Uh oh, a “why?” question in biology! There are many potential, and not mutually exclusive, answers to such questions. Ultimately there is a historical, evolutionary answer that underpins it all (“ostriches evolved two kneecaps because…”). But we like ostrich knees and their funky double-kneecaps (patellae; singular = patella) so we wanted to know why they get so funky. One level of addressing that question is more like a “how?” they have them. So we started there, with what on the surface is a simple analysis. And we published that paper this week, with all of the supporting data (CT, MRI, FEA).
Stomach-Churning Rating: 6/10 because there is a gooey image of a real dissection later in the post, not just tidy 3D graphics.
First author Kyle Chadwick was my research technician for 2 years on our sesamoid evolution grant, and we reported earlier on the detailed 3D anatomy of ostrich knees (this was all part of his MRes degree with me, done in parallel with his technician post). Here, in the new paper with Sandra Shefelbine and Andy Pitsillides, we took that 3D anatomy and subjected it to some biomechanical analysis in two main steps.

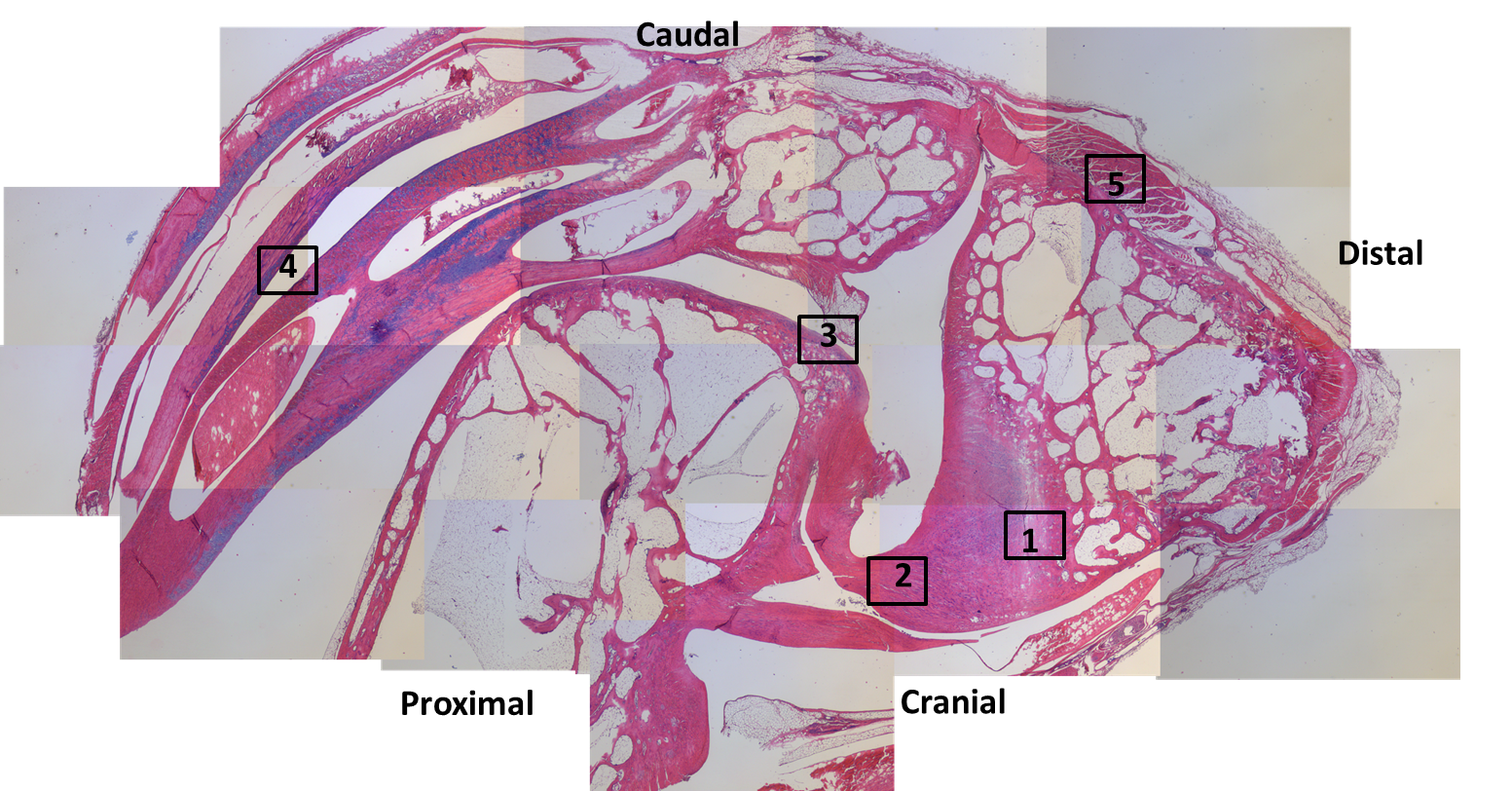
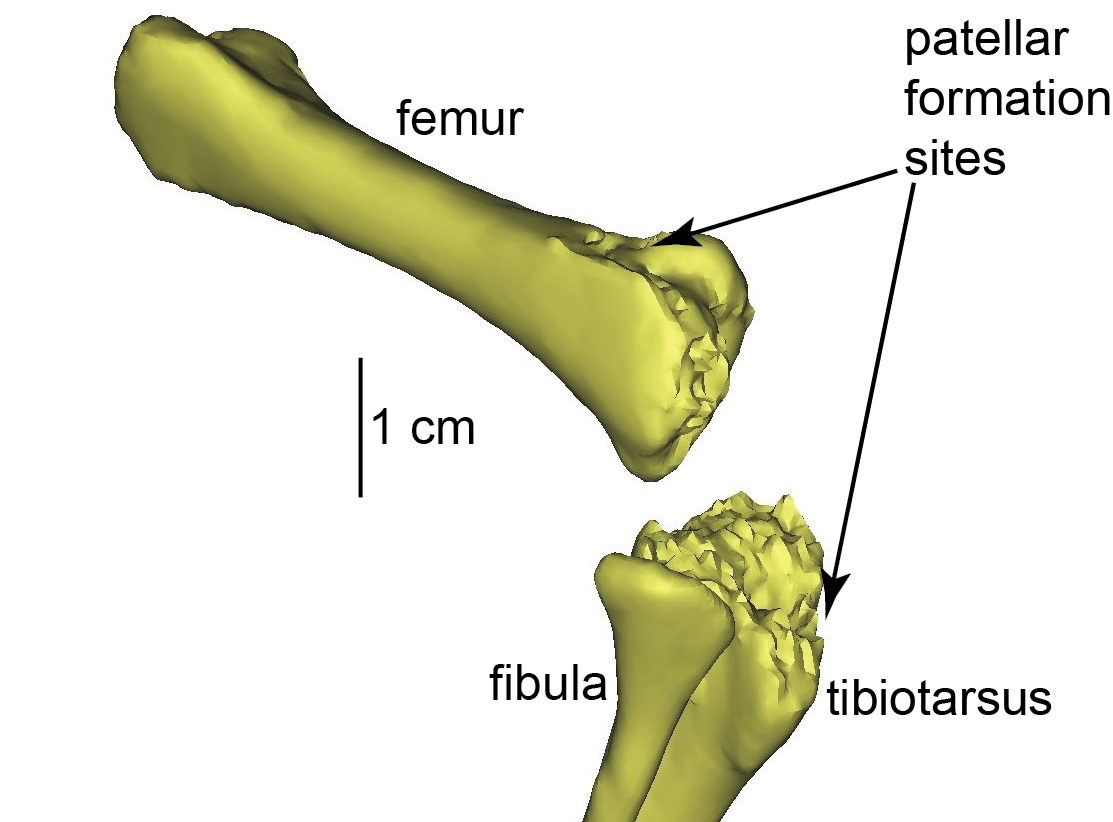
Ostrich (right) knee bones. The patellae are the two knobbly bits in the knee.
First, we used our previous biomechanical simulation data from an adult ostrich (from our paper by Rankin et al.) to estimate the in vivo forces that the knee muscles exert onto the patellar region during moderately large loading in running (not maximal speed running, but “jogging”). That was “just” (Kyle may laugh at the “just”– it wasn’t trivial) taking some vectors out of an existing simulation and adding them into a detailed 3D model. We’ve done similar things before with a horse foot’s bones (and plenty more to come!), but here we had essentially all of the soft tissues, too.

Ostrich knee with muscles as 3D objects.
