Today is the 210th anniversary of Charles R. Darwin’s birthday so I put together a quick post. I’d been meaning to blog about some of our latest scientific papers, so I chose those that had an explicit evolutionary theme, which I hope Chuck would like. Here they are, each with a purty picture and a short explainer blurb! Also please check out Anatomy To You’s post by Katrina van Grouw on Darwin’s fancy pigeons.
Stomach-Churning Rating: 1/10 science!
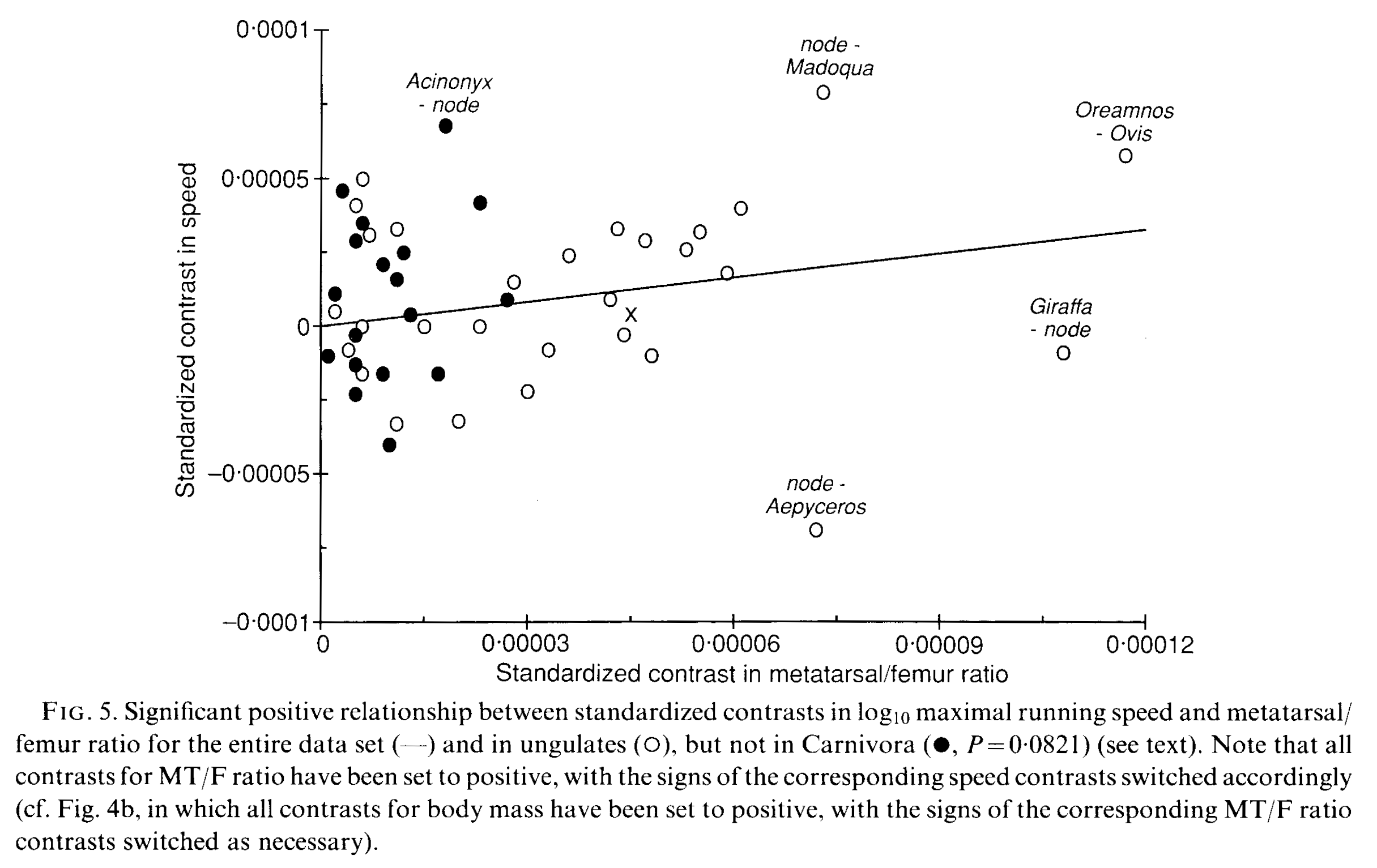
First, Brandon Kilbourne at the Naturkunde Museum in Berlin kindly invited me to assist in a paper from his German fellowship studying mustelid mammals (otters, weasels, wolverines, badgers, etc.; stinky smaller carnivorous mammals). Here we (very much driven by Brandon; I was along for the ride) didn’t just look at how forelimb bone shape changes with body size in this ecologically diverse group. We already knew bigger mustelids would have more robust bones, although it was cool to see how swimming-adapted and digging-adapted mustelids evolved similarly robust bones; whereas climbing ones had the skinniest bones.
