Stomach-Churning Rating: 10/10 may be too high (it’s all plastinated anatomy; not gooey bloody stuff) but I’m being wary. There are graphic images of humanity and opinions will vary on the tastefulness; I think they are beautiful. (And to me, Bodyworlds plastination leaves specimens looking more like puppets or statues than disturbing undead) There are images of reproductive anatomy that are not appropriate for children unless parental guidance is along for a “birds and the bees” chat. Got it? OK.
I had a spare hour in Cambridge this weekend so I dared the crowds in the revamped UMZC’s upper floor. In my prior visit and post I’d experienced and described the lower floor, which is almost exclusively mammals. This “new” floor has everything else that is zoological (animal/Metazoa) and again is organized in an evolutionary context. And here is my photo tour as promised!
Inviting, soft lighting perfuses the exhibits from the entryway onwards.
All images can be clicked to mu-zoom in on them.
Stomach-Churning Rating: 5/10 for spirit animals, by which I mean dissected/ghostly pale whole specimens of animals in preservative fluids.
The exhibits are on a square balcony overlooking the lower floor, so you can get some nice views. It does make the balcony crowded when the museum is busy, so take that in mind if visiting. Strollers on this upper floor could be really difficult. But the ceiling is very tall so it is not cramped in a 3D sense. The lower floor is more spacious.
Like phylogenies? You got em! Tucked away at the beginning of each major group; not occupying huge valuable space or glaringly obvious like AMNH in NYC but still noticeable and useful. To me, it strikes a good balance; gives the necessary evolutionary context for the displayed specimens/taxa.
Introductory panels explain how names are given to specimens, how specimens are preserved and more.
The exhibits give due focus to research that the UMZC is doing or has been famous for. Hey I recognize that 3D tetrapod image in the lower left! 🙂
There is ample coverage of diversity throughout Metazoa but my camera tended to be drawn to the Vertebrata. Except in some instances like these.
Some larger chelicerates.
Some smaller, shadowy sea scorpion (eurypterid) fossils.
Watch here for more about ophiuroids (brittlestars) in not too long!
A BIG fish brain! Interesting! Before I go through specimens in evolutionary “sequence”, I will feature another thing i really liked: lots of dissected spirit-specimens that show off cool anatomy/evolution/adaptation (and technical skills in anatomical preparation). Mostly heads; mostly fish.
Salps and other tunicates! Our closest non-vertebrate relatives- and some insight into how our head and gut came to be.
Salp-reflection.
Lamprey head: not hard to spot the commonalities with the salps; but now into Vertebrata.
Hagfish head: as a fellow cyclostome/agnathan, much like a lamprey but never forget the slime glands!
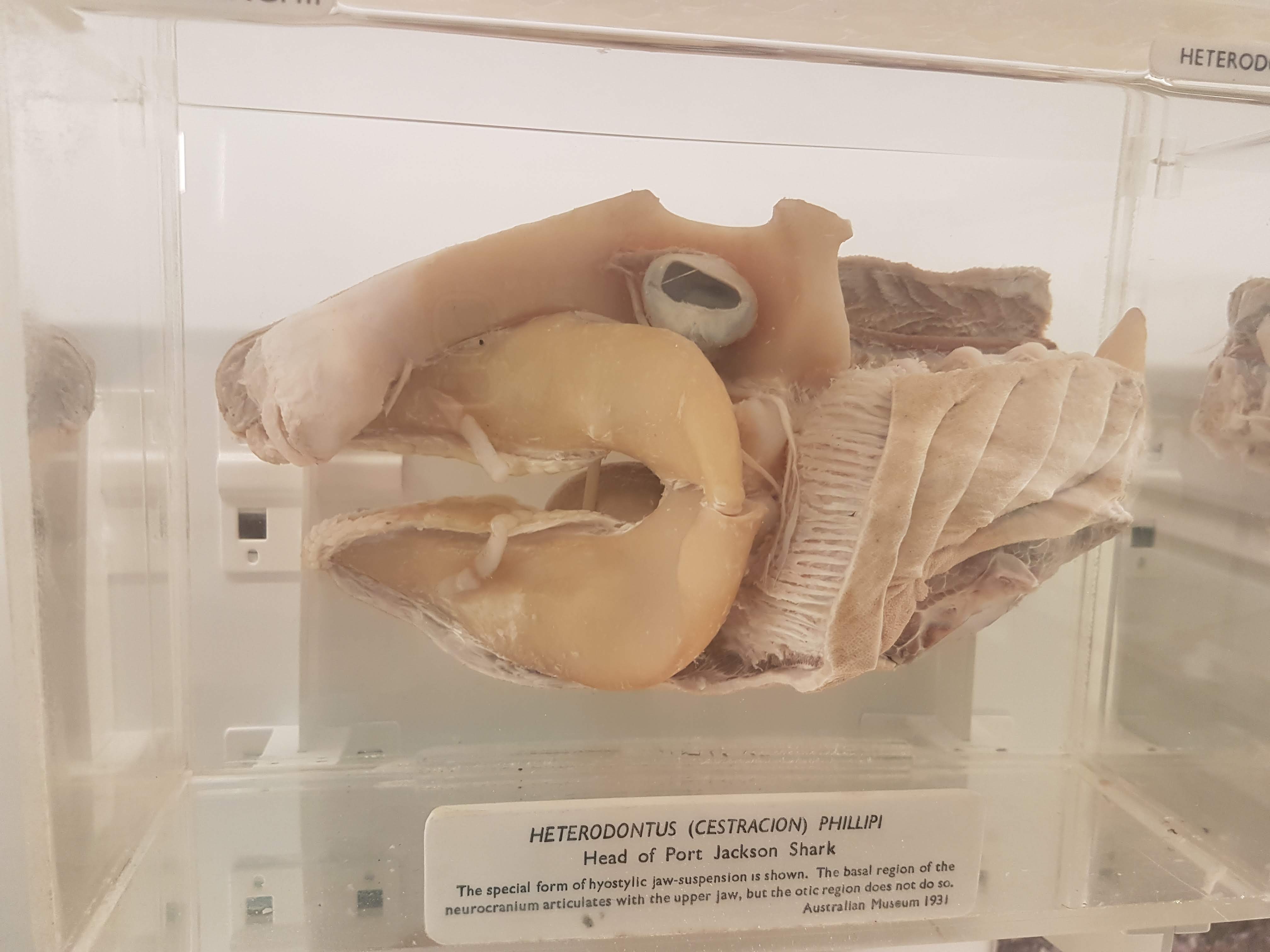
Shark head. Big fat jaws; all the better to bite prey with!
Lungfish (Protopterus) head showing the big crushing tooth plates (above).
Sturgeon vertebrae: tweak some agnathan/shark bits and here you are.
Worm (annelid) anatomy model, displaying some differences from/similarities to Vertebrata. (e.g. ventral vs. dorsal nerve cord; segmentation)
Dissected flipper from a small whale/other cetacean. Still five fingers, but other specializations make it work underwater.
Wonderful diversity of tooth and jaw forms in sharks, rays and relatives. I like this display a lot.
More of the above, but disparate fossil forms!
On with the evolutionary context! Woven throughout the displays of modern animals are numerous fossils, like these lovely placoderms (lineage interposed between agnathans, sharks and other jawed fish).
Goblin shark head.
I seem to always forget what ray-finned fish this is (I want to say wolffish? Quick Googling suggests maybe I am right), but see it often and like its impressive bitey-ness.
Bichir and snakefish; early ray-finned fish radiations.
Armoured and similar fish today.
Armoured fish of the past; some convergent evolution within ray-fins.
Convergence- and homology- of amphibious nature in fish is another evolutionary pattern exemplified here.
Gorgeous fossils of ray-finned fish lineages that arose after the Permian extinctions, then went extinct later in the Triassic.
Note the loooooong snout on this cornetfish but the actual jaws are just at the tip.
Flying fish– those ray-fins are versatile.