Second, the 3D model that the muscular forces were applied to was a finite element model: i.e., the original 3D anatomical model broken up into a mesh, whose voxels each had specific properties, such as resistance to shape change under loading in different directions. The response of that model to the loads (a finite element analysis; FEA) gave us details on the stresses (force/area) and strains (deformations from original shape) in each voxel and overall in anatomical regions.

Finite element model setup for our study. If you do FEA, you care about these things. If not, it’s a pretty, sciencey picture.
The great thing about a computer/theoretical model is that you can ask “what if?” and that can help you understand “how?” or even “why?” questions that experiments alone cannot address. Ostriches aren’t born with fully formed bony kneecaps; indeed those patellae seem to mature fairly late in development, perhaps well after hatching. We need to know more about how the patellae form but they clearly end up inside the patellar (knee extensor) tendon that crosses the knee. So we modelled our adult ostrich without bony patellae; just with a homogeneous patellar tendon (using the real anatomy of that tendon with the bony bits replaced by tendon); and subjected it to the loading environment for “jogging”.
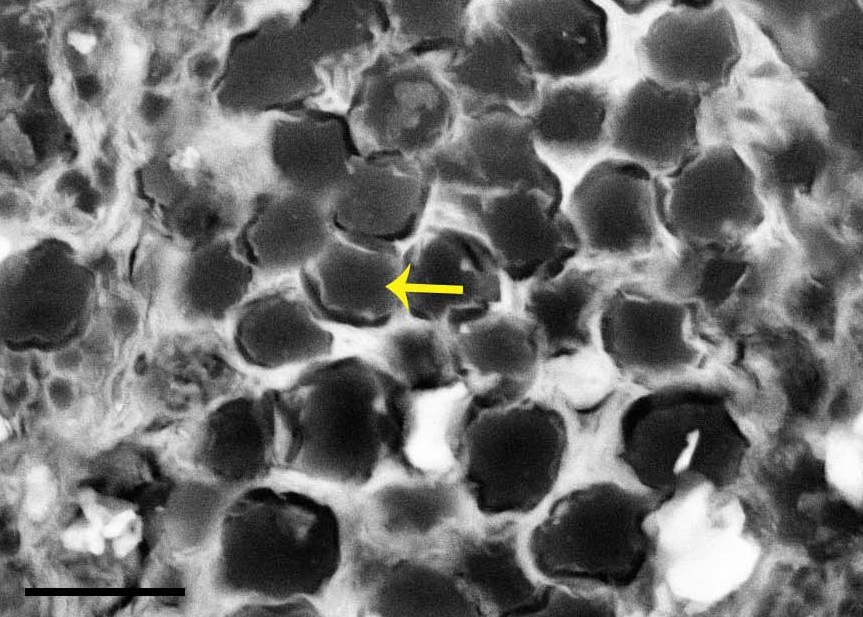
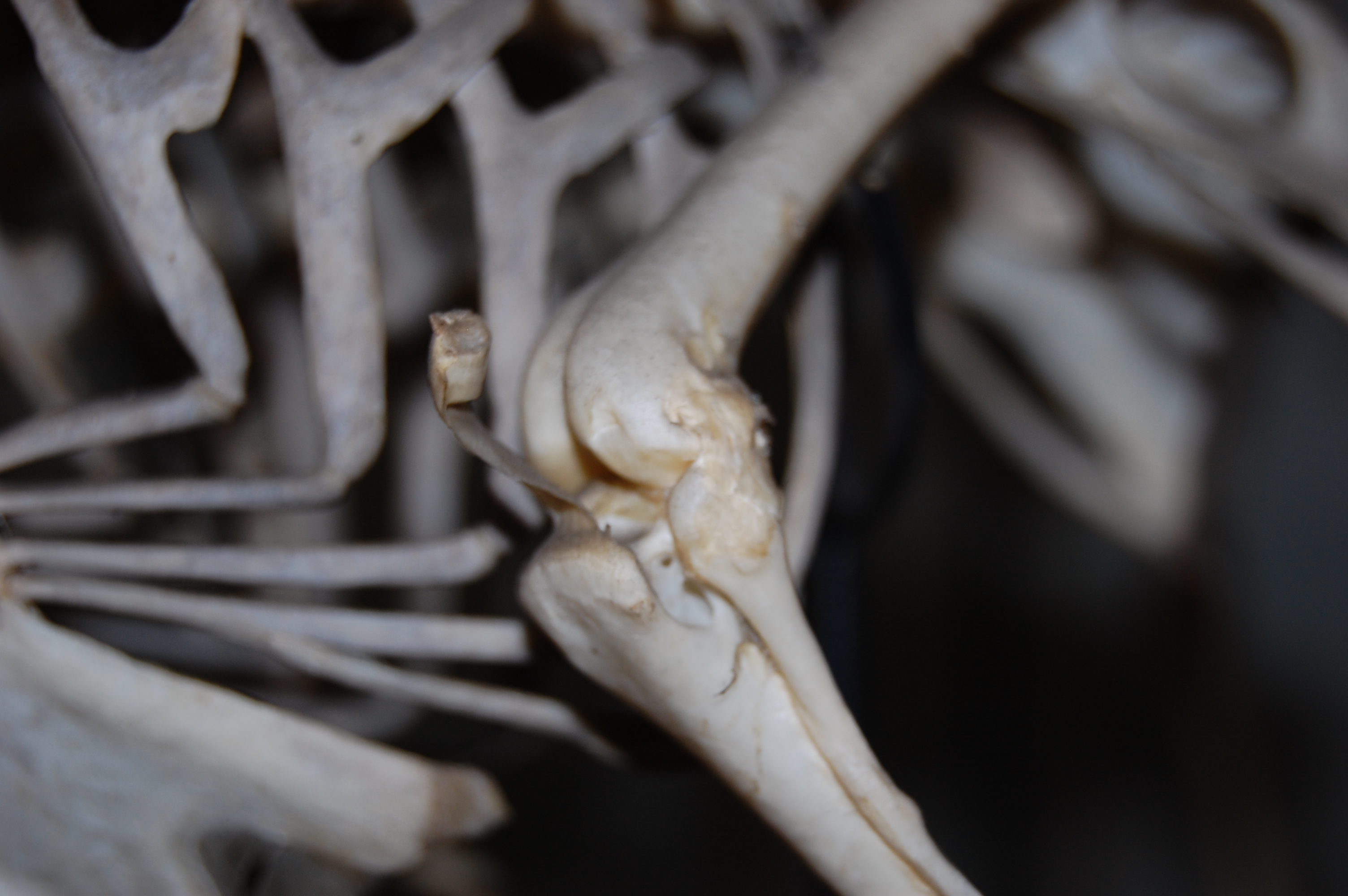
The right knee of an ostrich hatchling. The patellae have yet to form; indeed there is little bone around the knee region at all, yet.
We then inspected our FEA’s results in light of modern theory about how tissues respond to loading regimes. That “mechanobiology” theory, specifically “tissue differentiation”, postulates that tendon will tend to turn into fibrocartilage if it is subjected to high compression (squishing) and shear (pushing). Then, the fibrocartilage might eventually be reworked into bone as it drops the compression and shear levels. So, according to that theory (and all else being equal; also ignoring the complex intermediate states that would happen in reality), the real ostrich’s kneecaps should be located in the same positions where the FEA, under the moderately large loads we applied, predicts the homogeneous tendon to have high compression and shear. But did the real anatomy match the mechanical environment and tissue differentiation theory’s predictions?
