It has been almost three months since my last post here, and things have fallen quiet on our sister blog Anatomy to You, too. I thought it was time for an update, which is mostly a summary of stuff we’ve been doing on my team, but also featuring some interesting images if you stick around. The relative silence here has partly been due to me giving myself some nice holiday time w/family in L.A., then having surgery to fix my right shoulder, then recovering from that and some complications (still underway, but the fact that I am doing this post is itself evidence of recovery).
Stomach-Churning Rating: 4/10; semi-gruesome x-rays of me and hippo bits at the end, but just bones really.

X-ray of my right shoulder from frontal view, unlabelled

Labelled x-ray
So my priorities shifted to those things and to what work priorities most badly needed my limited energy and time. I’ve also felt that, especially since my health has had its two-year rough patch, this blog has been quieter and less interactive than it used to be, but that is the nature of things and maybe part of a broader trend in blogs, too. My creative juices in terms of social media just haven’t been at their ~2011-2014 levels but much is out of my control, and I am hopeful that time will reverse that trend. Enough about all this. I want to talk about science for the rest of this post.
My team, and collaborators as well, have published six recent studies that are very relevant to this blog’s theme- how about we run through them quickly? OK then.
- Panagiotopoulou, O., Pataky, T.C., Day, M., Hensman, M.C., Hensman, S., Hutchinson, J.R., Clemente, C.J. 2016. Foot pressure distributions during walking in African elephants (Loxodonta africana). Royal Society Open Science 3: 160203.
Our Australian collaborators got five African elephants together in Limpopo, South Africa and walked them over pressure-measuring mats, mimicking our 2012 study of Asian elephants. While sample sizes were too limited to say much statistically, in qualitatively descriptive terms we didn’t find striking differences between the two species’ foot pressure patterns. I particularly like how the centre of pressure of each foot (i.e. abstracting all regional pressures down to one mean point over time) followed essentially the same pattern in our African and Asian elephants, with a variable heelstrike concentration that then moved forward throughout the step, and finally moved toward the outer (3rd-5th; especially 3rd) toes as the foot pushed off the ground, as below.