Diversity of unusual ray-finned fish, including deep-water and bottom-dwelling forms.
Can you find the low-slung jaws of a dory?
Recent and fossil perch lineage fish.
It’s hard to get far into talking about evolution without bringing up the adaptive radiation of east African cichlid fish, and UMZC researchers are keen on this topic too.
Lobe-fins! Everybody dance!
Rhizodonts & kin: reasons to get out of Devonian-Carboniferous waters.
A Cretaceous fossil coelacanth (skull); not extremely different from living ones’.
Let’s admire some fossil and modern lungfish skulls, shall we? Big platey things (here, mainly looking at the palate) with lots of fusions of tiny bones on the skull roof.
Eusthenopteron fossils aren’t that uncommon but they are still great to see; and very important, because…
OK let’s stop messing around. The UMZC has one of the best displays of fossil stem-tetrapods in the world! And it should.
Another look at the pretty Acanthostega models.
Acanthostega vs. primate forelimb: so like us.
Ichthyostega parts keep Acanthostega company.
A closer look at the “Mr. Magic” Ichthyostega specimen, which takes some unpacking but is incredibly informative and was a mainstay of our 2012 model. Back of skull, left forelimb, and thorax (from left to right here).
Eucritta, another stem-tetrapod.
Closer look at Eucritta‘s skull.
Weird stem-tetrapod Crassigyrinus, which we’re still trying to figure out. It’s a fabulous specimen in terms of completeness, but messy “roadkill” with too many damn bones.
The large skull of Crassigyrinus, in right side view.
Early temnospondyl (true amphibian-line) skulls and neck.
Nectrideans or the boomerangs of the Palaeozoic.
Cool fossil frogs.
Giant Japanese salamander!
Fire salamanders: not as colourful as the real thing, but here revealing their reproductive cycle in beautiful detail.
Closeup of oviduct in above.
Sexual dimorphism in Leptodactylus frogs: the males have bulging upper arms to (I am assuming) help them hold onto females during amplexus (grasping in mating competitions).
Did I forget that Leptodactylus has big flanges on the humerus in males, to support those muscles? Seems so.
An early stem-amniote, Limnoscelis (close to mammals/reptiles divergence); cast.
Grand sea turtle skeleton.
One of my faves on display: a real pareiasaurian reptile skeleton, and you can get a good 3D look around it.
Details on above pareiasaurian.
Mammals are downstairs, but we’re reminded that they fit into tetrapod/amniote evolution nonetheless.
Let there be reptiles! And it was good.
Herps so good. (slow worm, Gila monster, glass lizard)
A curator is Dr Jason Head so you bet Titanoboa is featured!
Crocodylia: impressive specimens chosen here.
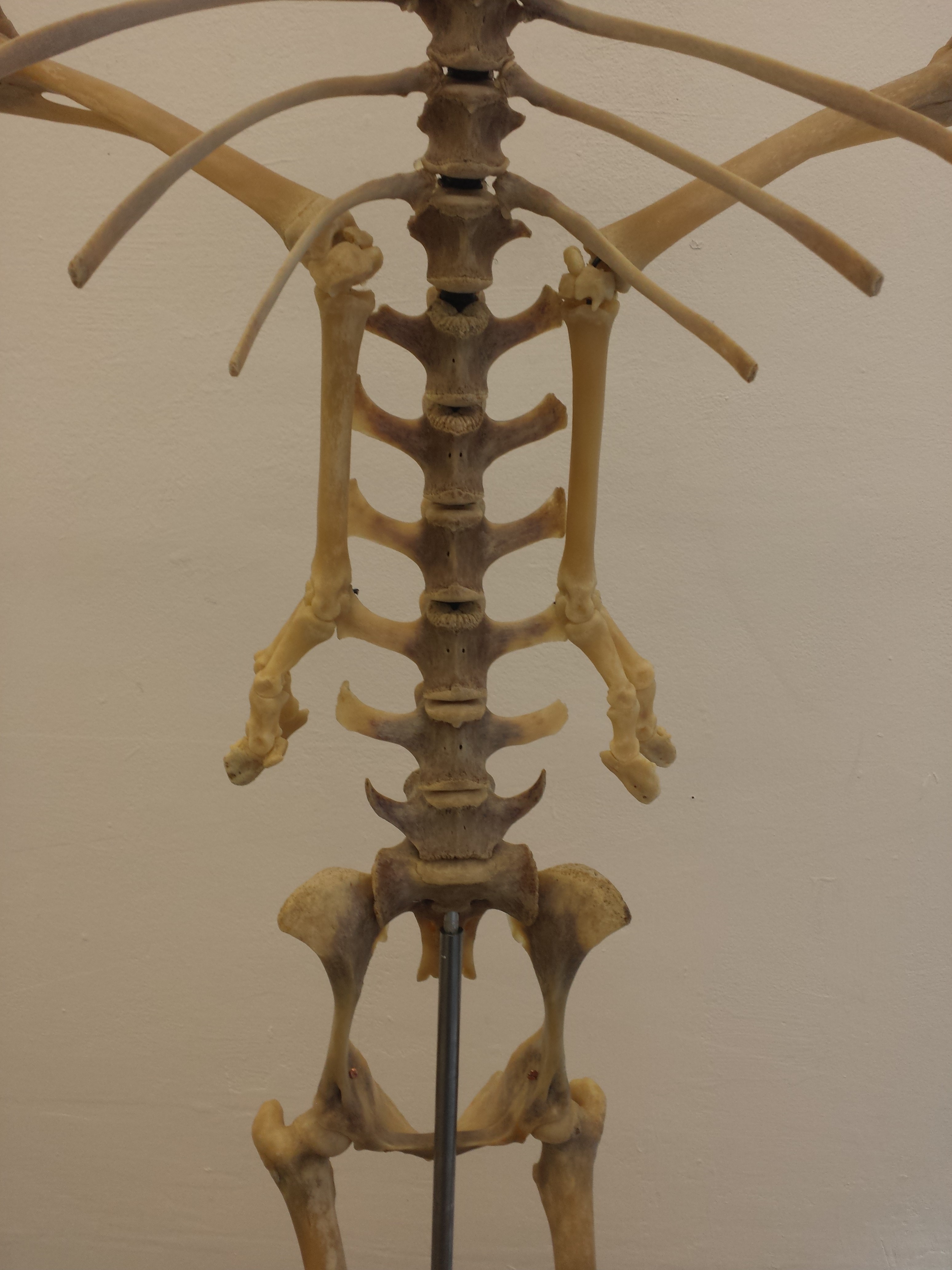
It ain’t a museum without a statuesque ratite skeleton. (There are ~no non-avian dinosaurs here– for those, go to the Sedgwick Museum across the street, which has no shortage!)
Avian diversity takes off.
Glad to see a tinamou make an appearance. They get neglected too often in museums- uncommon and often seemingly unimpressive, but I’m a fan.
I still do not understand hoatzins; the “cuckoo” gone cuckoo.
Dodo parts (and Great Auk) near the entrance.
Wow. What an oilbird taxidermy display! :-O
There we have it. Phew! That’s a lot! And I left out a lot of inverts. This upper floor is stuffed with specimens; easier there because the specimens are smaller on average than on the lower floor. Little text-heavy signage is around. I give a thumbs-up to that– let people revel in the natural glory of what their eyes show them, and give them nuggets of info to leave them wanting more so they go find out.
Now it’s in your hands– go find out yourself how lovely this museum is! I’ve just given a taste.