The really exciting and novel (yes I am using that much-abused word!) aspect of the paper is that Brandon conjured some sorcery with the latest methods for analysing evolutionary trends, to test how forelimb bone shapes evolved. Was their pattern of evolution mostly a leisurely “random walk” or were there early bursts of shape innovation in the mustelid tree of life, or did shape evolve toward one or more optimal shapes (e.g. suited to ecology/habitat)? We found that the most likely pattern involved multiple rates of evolution and/or optima, rather than a single regime. And it was fascinating to see that the patterns of internal shape change deviated from external shape change such as bone lengths: so perhaps selection sometimes works independently at many levels of bone morphology?

Various evolutionary models applied to the phylogeny of mustelids.
Then there, coincidentally, was another paper originating in part from the same museum group in Berlin. This one I’d been involved in as a co-investigator (author) on a Volkswagen (yes! They like science) grant back about 8 years ago and since. There is an amazing ~290 million year old fossil near-amniote (more terrestrial tetrapod) called Orobates pabsti, preserved with good skeletal material but also sets of footprints that match bones very well, allowing a rare match of the two down to this species level. John Nyakatura’s team had 3D modelled this animal before, so we set out to use digital techniques to test how it did, or did not, move—similar to what I’d tried before with Tyrannosaurus, Ichthyostega and so forth. The main question was whether Orobates moved in a more “ancestral” salamander-like way, a more “derived” lizard-like way (i.e. amniote-ish), or something else.
The approach was like a science sledgehammer: we combined experimental studies of 4 living tetrapods (to approximate “rules” of various sprawling gaits), a digital marionette of Orobates (to assess how well its skeleton stayed articulated in various motions), and two robotics analysis (led by robotics guru Auke Ijspeert and his amazing team): a physical robot version “OroBOT” (as a real-world test of our methods), and a biomechanical simulation of OroBOT (to estimate hard-to-measure things in the