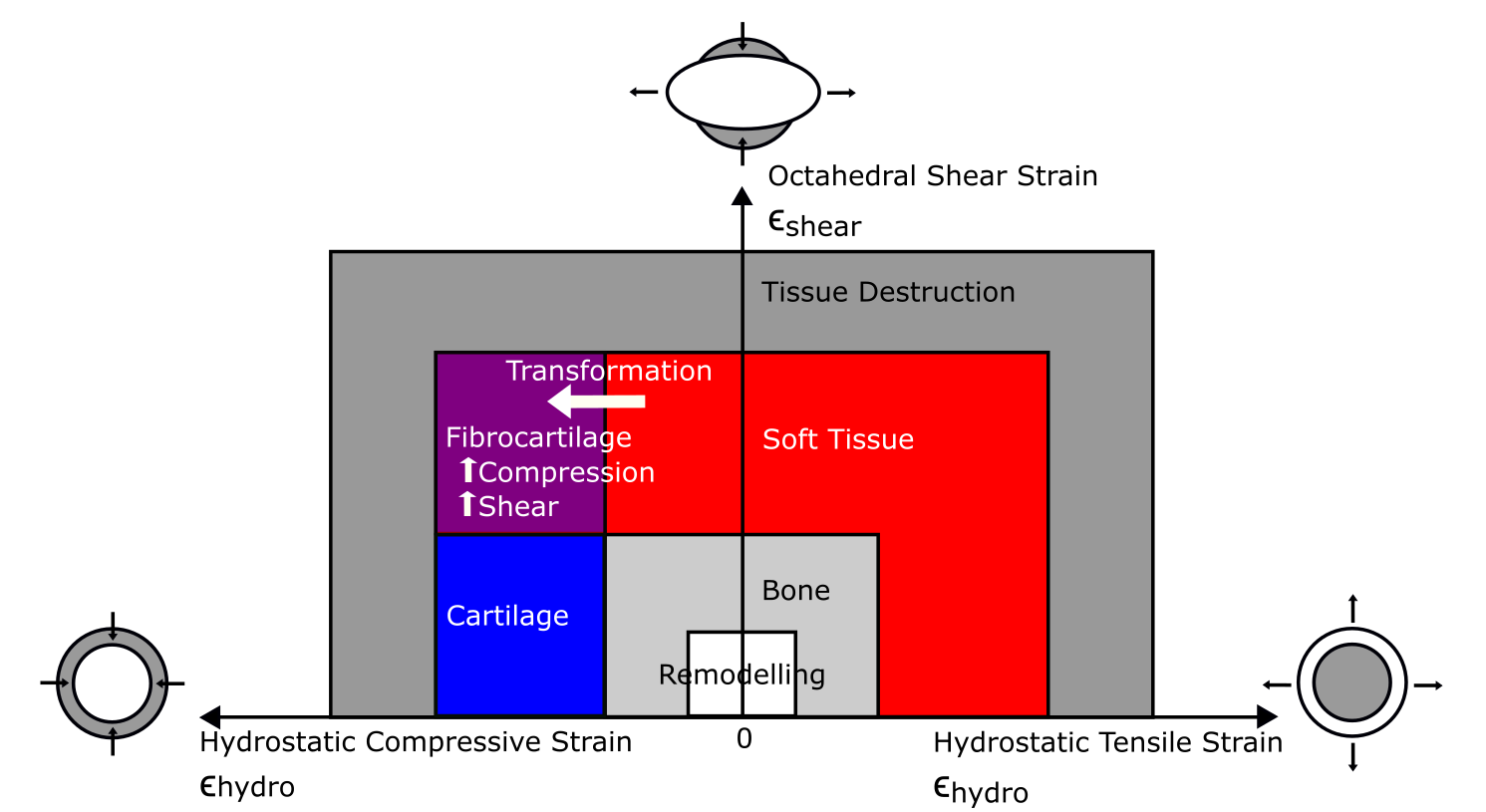
Tissue differentiation diagram displaying the theoretical pathways for transformation of tissues. If tendon (red) experiences high shear (going up the y-axis) and high compression (going toward the left), it should turn into fibrocartilage (purple). Transformation into bone (diagonally to the bottom right) would reduce the shear and compression.
Well, sort of. The image below takes some unpacking but you should be able to pick out the red areas on the bottom row where the patellae actually are, and the yellow shaded regions around some of those patellar regions are where the compression and shear regimes are indeed high and overlapping the actual patellar regions. The upper two rows show the levels of compression (or tension; pulling) and shear, but the bottom row gets the point across. It’s not a bad match overall for the first (“real”; common to all living birds) patella, located on top of the upper knee (femur). It’s not a good match overall for the second (unique to ostriches) patella, located below the first one (and attached to the tibia bone).

FEA results! (click to embiggen)
Kyle says, “Being a part of this project was exciting because of the application of engineering concepts to interesting biological (including evolutionary) questions. Also, it never gets old seeing people’s reactions when I tell them I study ostrich knees.”
The study had a lot of nuances and assumptions. We only looked at one instant in slow running and only at one adult ostrich, not at the full development of ostrich anatomy and loading. That’s harder. We started simple. The tissue differentiation theory is used more for fracture healing than for sesamoid bone formation but there’s some reason to suspect that similar mechanisms are at play in both. And there’s much more; if you want the gory details see the paper.
So did we solve why, or how, ostriches have two kneecaps? We felt that the mechanical environment of our FEA was a good theoretical explanation of where the first patella forms. We originally expected the second patella, which evolved more recently and might be more mechanically sensitive as a result, to be a better match than the first one, but it was the opposite. C’est la science!