African elephant foot COP traces vs. time in red; Asian elephant in orange-yellow. Left and right forefeet above; hindfeet below.
Gradually, this work is moving the field toward better ability to use similar techniques to compare elephant foot mechanics among species, individuals, or over time– especially with the potential of using this method (popular in human clinical gait labs) to monitor foot (and broader musculoskeletal) health in elephants. I am hopeful that a difference can be made, and the basic science we’ve done to date will be a foundation for that.
- Panagiotopoulou, O., Rankin, J.W., Gatesy, S.M., Hutchinson, J.R. 2016. A preliminary case study of the effect of shoe-wearing on the biomechanics of a horse’s foot. PeerJ 4: e2164.
Finally, about six years after we collected some very challenging experimental data in our lab, we’ve published our first study on them. It’s a methodological study of one horse, not something one can hang any hats on statistically, but we threw the “kitchen sink” of biomechanics at that horse (harmlessly!) by combining standard in vivo forceplate analysis with “XROMM” (scientific rotoscopy with biplanar fluoroscopy or “x-ray video”) to conduct dynamic analysis of forefoot joint motions and forces (with and without horseshoes on the horse), and then to use these data as input values for finite element analysis (FEA) of estimated skeletal stresses and strains. This method sets the stage for some even more ambitious comparative studies that we’re finishing up now. And it is not in short supply of cool biomechanical, anatomical images so here ya go:
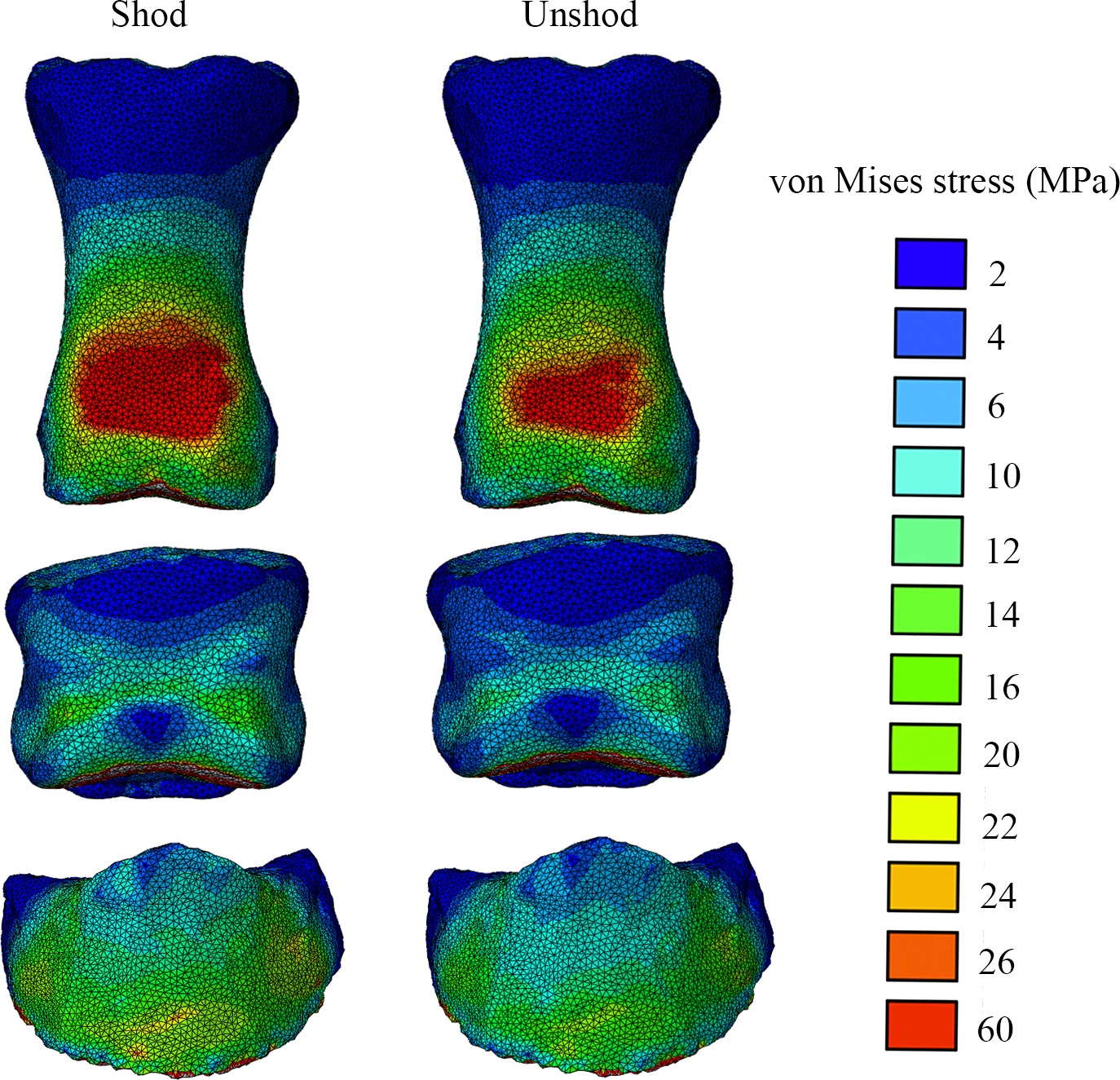
Above: The toe bones (phalanges) of our horse’s forefoot in dorsal (cranial/front) view, from our FEA results, with hot colours showing higher relative stresses- in this case, hinting (but not demonstrating statistically) that wearing horseshoes might increase stresses in some regions on the feet. But more convincingly, showing that we have a scientific workflow set up to do these kinds of biomechanical calculations from experiments to computer models and simulations, which was not trivial.
And a cool XROMM video of our horse’s foot motions:
- Bates, K.T., Mannion, P.D., Falkingham, P.L., Brusatte, S.L., Hutchinson, J.R., Otero, A., Sellers, W.I., Sullivan, C., Stevens, K.A., Allen, V. 2016. Temporal and phylogenetic evolution of the sauropod dinosaur body plan. Royal Society Open Science 3: 150636.
I had the good fortune of joining a big international team of sauropod experts to look at how the shapes and sizes of body segments in sauropods evolved and how those influenced the position of the body’s centre of mass, similar to what we did earlier with theropod dinosaurs. My role was minor but I enjoyed the study (despite a rough ride with some early reviews) and the final product is one cool paper in my opinion. Here’s an example:

The (embiggenable-by-clicking) plot shows that early dinosaurs shifted their centre of mass (COM) backwards (maybe related to becoming bipedal?) and then sauropods shifted the COM forwards again (i.e. toward their forelimbs and heads) throughout much of their evolution. This was related to quadrupedalism and giant size as well as to evolving a longer neck; which makes sense (and I’m glad the data broadly supported it). But it is also a reminder that not all sauropods moved in the same ways- the change of COM would have required changes in how they moved. There was also plenty of methodological nuance here to cover all the uncertainties but for that, see the 17 page paper and 86 pages of supplementary material…
- Randau, M., Goswami, A., Hutchinson, J.R., Cuff, A.R., Pierce, S.E. 2016. Cryptic complexity in felid vertebral evolution: shape differentiation and allometry of the axial skeleton. Zoological Journal of the Linnean Society 178:183-202.
Back in 2011, Stephanie Pierce, Jenny Clack and I tried some simple linear morphometrics (shape analysis) to see how pinniped (seal, walrus, etc) mammals changed their vertebral morphology with size and regionally across their backbones. Now in this new study, with “Team Cat” assembled, PhD student Marcela Randau collected her own big dataset for felid (cat) backbones and applied some even fancier techniques to see how cat spines change their shape and size. We found that overall the vertebrae tended to get relatively more robust in larger cats, helping to resist gravity and other forces, and that cats with different ecologies across the arboreal-to-terrestrial spectrum also changed their (lumbar) vertebral shape differently. Now Marcela’s work is diving even deeper into these issues; stay tuned…

Example measurements taken on felid vertebrae, from the neck (A-F) to the lumbar region (G-J), using a cheetah skeleton.
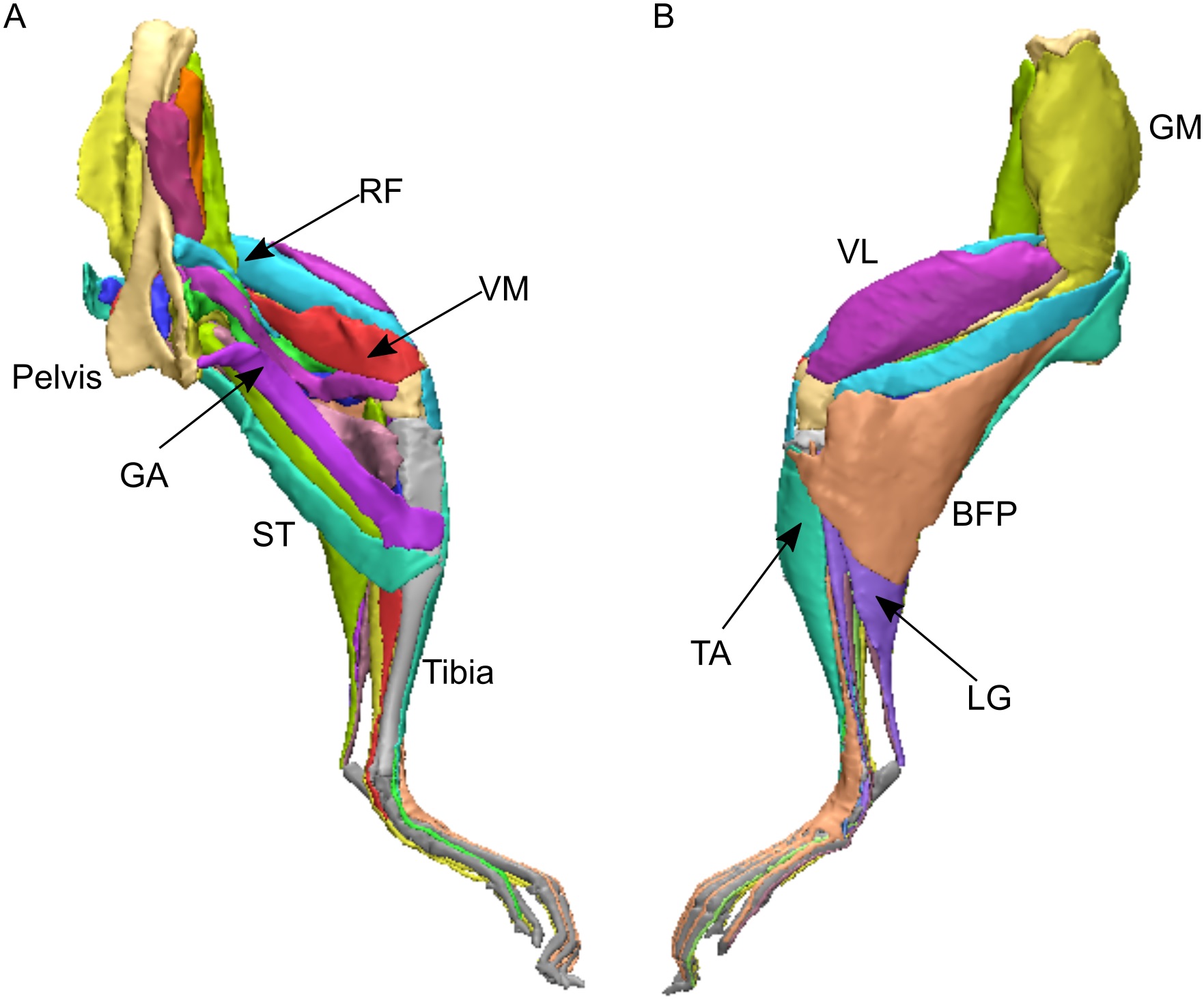
- Charles, J.P., Cappellari, O., Spence, A.J., Hutchinson, J.R., Wells, D.J. 2016. Musculoskeletal geometry, muscle architecture and functional specialisations of the mouse hindlimb. PLOS One 11(4): e0147669.
RVC PhD student James Charles measured the heck out of some normal mice, dissecting their hindlimb muscle anatomy, and using microCT scans produced some gorgeous images of that anatomy too. In the process, he also quantified how each muscle is differently specialized for the ability to produce large forces, rapid contractions or fine control. Those data were essential for the next study, where we got more computational!
- Charles, J.P., Cappellari, O., Spence, A.J., Wells, D.J., Hutchinson, J.R. 2016. Muscle moment arms and sensitivity analysis of a mouse hindlimb musculoskeletal model. Journal of Anatomy 229:514–535.
James wrangled together a lovely musculoskeletal model of our representative mouse subject’s hindlimb in the SIMM software that my team uses for these kinds of biomechanical analyses. As we normally do as a first step, we used the model to estimate things that are hard to measure directly, such as the leverages (moment arms) of each individual muscle and how those change with limb posture (which can produce variable gearing of muscles around joints). James has his PhD viva (defense) next week so good luck James!