other analyses, and matches of motions to footprints). And, best of all, we made it all transparent: you can go play with our interactive website, which I still find very fun to explore, and test what motion patterns do or do not work best for Orobates. We concluded that a more amniote-like set of motions was most plausible, which means such motions might have first evolved outside of amniotes.

OroBOT in tha house!
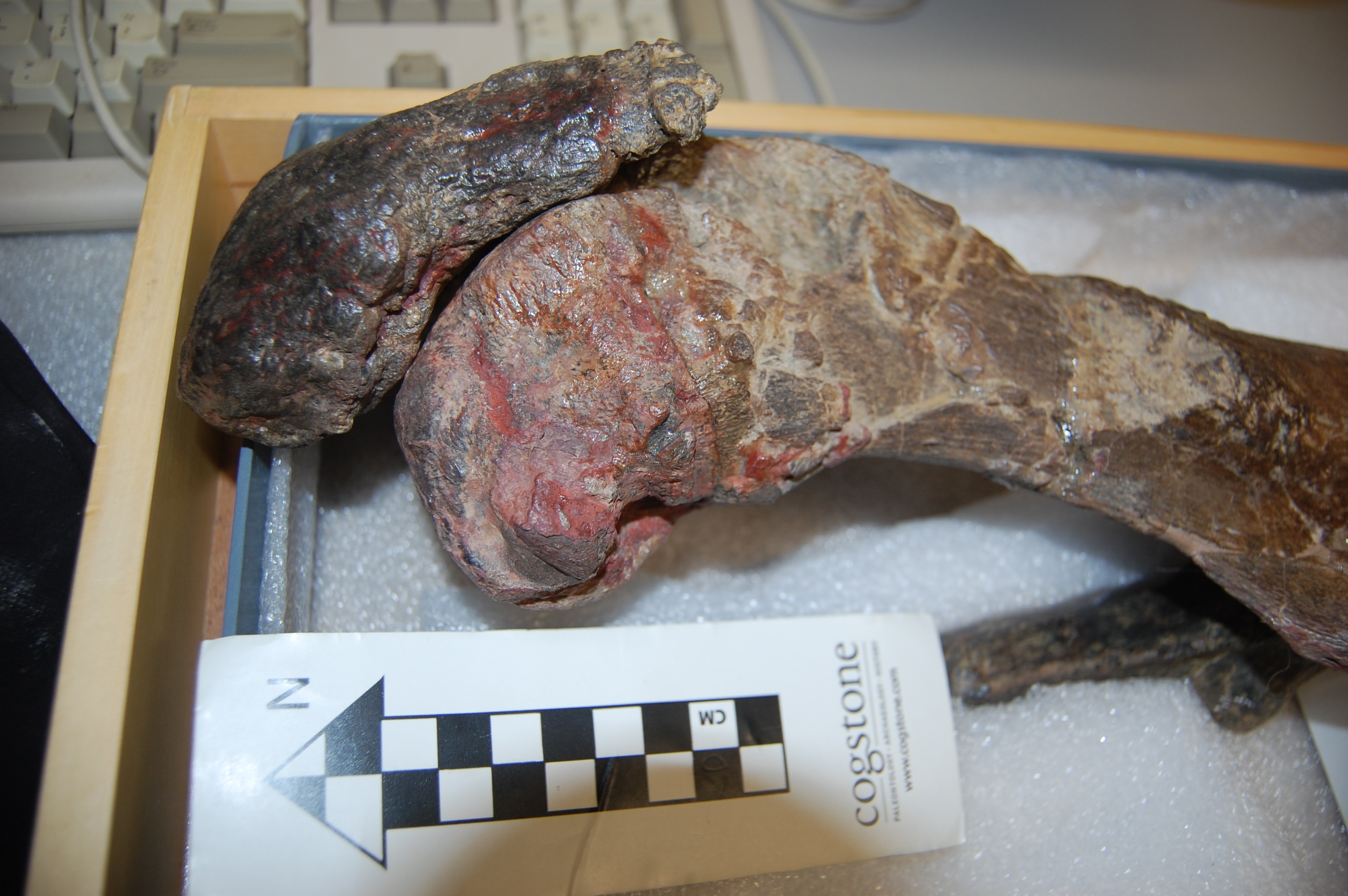
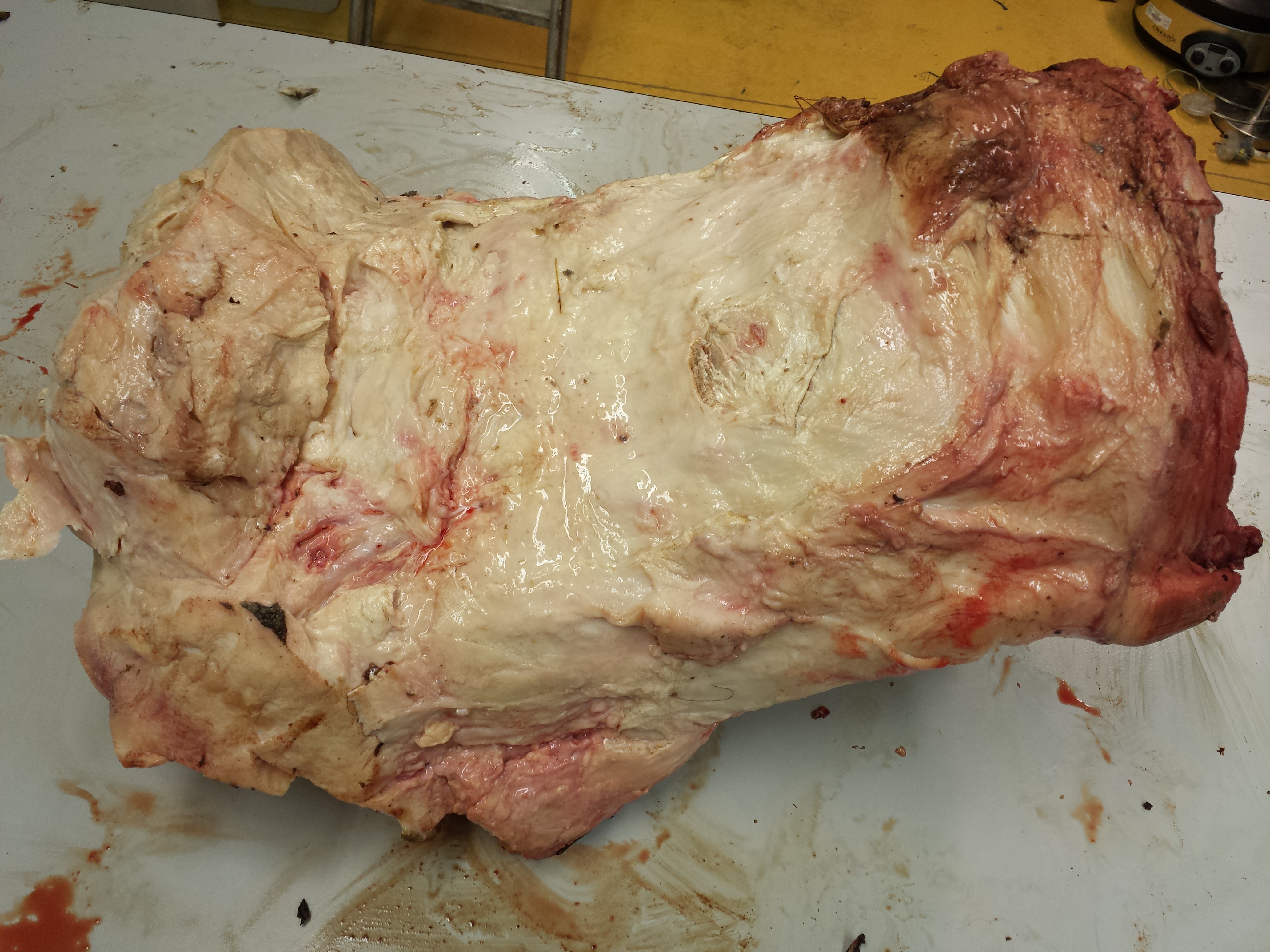
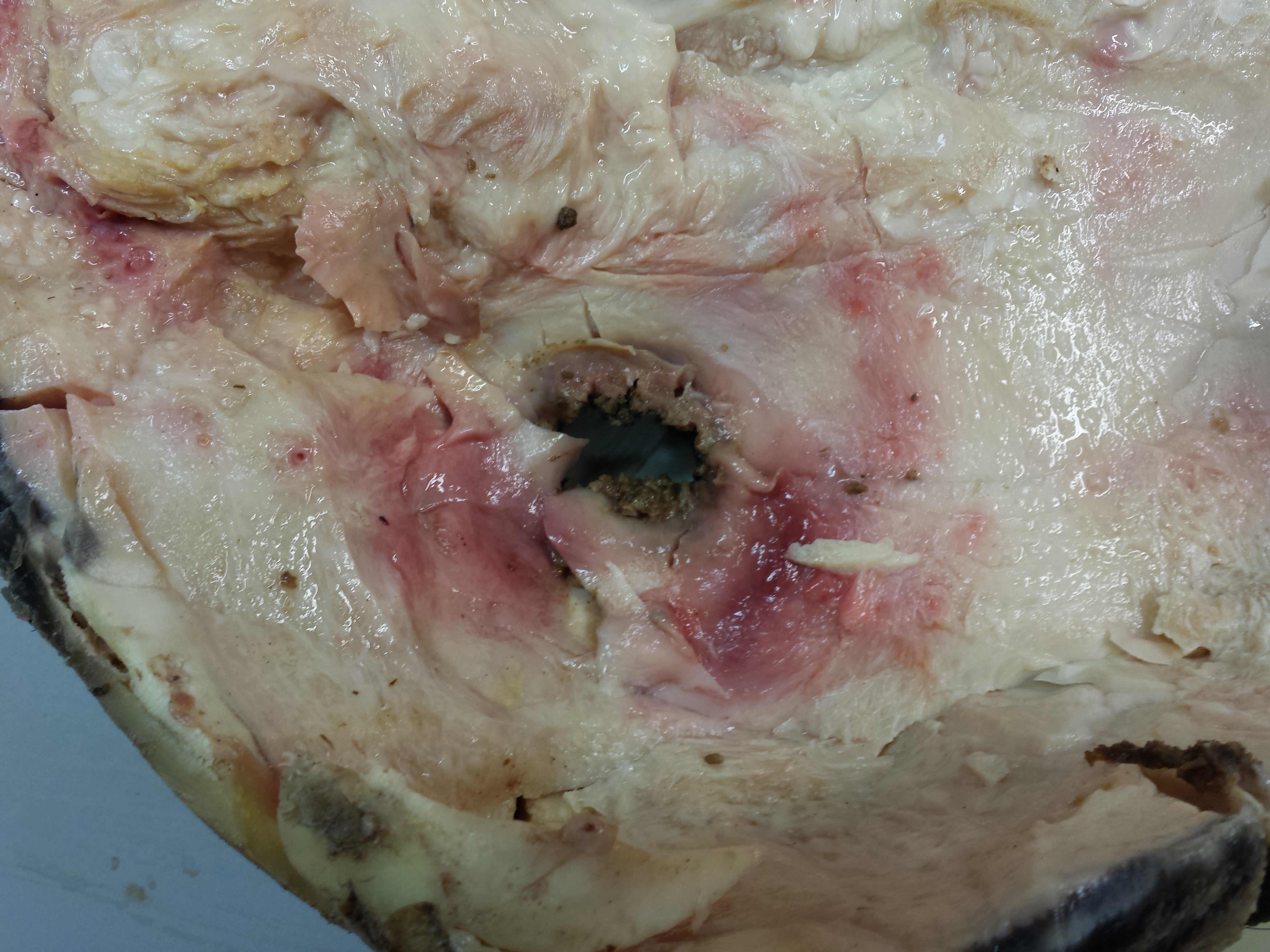
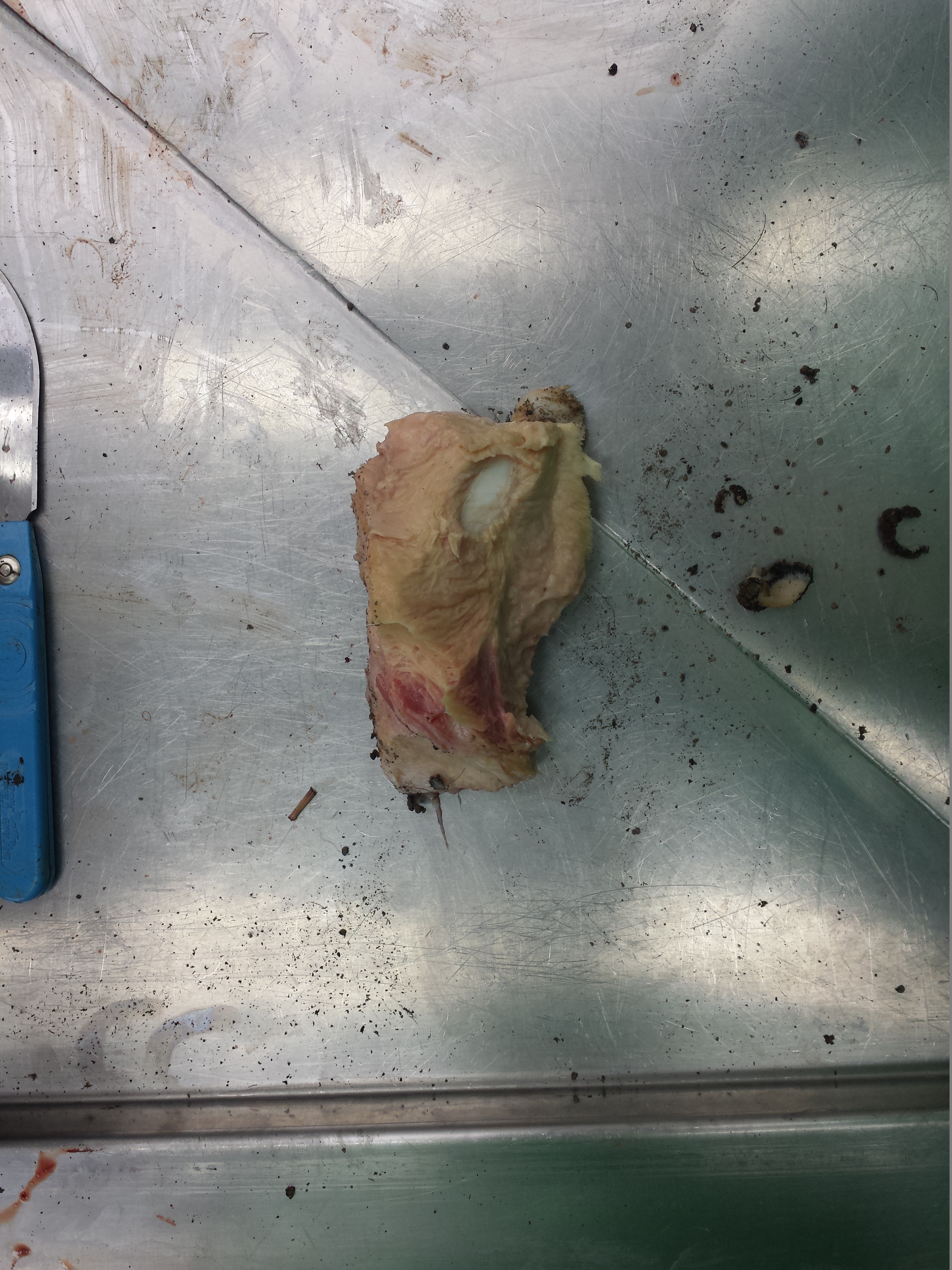
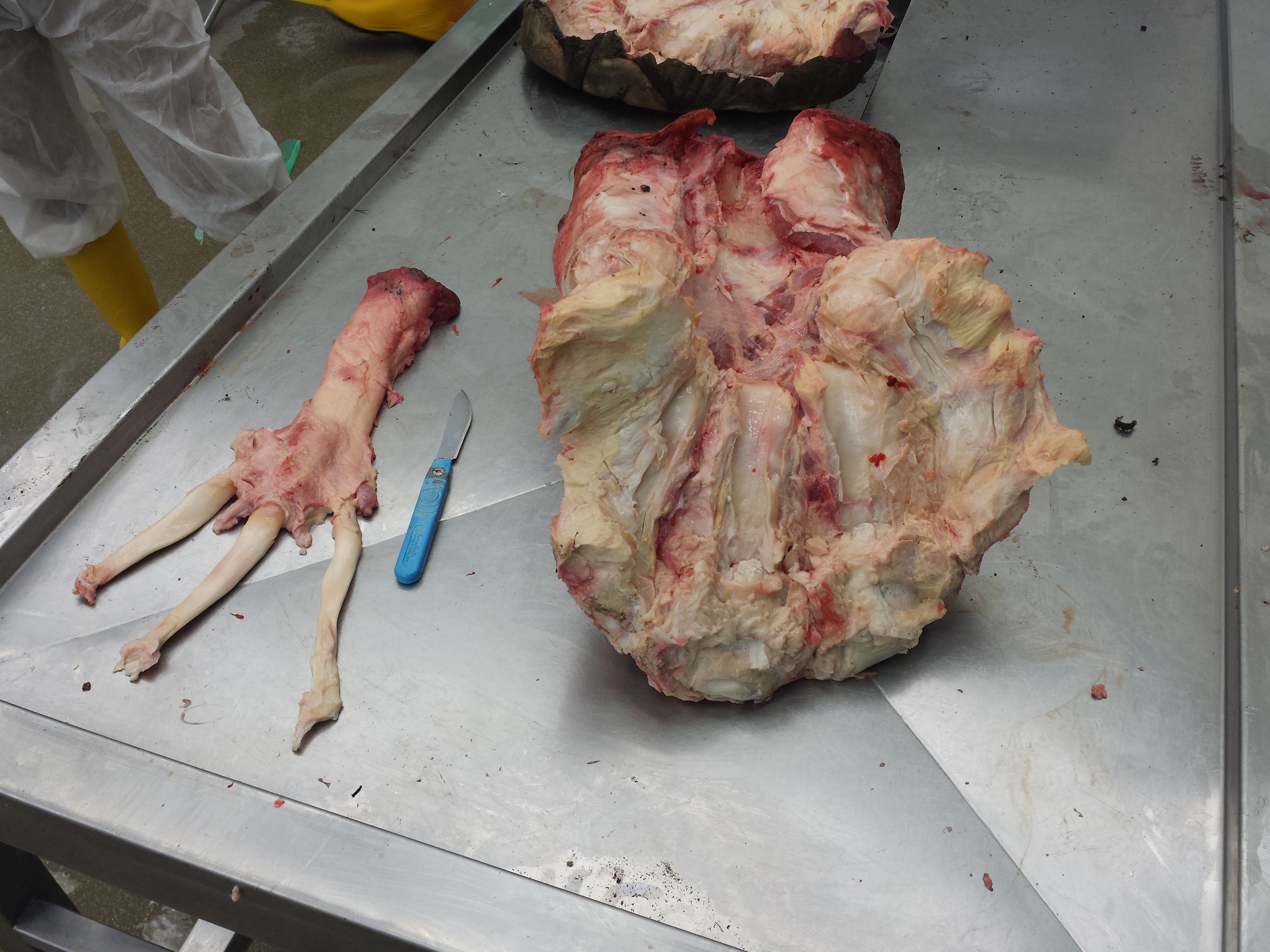
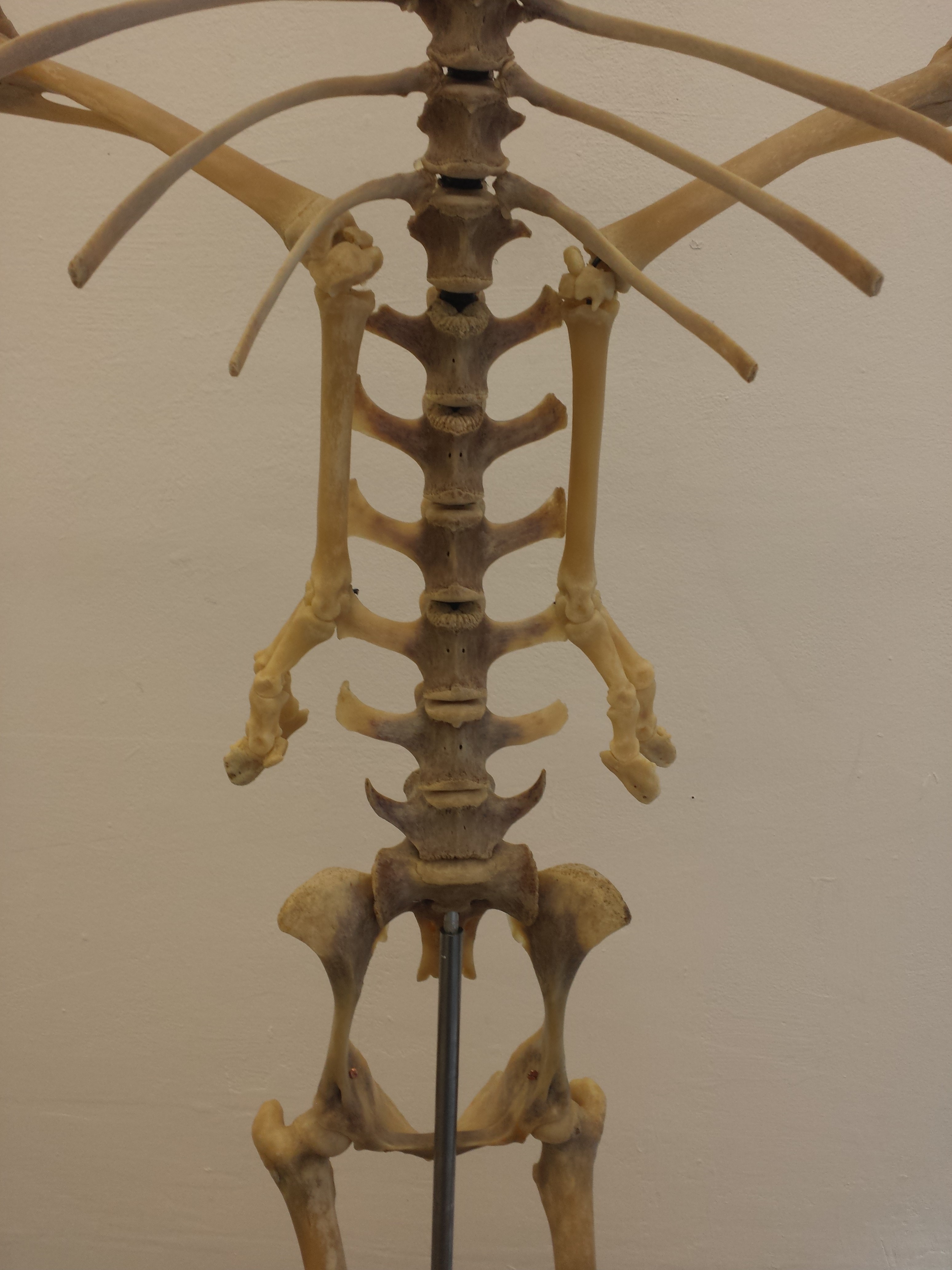
You may remember Crassigyrinus, the early tetrapod, from a prior post on Anatomy To You. My PhD student Eva Herbst finished her anatomical study of the best fossils we could fit into a microCT-scanner and found some neat new details about the “tadpole from hell”. Buried in the rocky matrix were previously unrecognized bones: vertebrae (pleurocentra; the smaller nubbins of what may be “rhachitomous” bipartite classic tetrapod/omorph structure), ribs (from broad