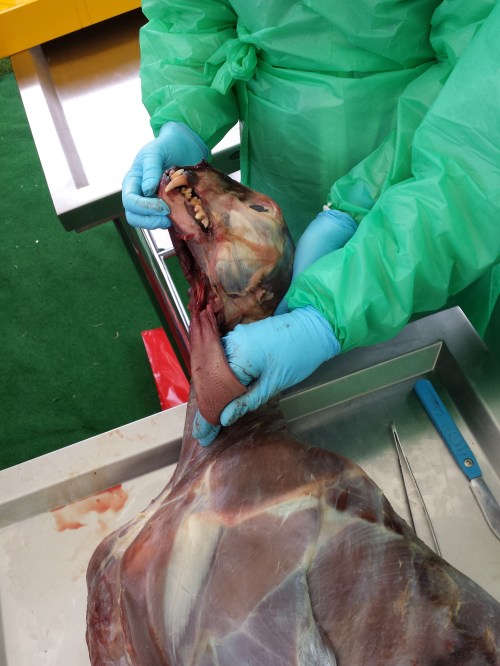
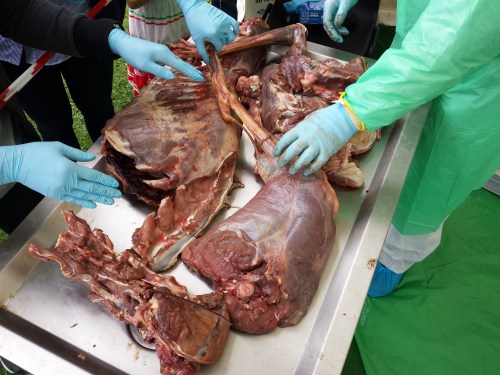
This week we conducted wallaby leg dissections for a study of the kneecaps of marsupials (pouched mammals). Placental (non-pouched) mammals like us almost all have bony kneecaps but many marsupials do not. Kneecaps do important things, acting like gears around the knee joints (e.g. this old post), and yet it is unclear why some marsupials have lost, kept or even re-evolved them as bones. So we’re investigating that and already noticed that one of our wallabies has bony kneecap(s) whereas the other doesn’t, so we’re checking out why and taking tissue samples to do histology (sectioning for microscopic imaging of tissue composition and structure) on so we can see what the knee tendon/kneecap tissues are made of. Some marsupials turn their kneecaps into fibrocartilage rather than bone or tendon and that can be impossible to identify without histology.
The wallabies are small, about 20lbs or so and just take a day or so. Like a turkey. And it’s Thanksgiving today, so here I am with a post about thawing specimens for science, rather than for food. Maybe the title will make sense now.
Stomach-Churning Rating: 7/10; thawed wallaby bits from the get-go.
Thawed lower leg and foot of wallaby. The stickers are for an old study that would take too long to explain…
This post was directly inspired by journalist Jason Bittel’s inquiry to me about my tweet on the wallaby thawing; he wondered if there might be a fun story linking thawing-for-science with thawing-for-Thanksgiving. Some highfalutin editors didn’t agree, so no printed/online story came of this, but I am not so highfalutin, hence this blog post.
Thawed wallaby forelimbs. I’m also looking into the “false thumbs” that some marsupials have (“sixth fingers”), much as elephants and other mammals may have.
Thawing is second nature for our lab’s team; we do it all the time. Avid readers will be unsurprised to learn that just about everything I’ve worked on has been frozen at some time, and thus has been thawed out at some time(s). Normally we don’t freeze if we need live tissue or undistorted tissue, e.g. to measure physiology or very fine microstructure– freezing disrupts all of that. We would instead use physiological saline solution or else a preservative like formalin. And you can only freeze and then thaw a specimen for two times or so before it becomes too useless even for anatomical study.
A small specimen like this salamander can be thawed out simply by running it under warm water for a little while or leaving it out for an hour.
We just leave specimens in a cart, or on a table or sometimes in a cold-room shelving area, for slower thawing. Space heaters tend to overdo things. We don’t do any rough calculation from some sort of thermodynamic first principles of time-to-thaw vs. specimen size (I wish we were that smart!); just seat-of-pants guessing and checking (yes, poking specimens to check their thawedness is a method of choice). Cutting things in half along the way, or skinning them, may be used to accelerate the thawing process. But it’s about as unscientific a method as we use.
The hardest specimens to thaw of course have been the largest specimens. Elephant legs can be >2 metres long and hundreds of kilograms (especially when frozen). A week at room temperature tends to work OK for getting them to a dissectable state. One has to balance the outer deterioration with the inner frigidness. We’re not so concerned about microbe growth in most cases, as one would be with a thawing turkey, and not at all about consumption. We just want to be able to dissect it and make observations, mostly via eyeballing the specimens as we dissect them,
Left hindfoot of an Asian elephant. Still frozen; this was bandthawed- I mean bandsawed- to see its internal anatomy nice and clearly. You may see this specimen again somewhere else– stay tuned! 🙂
Moisture and fluids can be a challenge: generally the rooms we thaw in are low humidity so moisture may not be an issue once the ice melts away, and we have drains nearby. We try to remove ice first or have towels to wipe/soak fluids up as thawing progresses. But if a specimen is sitting in a cart or storage bag with too much ice early on, that can thaw first and then turn the specimen into a nasty slurry of the stuff you’re interested in and the less desirable muck. So we try to avoid that.
De-thawing too early is bad. The smell gets progressively worse– and once the interior of the specimen is thawed enough, then bacteria get in there and the interior becomes a brewing ground for heat production (rather than remaining a cooler region), which accelerates decay, so we don’t want that. We have to check on thawing specimens regularly and move them to cooler storage areas, or begin dissection earlier, if the decay process is noticeably getting excessive.
Any insulation affects thawing time- so scales, feathers, thick skin, shells, fat (for a short while until it decays), and other layers will slow thawing—and may keep heat inside, if there begins to be thawing of the core. So sometimes you open up a specimen that seems dry and clean on the outside and the inside is unpleasant. But with experience that is not hard to avoid.
Thawed wallaby patella prepared for histology.
The foulest specimen I’ve thawed by far was a monitor lizard… it was shipped to me in California from Arizona when I was a PhD student. This was in August’s heat and the box of the big lizard sat thawing at the post office for 2 weeks before they contacted me and asked why a smelly box was bleeding. I came and got it and brought it back to our department but the smell was so bad it set off our building health & safety person’s alarm bells (sorry, David!) and they emailed around a “toxic alert” warning, until I bashfully made it clear that my lizard was the cause, not some toxic chemical. I got in some trouble and was very ashamed. But we put the specimen into a big tank of brine solution and the smell was reduced—the specimen may well still be preserved there 20 years later; I do wonder! Anyway, that experience was so horrendous – and I have a strong stomach—that I regularly recall it and seek to avoid a repeat. It was the most disgusting thing I’ve ever experienced. I do not recommend it.
What we tend to want to get from thawed specimens is: (1) descriptive anatomy (what connects where), and maybe (2) quantitative measurements (laborious metrics of “muscle architecture”– how much does each muscle weigh, how long is it, etc; over and over again for many muscles…). These data not only serve to tell us what makes animals different (and how this evolved) but also the data are used to test questions such as how animals work. In the case of things like wallabies, ultimately we’d love to know what their kneecaps do if they are bony or not; what difference does it make and why might there be differences? We’d spotted one wallaby already that seemed to have a bony kneecap on one leg, and a non-bony one on the other leg, so that asymmetry got us excited.
What’s surprising to learn about thawing animals for science? Well, my first thought is that it’s beautiful. I don’t tend to think of it as gross. I’ve rhapsodized about this before. Animals are wonderful inside and out, and I regularly pause during a dissection to marvel at how amazing the anatomical specializations of animals are. Simple details- shapes, colours, configurations- can be gorgeous. (Often the blood is minimal, drained out early, so that doesn’t detract from or hide the detailed imagery) The gentle yet complex path of a tendon around a joint can yield profound visual enchantment in its elegance. This is all the more true once one ponders how these complex structures evolved, and how much diversity of form and function is out there to study—and how little we know about it! We still don’t know well how to fix many problems humans have with their anatomy, and that’s orders of magnitude worst for most animals, because we don’t understand how anatomy works, or even what the anatomy is like in some cases. So that keeps me busy discovering things. Every specimen is different with surprising little variations, or big ones—sometimes there is one muscle, sometimes it is clearly divided into two muscles, in the same species or even the left vs. right legs. I love seeing those intricacies and wondering about them.