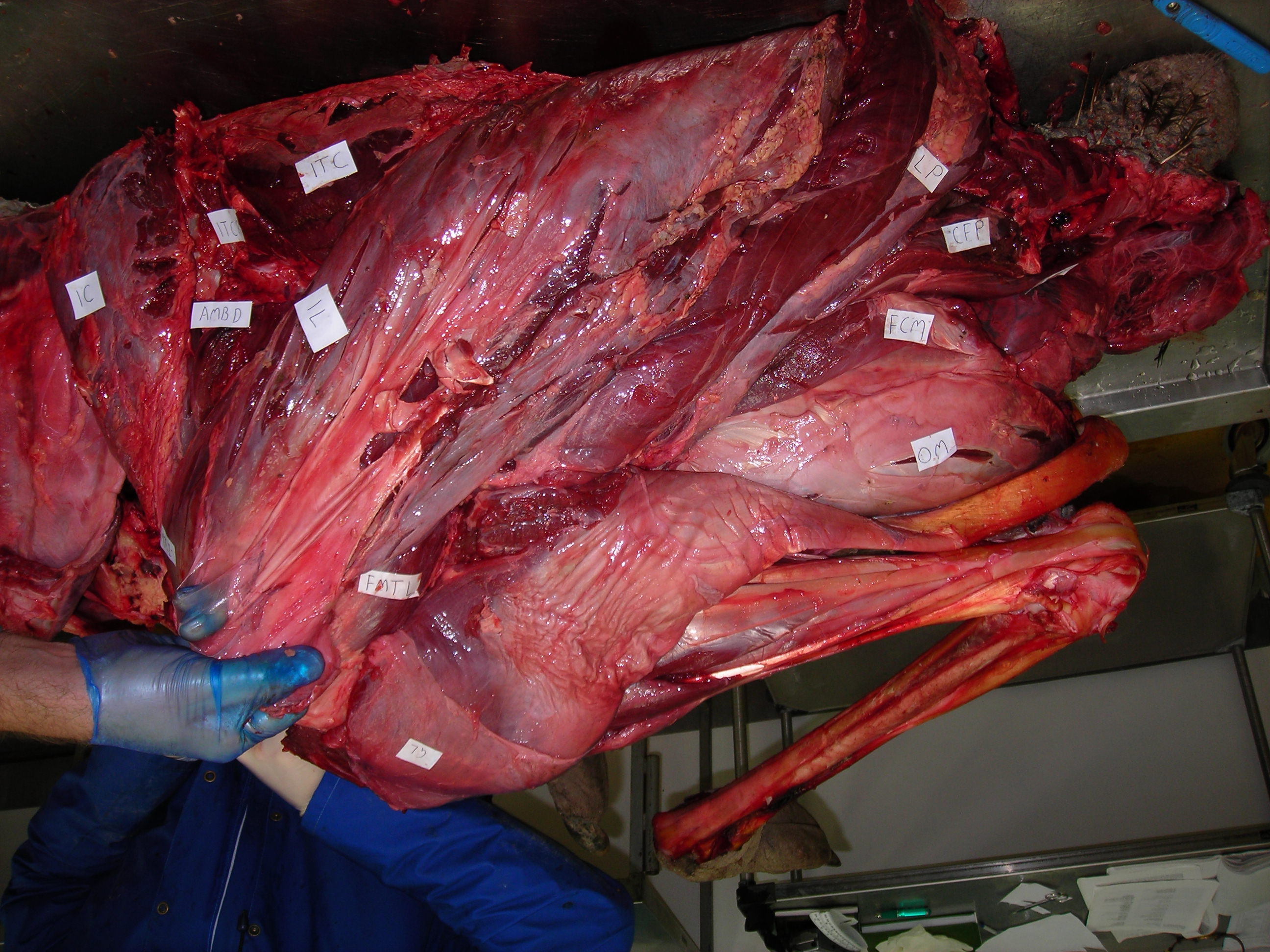
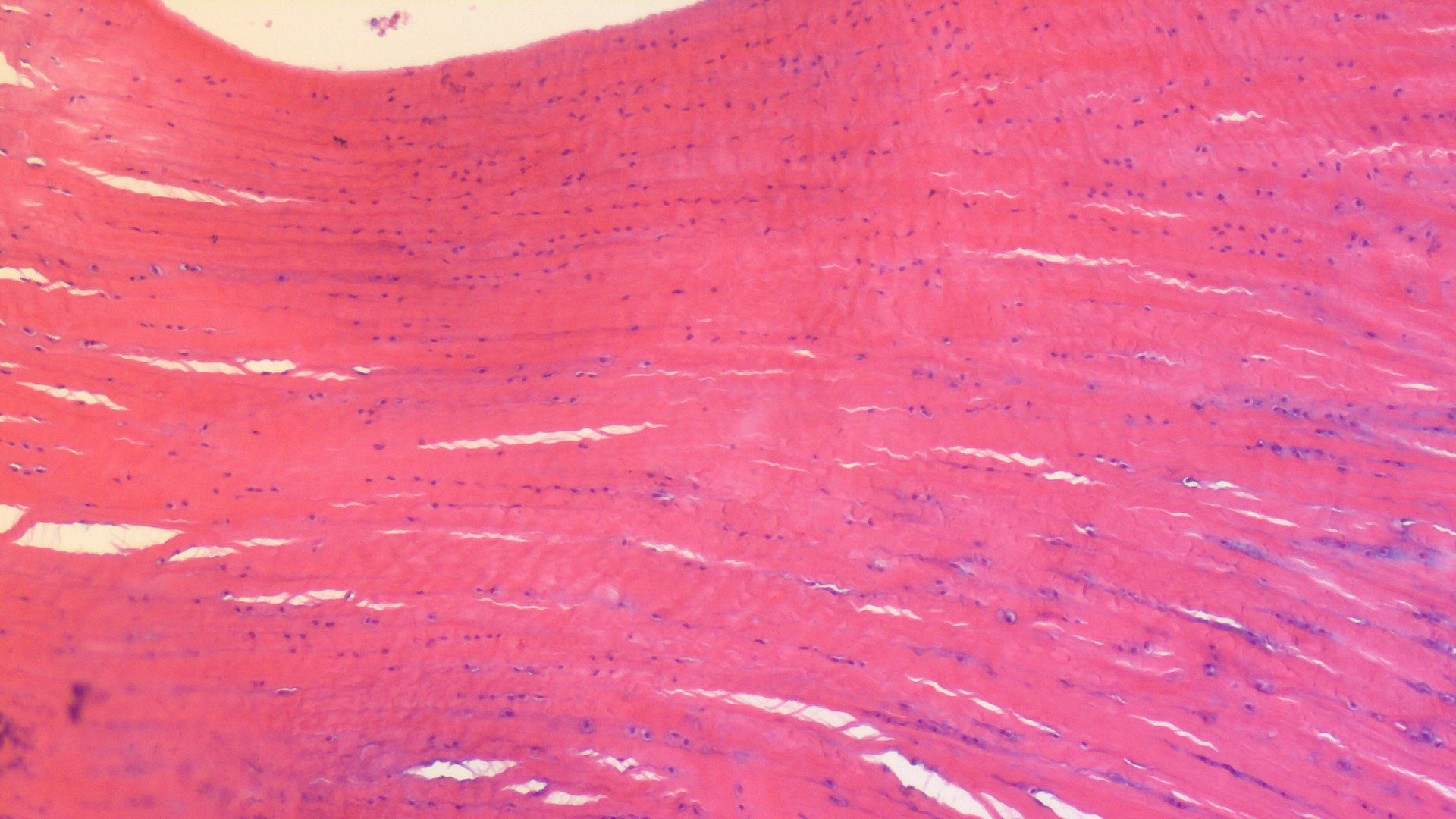
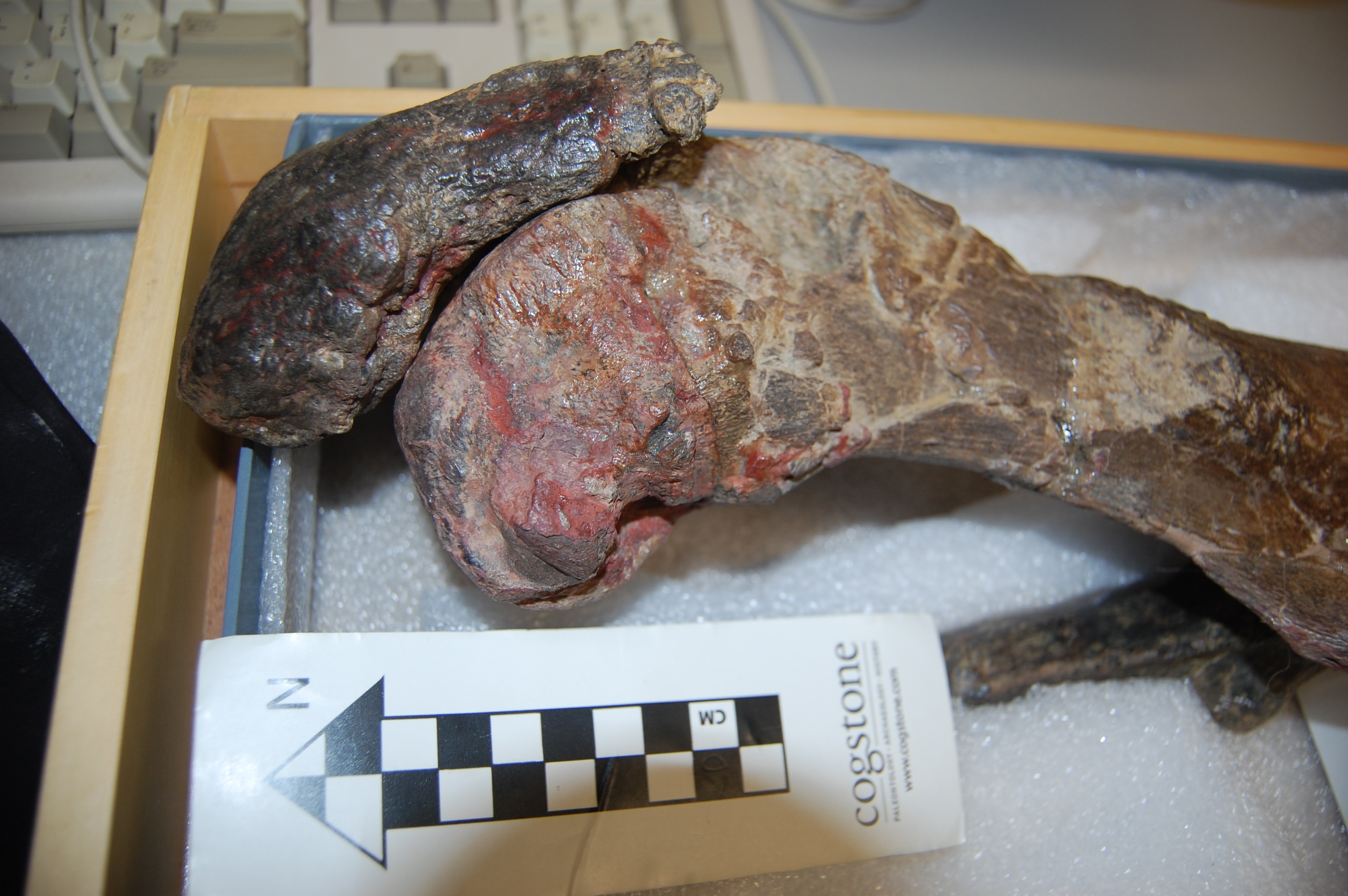
Enough models, let’s have some reality! I warned you this post would get messy, and here it is. Left leg (skinned) of an ostrich showing the muscles around the knee. The patellar region would be in the gloved hand of the lucky individual shown.
This study, for me, was a fun experience in moving toward more fusion of “evo-devo” and biomechanical analyses, a research goal of mine lately– but there’s still a ways to go with the “how?” and “why?” questions even about ostrich kneecaps.
We felt that the best conclusion supported by our analyses was that, rather than have homogeneous stresses and strains throughout their knee tissues (e.g. the patellar tendon), ostriches have a lot of regional diversity in how those tissues are loaded (in the condition we modelled, which is adequately representative of some athletic exertion). Look at the complex FEA coloured results above again, the top two rows: there are a lot of different shades of compression/tension and shear; not homogeneous strains. That diversity of regional loading sets those tissues up for potential transformation throughout growth and development. And thus ONE of the reasons why ostriches might have two kneecaps is that the heterogeneous loading of their knee tendon favours formation of heterogeneous tissue types.
Another, compatible, explanation is that these different tissues might have consequences for how the muscles, tendon and joint operate in movement behaviours. In due time there will be more about that. In the meantime, enjoy the paper if this post makes you want to know more about the amaaaaaazing knees of ostriches!