thoracic ones to thin rear ones), pelvic (pubis; lower front), and numerous limb bones. One interesting trait we noticed was that the metatarsals (“sole bones” of the foot) were not symmetrical from left-to-right across each bone, as shown below. Such asymmetry was previously used to infer that some early tetrapods were terrestrial, yet Crassigyrinus was uncontroversially aquatic, so what’s up with that? Maybe this asymmetry is a “hangover” from more terrestrial ancestry, or maybe these bones get asymmetrical for non-terrestrial reasons.

The oddly asymmetrical metatarsals of Crassigyrinus.
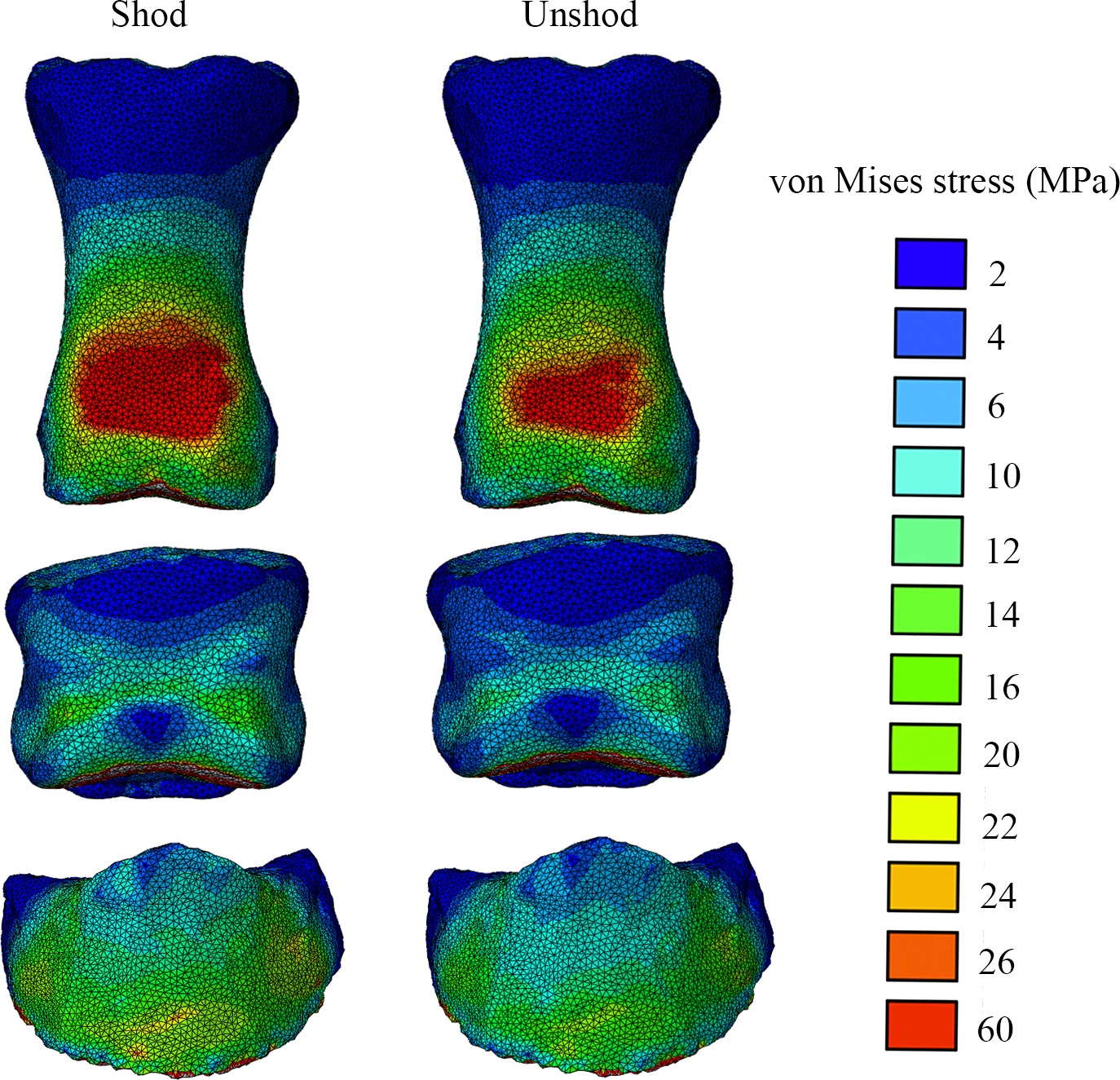
Finally, Dr. Peter Bishop finished his PhD at Griffith University in Australia and came to join us as a DAWNDINOS postdoc. He blasted out three of his thesis chapters (starting here) with me and many others as coauthors, all three papers building on a major theme: how does the inner bone structure (spongy or cancellous bone) relate to hindlimb function in theropod dinosaurs (including birds) and how did that evolve? Might it tell us something about how leg posture or even gait