Thawed wallaby shank sliced open to show lovely digital flexors and gastrocnemius muscles. So many questions are raised by this!
If you’re thawing for Thanksgiving, or thawing for science, or thawing out family relations during a gathering, or thawing yourself out from the winter’s cold– my best wishes to you! May we all enjoy what we thaw.
Putting my morphologist hat back on today, I had an opportunity to dissect an Elegant-crested tinamou (Eudromia elegans) for the second time in my life. The last time was during my PhD work ~20 years ago. In today’s dissection I was struck by another reminder of how studying anatomy is a lifelong learning experience and sometimes it’s really fun and amazing even when it’s stinky.
Tinamou foot. I did know that tinamous don’t have a hallux; big “perching toe” (1st/”big toe” in us); true of ratites/palaeognaths more generally. Unlike a chicken or many other birds. Just the three main toes (2, 3 and 4) are here.
Stomach-Churning Rating: 7/10; you gotta have guts to learn about intestine-churning stuff.
Tinamous are neat little partridge-like ground birds but they are not close cousins of partridges or guineafowl at all. Their closest cousins are other ratites/palaeognaths such as ostriches and kiwis. And hence they are found in South America, especially Patagonia in Argentina. I’ve seen them there, much to my enjoyment.
Said tinamou.
What struck me today was that, as I delved into the digestive system of this bird, I saw features that were unfamiliar to me even after having dissected many species of birds from many lineages. The intestinal region was very lumpy, with little bud-like pockets full of dense droppings. Furthermore, on separating the tubes of the small and large intestines I realized that most of the intestinal volume itself was caecum (normally a modest side-pocket near the juncture of the small and large intestines). Indeed, that caecum was caeca (plural): it had two massive horns; it was a double-caecum, feeding back into the short rectum and cloaca. Birds have variable caeca and it is typical to see subdivision into two parts, but I’d never seen it to this degree.
Oh why not, here’s the gizzard/stomach showing its grinding pebbles and bits of food, plus the strong outer muscle layers (pink) for driving that grinding. Small intestine heads toward the bottom of the image. Yes, we do need a better dissection light…
I had to question my anatomical knowledge at this point, wondering if I was identifying things incorrectly—did I really screw up somehow and these were other organs, like giant ovaries? But no, they were clearly full of faecal matter; they were digestive organs. I finished the dissection, still puzzled, and hit the literature. Right away, Google-Scholaring for “tinamou caecum” I found the answer, here (free pdf link):
“at least one species (Elegant Crested Tinamou, [Eudromia elegans]), the ceca contain multiple sacculations, resulting in structures that look much like two bunches of fused grapes.”
The caeca in question.
OK buddy, those are the little lumpy buds I saw. Bunches of grapes—exactly.
And later:
“The paired ceca of the Elegant Crested Tinamou are extraordinary and probably unique within Aves (Fig. 3): long and wide (12.5-13.0 X 2.2- 2.5 cm; Wetmore 1926) and internally honeycombed by many small diverticula. These outpocketings gradually diminish in size and organization from the base to the tip of the organ, apically showing a more spiral form of internal ridges like ratite ceca. Externally, the basal diverticula protrude from the ceca as pointed lobes, gradually becoming flatter but still clearly apparent toward the organ’ s tip.”
Whoa! I never knew that! So I happened to be dissecting a bird, fairly common in its homeland, that has a really bizarre and singular form of caeca/ceca! That hit my morphologist sweet-spot so I was very pleased and decided to share with you. It is one of those many examples of times when you quickly go from confusion to illumination as a scientist, emerging with a neat fact about animal biology. And journal articles help you get there!
The bare “brood patch” on the back end of the tinamou’s belly; a nicely hotspot for keeping eggs warm. Perhaps for brooding bad puns, too.
Who needs “Ice Road Truckers” when you have the “John’s Freezer” team on the road with fossils, amphibians, felids and 3D phenotype fun? No one, that’s who. We’re rocking the Cheltenham Science Festival for our first time (as a group), and pulling out all the stops by presenting two events! Here’s the skinny on them, with updates as the week proceeds.
Stomach-Churning Rating: 2/10 for now (just bones), but it could change once the cheetah dissection is under way… 8/10 bloody cheetah bits but only at the end (updated)
Right now, Lauren Sumner-Rooney (of “Anatomy To You” and other fame) is on-site with a rotating team of others from our lab, in the “Free Activity Tents” area of the Imperial Gardens/Square, inside a marquee where we’ll be showing off our NERC-funded tetrapod research all week. This “First Steps” event features not only our past and present work with Jenny Clack, Stephanie Pierce, Julia Molnar and others on Ichthyostega & its “fishapod” mates, but also our “scampering salamanders” research in Spain, Germany and England. I’ve blogged a lot about all that, and won’t repeat it here, but you can see a 3D-printed Ichthyostega skeleton, view the skeleton in a virtual reality 3D environment, see related specimens and engage in kid-friendly activities, and talk to our team about this and other related research.
Ichthyostega 3D printed backbone is born!
The central themes of that event are how bone structure relates to function and how we can use such information, along with experimental measurements and computer models of real salamanders, to reconstruct how extinct animals might have behaved as well as how swimming animals became walking ones. How did fins transform into limbs and what did that mean for how vertebrates made the evolutionary transition onto land? If you know my team’s work, that encapsulates our general approach to many other problems in evolutionary biomechanics (e.g. how did avian bipedalism evolve?). Added benefits are that you too can explore this theme in a hands-on way, and you can talk with us about it in person. That continues all week (i.e. until Saturday evening); I’ll be around from Thursday afternoon onwards, too. Kids of all ages are welcome!
Ichthyostega 3D print taking shape!
Then, on Saturday for our second free event we join forces with Ben Garrod (master of primate evolution, the secrets of bones, and “Attenborough and the Giant Dinosaur”) and RVC’s forensic pathologist Alexander Stoll as well as Sophie Regnault (“sesamoid street” PhD student w/me). As the “Large Animal Dissection” title hints, it’s not the right kind of gig to bring small kids to. There will be blood and stuff— we’ll be dissecting a cheetah together from 10am-4pm. This will involve walking through all the major organ systems, giving evolutionary anecdotes, and plenty more, with an aim to understand how the magnificent adaptations of cheetahs evolved—but also to investigate what problem(s) this animal faced that led to its sad demise. By the day’s end, there will just be a skeleton left. Get a front row seat early for this event, which serendipitously ties into “Team Cat”’s Leverhulme Trust-funded research project (we wanted a big animal and it just happened to be a cheetah; I had hoped for a giant croc or a shark or something but can’t complain!).
Ichthyostega 3D print is ready!
If you miss these events, please do cry bitter tears of regret. But don’t despair, there will be another “big cat dissection” in the London area in ~November (watch here for details), and plenty more fossil tetrapod stuff to come, plus a LOT more dinosaurs on the horizon!
Guess the bones! (photo by Zoe Self)
And please come back to this blog post for more pics and stories as the week carries on… For hashtag afficionados, you can follow the fun on Twitter etc. at #firststepsCSF16. What a world we live in!
Update 1: While you’re here, check out our Youtube playlists of tetrapod-related videos:
Update 2: Photos of our main stand (about tetrapod evolution)
Our poster/banner display looks nice.
Our tent brings in some punters.
Our bones excite people young and old, sighted and blind.
Fun with stickers and lab t-shirts.
And…
Update 3: Cheetah meat & greet
Ben, Alex, Sophie and I tackled the cheetah dissection today and it went GREAT! Much better than I’d optimistically expected. Rain didn’t scare the crowds off and neither did the gore, which there was some of (gelatinous spinal cords, lumpy tumors and at least one flying tiny bit of cheetah flesh that landed on a good-natured audience member!). Photos will tell the tale:
Peek-a-boo!