The horse and mouse papers are exemplars of what my team now does routinely. For about 15 years now, I’ve been building my team toward doing these kinds of fusion of data from anatomy, experimental biomechanics, musculoskeletal and other models, and simulation (i.e. estimating unmeasurable parameters by telling a model to execute a behaviour with a given set of criteria to try to perform well). Big thanks go to collaborator Jeff Rankin for helping us move that along lately. Our ostrich study from earlier this year shows the best example we’ve done yet with this, but there’s plenty more to come.
I am incredibly excited that, now that my team has the tools and expertise built up to do what I’ve long wanted to do, we can finally deliver the goods on the aspirations I had back when I was a postdoc, and which we have put enormous effort into pushing forward since then. In addition to new analyses of horses and mice and other animals, we’ll be trying to push the envelope more with how well we can apply similar methods to extinct animals, which brings new challenges– and evolutionary questions that get me very, very fired up.
Here we are, then; time has brought some changes to my life and work and it will continue to as we pass this juncture. I suspect I’ll look back on 2016 and see it as transformative, but it hasn’t been an easy year either, to say the least. “Draining” is the word that leaps to mind right now—but also “Focused” applies, because I had to try to be that, and sometimes succeeded. I’ve certainly benefited a lot at work from having some talented staff, students and other collaborators cranking out cool papers with me.
I still have time to do other things, too. Once in a while, a cool critter manifests in The Freezers. Check out a hippo foot from a CT scan! It’s not my best scan ever (noisy data) but it shows the anatomy fairly well, and some odd pathologies such as tiny floating lumps of mineralized soft tissue here and there. Lots to puzzle over.


Welcome back!
Hope the shoulder feels better! Very interested in the horse paper – our horse was shod until recently. I always wanted it to go barefoot, and finally got my way. Finding the right foot shape now seems to be the biggest challenge.Just a head’s up – horses that have been wearing shoes for any length of time have different shaped feet than horses that have been barefoot for ages – there is a roll to the hoof that must change the dynamics of the step and how stress is applied to the different parts of the foot and leg.
Thanks Jennifer. There is a lot of scientific work on how shoes influence horses’ foot dynamics. Our study was mainly about methods and not about how a horse’s foot changes across its lifetime; even the shod/unshod part was just preliminary and about how 1 horse’s foot behaved differently at one time-slice in its life. It’s interesting how many questions are left about horses’ feet and how much less we know about most other species, even pigs and cows let alone elephants or rhinos’ feet. Lots left to do!
[…] from results based on our linear vertebral data from the beginning of the year (Randau, Goswami, et al. 2016), the 3-D vertebral coordinates carry a lot more information and we […]