evolved? There are big theories in “mechanobiology” variously named Wolff’s Law or the Trajectorial Theory that explain why, at certain levels, bony struts tend to align themselves to help resist certain stresses, and thus their alignment can be “read” to indicate stresses. Sometimes. It’s complicated!
Undaunted, Peter measured a bunch of theropod limb bones’ inner geometry and found consistent differences in how the “tracts” of bony struts, mainly around joints, were oriented. He then built a biomechanical model of a chicken to test if the loads that muscles placed on the joints incurred stresses that matched the tracts’ orientations. Hmm, they did! Then, with renewed confidence that we can use this in the fossil record to infer approximate limb postures, Peter scanned and modelled a less birdlike Daspletosaurus (smaller tyrannosaur) and more birdlike “Troodon” (now Stenonychosaurus; long story). Nicely fitting many other studies’ conclusions, Peter found that the tyrannosaur had a more straightened hindlimb whereas the troodontid had a more crouched hindlimb; intermediate between the tyrannosaur and chicken. Voila! More evidence for a gradual evolution of leg posture across Mesozoic-theropods-into-modern-birds. That’s nice.

Three theropods, three best-supported postures based on cancellous bone architecture.
If you are still thirsty for more papers even if they are less evolutionary, here’s the quick scoop on ones I’ve neglected until now:
(1) Former PhD student Chris Basu published his thesis work w/us on measuring giraffe walking dynamics with force plates, finding that they move mostly like other quadrupeds and their wobbly necks might cost them a little.
(2) Oh, and Chris’s second paper just came out as I was writing this! We measured faster giraffe gaits in the wilds of South Africa, as zoo giraffes couldn’t safely do them. And we found they don’t normally go airborne, just using a rotary gallop (not trot, pace or canter); unlike some other mammals. Stay tuned: next we get evolutionary with this project!
(2) How do you safely anaesthetize a Nile crocodile? There’s now a rigorous protocol (from our DAWNDINOS work).
(3) Kickstarting my broad interest in how animals do “extreme” non-locomotor motions, we simulated how greyhounds stand up, finding that even without stretchy tendons they should, barely, be able to do it, which is neat. Expect much more about this from us in due time.
(4) Let’s simulate some more biomechanics! Ashley Heers, an NSF research fellow w/me for a year, simulated how growing chukar birds use their wing muscles to flap their way up steeper inclines (“WAIR” for devotees), and the results were very encouraging for simulating this behaviour in more detail (e.g. tendons seem to matter a lot) and even in fossil species; and finally…
(5) Hey did you ever think about how bone shape differs between hopping marsupials (macropods) and galloping artiodactyl (even-toed) mammals? We did, in long-the-making work from an old BBSRC grant with Michael Doube et al., and one cool thing is that they mostly don’t change shape with body size that differently, even though one is more bipedal at faster speeds—so maybe it is lower-intensity, slower behaviours that (sometimes?) influence bone shape more?
So there you have the skinny on what we’ve been up to lately, messing around with evolution, biomechanics and morphology.