Sophie and Alex help us get set up in our tent.
Our initial rough schedule- although we ended up improvising more after lunch.
Dissectors assemble!
The beast revealed. It was skinned by the museum that loaned it to us.
Alex showing his talent: removing the viscera in one piece from end to end, starting with the tongue.
Impressive finding of a surgical fixture (plate and wires) on the tibia, which had been used to hold the shattered bone back together long enough for it to heal. Added to the kidney disease and liver-spleen-lung cancer, this cheetah was in the sorriest shape of any cadaver I’ve seen yet.
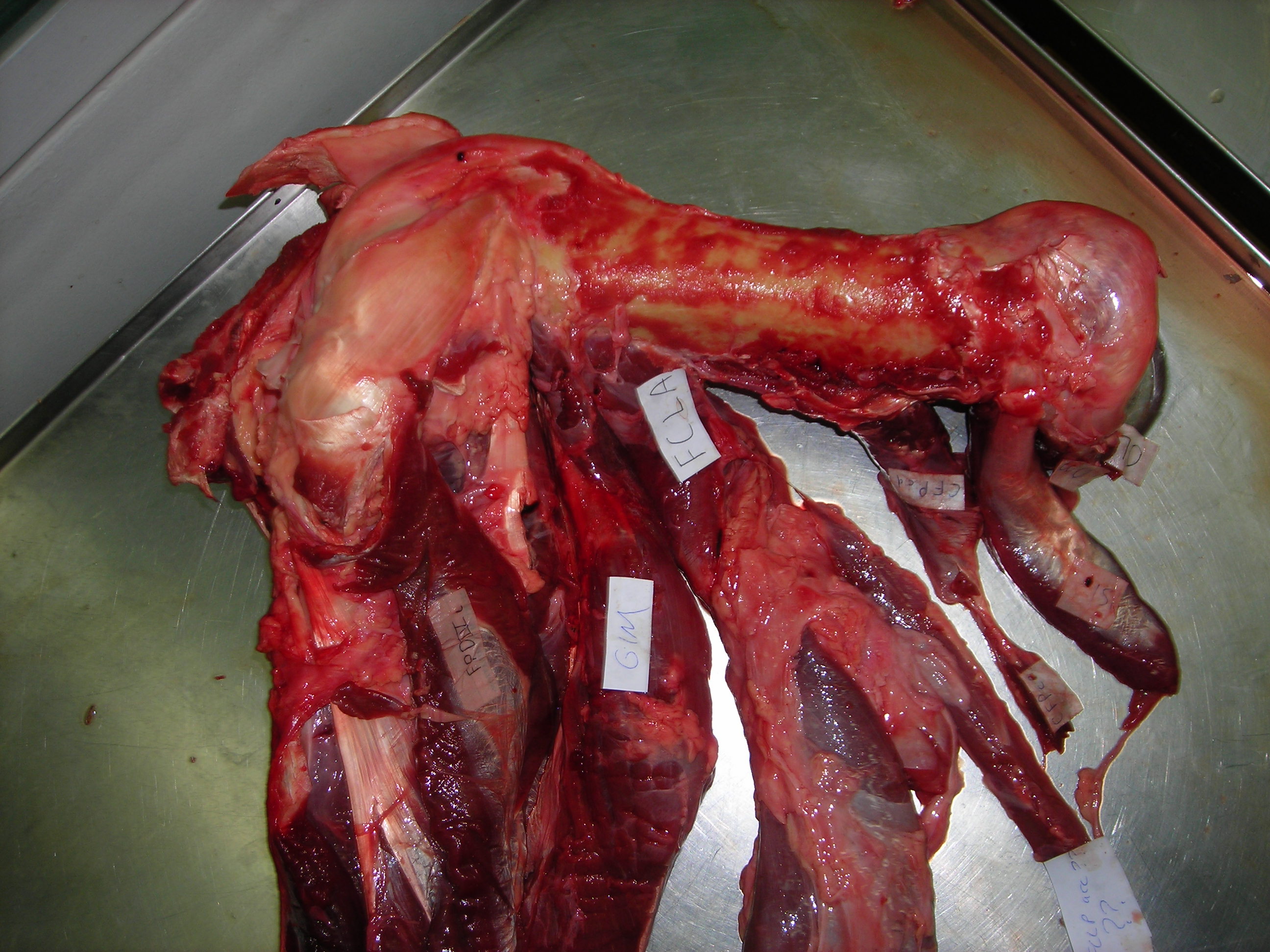
Cheetah coming to pieces: (from bottom) lumbar/pelvic region, hindlimb, thorax, forelimb and other bits.
Dr Adam Rutherford, an eye expert, did a nice dissection of the cheetah’s eye, here showing the tapetum lucidum (reflective membrane), which shows up as light blue colour. Its small size befits the not-very-nocturnal habits of cheetahs.
The lens of the cheetah’s eye. Now cloudy because of dehydration and crystalization, but still fascinating to see.
Happy Darwin Day from the frozen tundra sunny but muddy, frosty lands of England! I bring you limb muscles as peace offerings on this auspicious day. Lots of limb muscles. And a new theme for future blog posts to follow up on: starting off my “Better Know A Muscle” (nod to Stephen Colbert; alternative link) series. My BKAM series intends to walk through the evolutionary history of the coolest (skeletal/striated) muscles. Chuck Darwin would not enjoy the inevitable blood in this photo-tour, but hopefully he’d like the evolution. Off we go, in search of better knowledge via an evolutionary perspective!
There is, inarguably, no cooler muscle than M. caudofemoralis longus, or CFL for short. It includes the largest limb muscles of any land animal, and it’s a strange muscle that confused anatomists for many years– was it a muscle of the body (an axial or “extrinsic” limb muscle, directly related to the segmented vertebral column) or of the limbs (an “abaxial” muscle, developing with the other limb muscles from specific regions of the paraxial mesoderm/myotome, not branching off from the axial muscles)? Developmental biologists and anatomists answered that conclusively over the past century: the CFL is a limb muscle, not some muscle that lost its way from the vertebral column and ended up stranded on the hindlimb.
