it, too. Perhaps more than any other paper I’ve written, because of the long voyage this paper has taken, it has a special place in my heart. I’m proud of it and the work our team did together to produce it. Now it is also yours. And all 3200ish words of this lengthy blog post are, as well!
Last but not least, enjoy the wonderful digital painting that Luis Rey did for this paper (another of my team’s many failed attempts to get on the cover of a journal!); he has now blogged about it, too!

Dinosaur posture and body shape evolving up the evolutionary tree, with example taxa depicted. By Luis Rey.
News stories about this paper will be added below as they come out, featuring our favourites:
1) NERC’s Planet Earth, by Harriet Jarlett: “Dinosaur body shape changed the way birds stand”
2) Ed Yong on Phenomena: “Crouching bird, hidden dinosaur”
3) Charles Choi on Live Science: “Crouching bird, hidden evolutionary purpose?”
4) Brian Handwerk on Nat Geo Daily News: “Birds’ “Crouching” Gait Born in Dinosaur Ancestors”
5) Jennifer Viegas on Discovery News: “Heavier dino arms led evolution to birds”
6) Puneet Kollipara on Science News: “Birds may have had to crouch before they could fly”
And some superb videos- we’re really happy with these:
1) Nature’s “Crouching Turkey, Hidden Dragon”
2) Reuters TV’s “3D study shows forelimb enlargement key to evolution of dinosaurs into birds“
Synopsis: Decent coverage, but negligible coverage in the general press; just science-specialist media, more or less. I think the story was judged to be too complex/esoteric for the general public. You’d think dinosaurs, evolution, computers plus physics would be an “easy sell” but it was not, and I don’t think we made any big errors “selling” it. Interesting– I continue to learn more about how unpredictable the media can be.
Regardless, the paper has had a great response from scientist colleagues/science afficionados, which was the target audience anyway. I’m very pleased with it, too– it’s one of my team’s best papers in my ~18 year career.


[…] https://whatsinjohnsfreezer.com/2013/04/24/3d_dinosaurs/ […]
Awesome work John! I can think of one textbook that now needs to be rewritten 🙂
Thanks! Haha- actually I checked your book and yours was one of the few sources that mentioned the forelimbs’ potential effects on centre of mass, along with the tail’s.
“proving himself […] at writing software code and working with 3D graphics and biomechanical models”
Oh yes, I can attest to that: Viv is a wizard! He has helped me out tremendously with his code for generating SIMM models, but the stuff he showed me for making models from XROMM data was downright scary.
[…] encapsulates our general approach to many other problems in evolutionary biomechanics (e.g. how did avian bipedalism evolve?). The benefits are that you too can explore this theme in hands-on way, and you can talk with us […]
[…] evolved and how those influenced the position of the body’s centre of mass, similar to what we did earlier with theropod dinosaurs. My role was minor but I enjoyed the study (despite a rough ride with some early reviews) and the […]
[…] in how these little features of bird ankles originated, which we concluded to be a step along the transition to the more crouched legs that modern birds […]