The CFL is also a muscle that we know a fair amount about in terms of its fossil record and function, as you may know if you’re a dinosaur fan, and as I will quickly review later. We know enough about it that we can even dare to speculate if organisms on other planets would have it. Well, sort of…
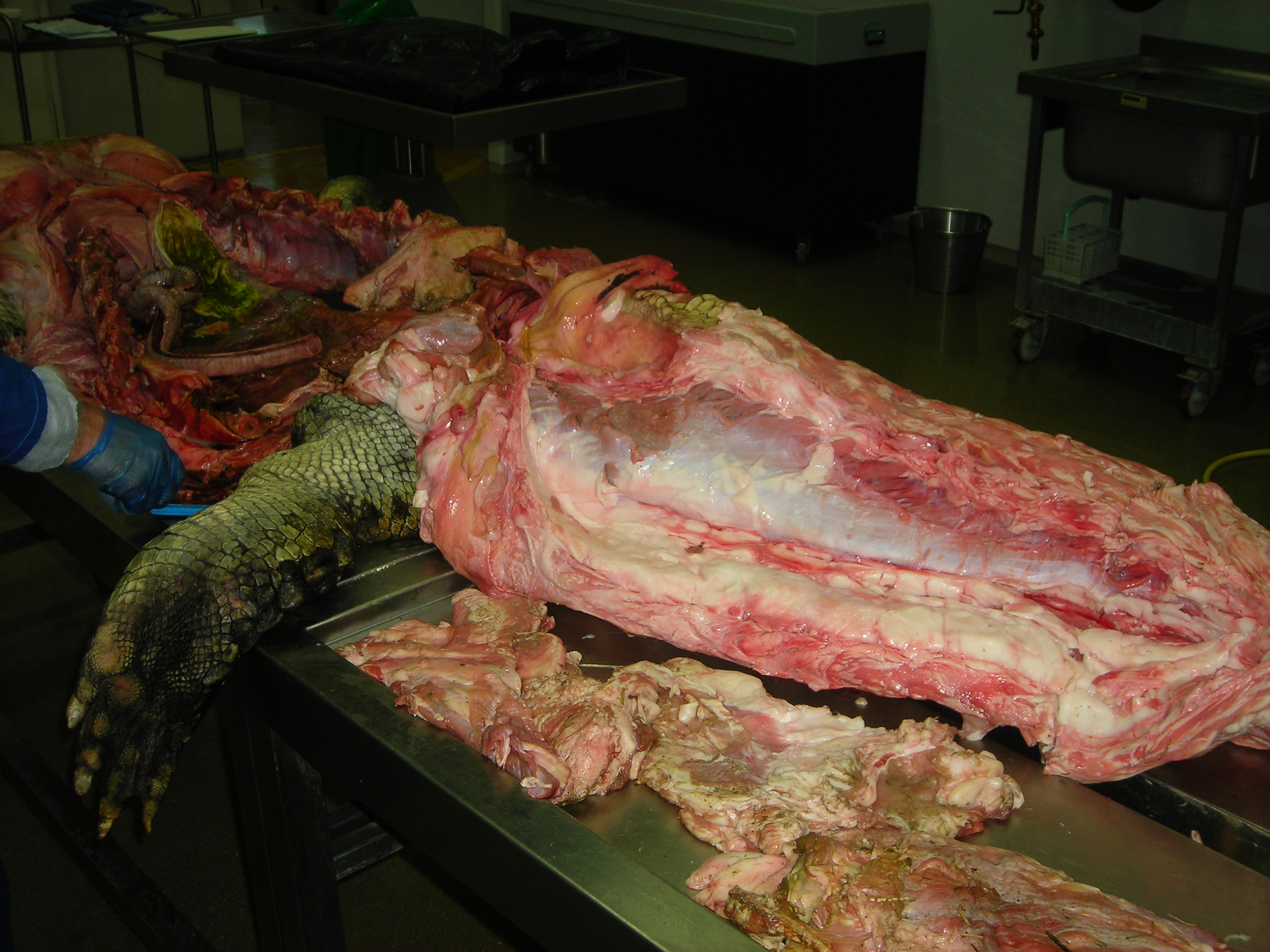
Stomach-Churning Rating: 8/10. Lots of meaty, bloody, gooey goodness, on and on, for numerous species. This is an anatomy post for those with an appetite for raw morphology.
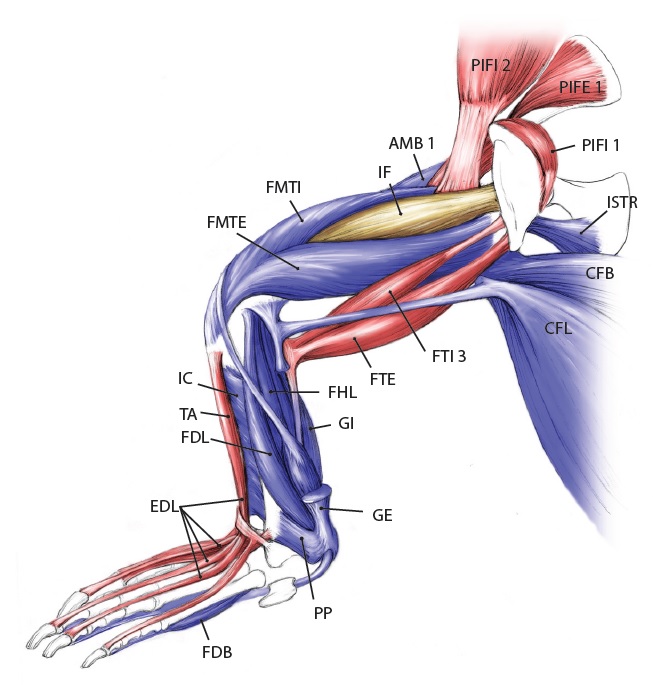
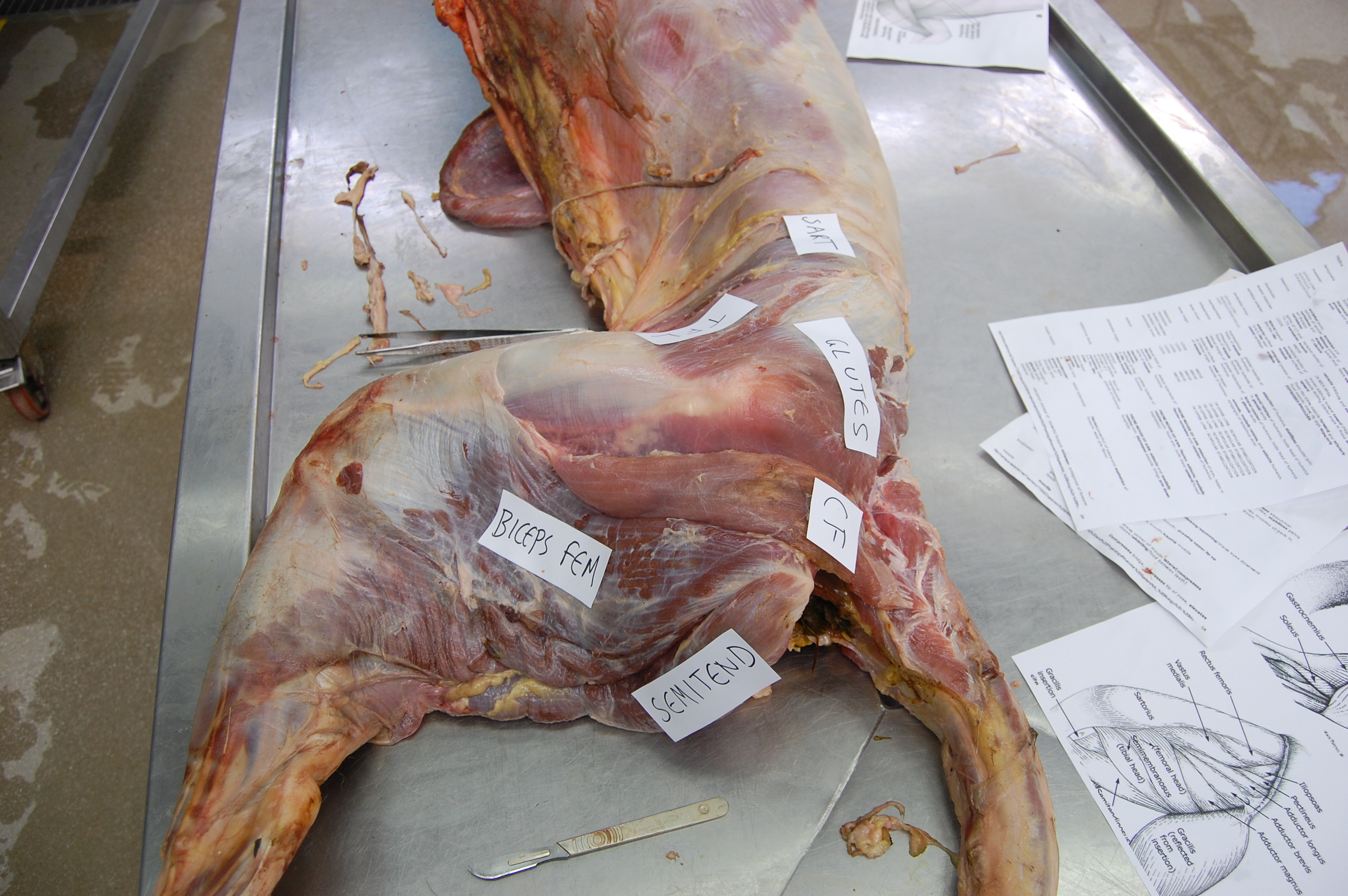
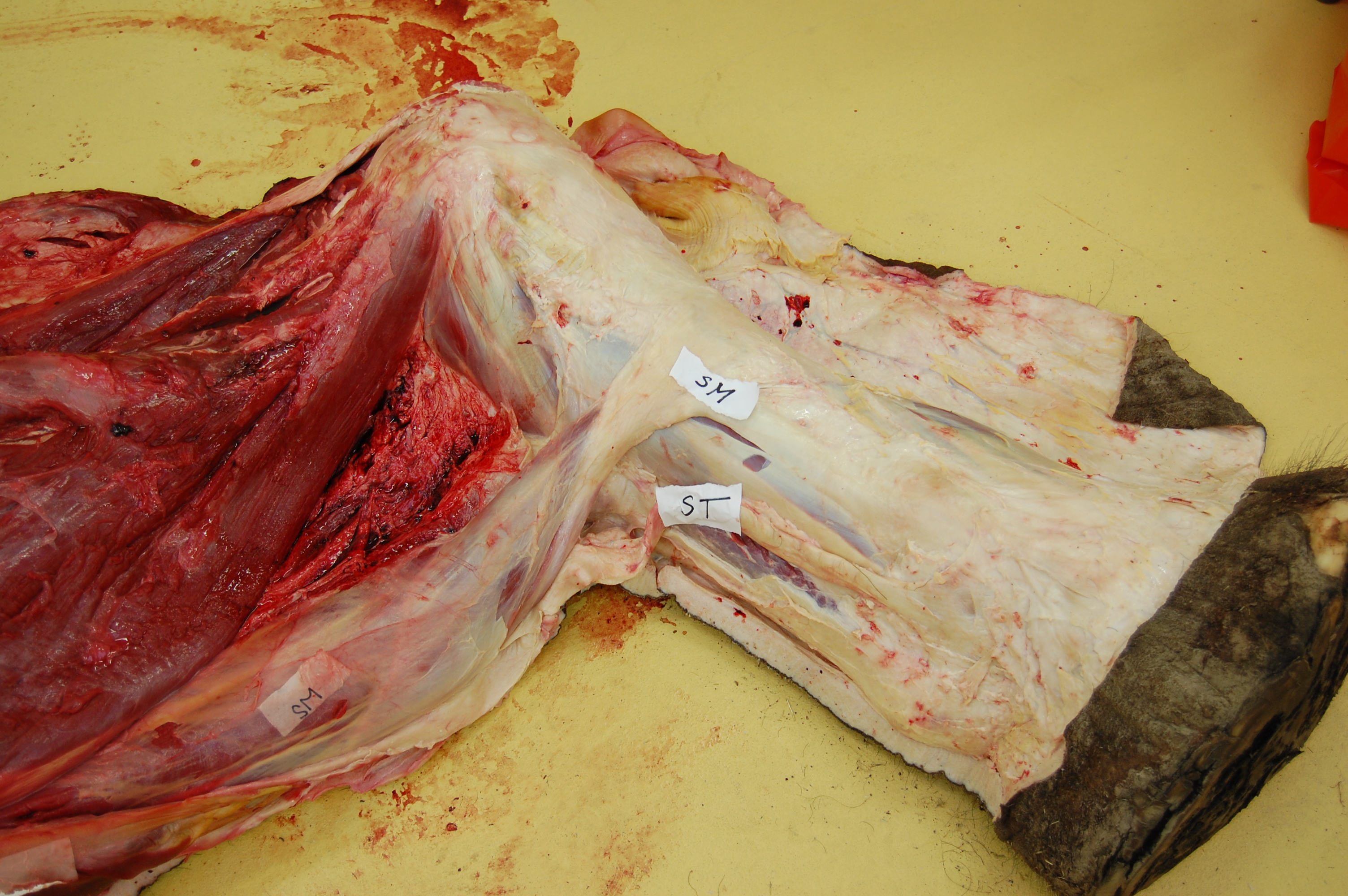
Let’s start from a strong (and non-gooey) vantage point, to which we shall return. The CFL in crocodiles and most other groups is (and long was) a large muscle extending from much of the front half or so of the tail to the back of the femur (thigh bone), as shown here:
Julia Molnar’s fabulous illustration of Alligator‘s limb muscles, from our 2014 paper in Journal of Anatomy. Note the CFL in blue at the bottom right.
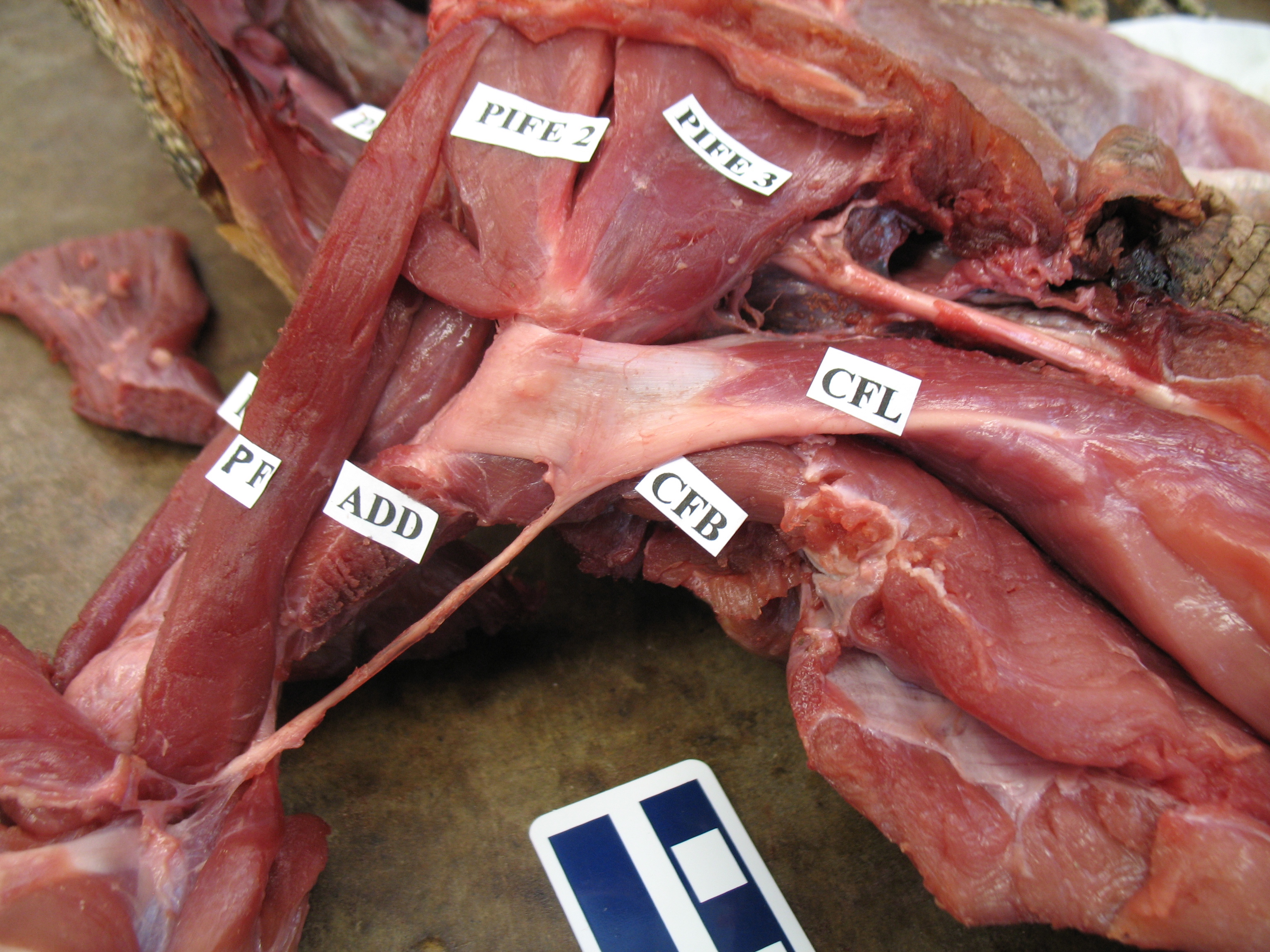
As the drawing shows, the CFL has a friend: the CFB. The CFB is a shorter, stumpier version of the CFL restricted to the tail’s base, near the hip. The “B” in its name means “brevis”, or runty. It gets much less respect than its friend the CFL. Pity the poor CFB.
But look closer at the CFL in the drawing above and you’ll see a thin blue tendon extending past the knee to the outer side of the lower leg. This is the famed(?) “tendon of Sutton“, or secondary tendon of the CFL. So the CFL has two insertions, one on the femur and one (indirectly) onto the shank. More about that later.
Together, we can talk about these two muscles (CFL and CFB) as the caudofemoralis (CF) group, and the name is nice because it describes how they run from the tail (“caudo”) to the femur (“femoralis”). Mammal anatomists were late to this party and gave mammal muscles stupidly unhelpful names like “gluteus” or “vastus” or “babalooey”. Thanks.
But enough abstract drawings, even if they rock, and enough nomenclature. Here is the whopping big CFL muscle of a real crocodile:
Huge Nile crocodile, but a relatively small CFL.
Bigger crocs have smaller legs and thus smaller leg muscles, relatively speaking. CFL at the top, curving to the left.
The giant Nile croc’s CFL muscle removed for measurements. 2.35 kg of muscle! Not shabby for a 278 kg animal.
However, maybe crocodile and other archosaur CFL muscles are not “average” for leggy vertebrates? We can’t tell unless we take an evolutionary tack to the question.
Where did the CFL come from, you may ask? Ahh, that is shrouded in the fin-limb transition‘s mysteries. Living amphibians such as salamanders have at least one CF muscle, so a clear predecessor to the CFL (and maybe CFB) was present before reptiles scampered onto the scene.
But going further back through the CF muscles’ history, into lobe-finned fish, becomes very hard because those fish (today) have so few fin muscles that, in our distant fishy ancestors, would have given rise eventually to the CF and other muscle groups. With many land animals having 30+ hindlimb muscles, and fish having 2-8 or so, there obviously was an increase in the number of muscles as limbs evolved from fins. And because a limb has to do lots of difficult three-dimensional things on land while coping with gravity, more muscles to enable that complex control surely were needed.
OK, so there were CF muscles early in tetrapod history, presumably, anchored on that big, round fleshy tail that they evolved from their thin, finned fishy one — but what happened next? Lizards give us some clues, and their CFL muscles aren’t all that different from crocodiles, so the CFL’s massive size and secondary “tendon of Sutton” seems to be a reptile thing, at least.
Courtesy of Emma Schachner, a large varanid lizard’s very freshly preserved CFL and other hindlimb muscles.
Courtesy of Emma Schachner, zoomed in on the tendons and insertions of the CFL muscle and others. Beautiful anatomy there!
Looking up at the belly of a basilisk lizard and its dissected right leg, with the end of the CFL labelled. It’s not ideally dissected here, but it is present.
An unspecified iguanid(?) lizard, probably a juvenile Iguana iguana, dissected to reveal its CFL muscle near its attachment to the femur. The muscle would extend further, about halfway down the tail, though.
Let’s return to crocodiles, for one because they are so flippin’ cool, and for another because they give a segue into archosaurs, especially dinosaurs, and thence birds:
A moderate-sized (45kg) Nile crocodile with its CFL muscle proudly displayed. Note the healthy sheath of fat (cut here) around the CFL.
American alligator’s CFL dominates the photo [by Vivian Allen].
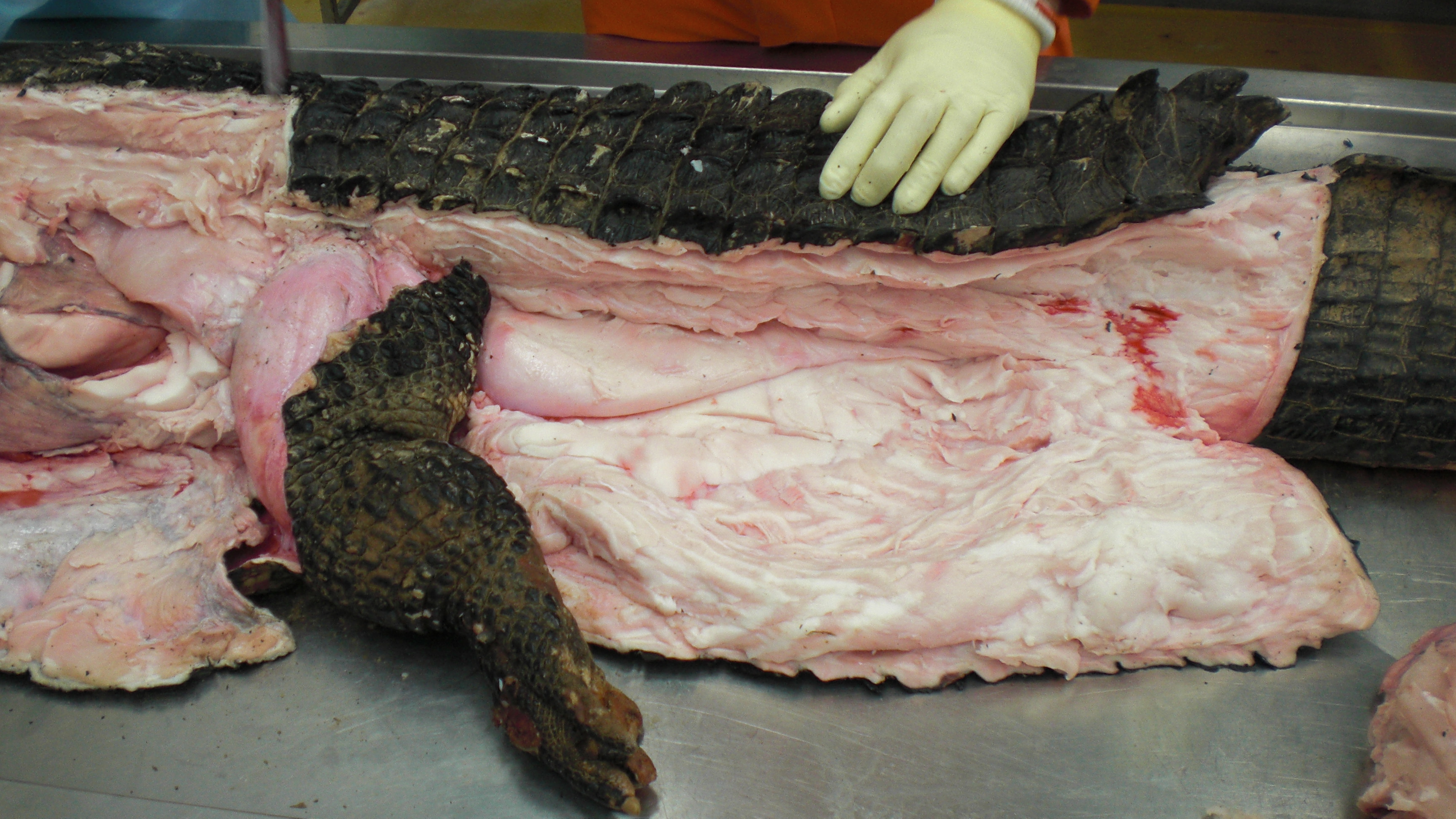
Black caiman, Melanosuchus, showing off its CFL muscle (pink “steak” in the middle of the tail near the leg), underneath all that dark armour and fatty superficial musculature.
A closer look at the black caiman’s thigh and CFL muscle.
Like I hinted above, crocodiles (and the anatomy of the CFL they share with lizards and some other tetrapods) open a window into the evolution of unusual tail-to-thigh muscles and locomotor behaviours in tetrapod vertebrates.
Thanks in large part to Steve Gatesy’s groundbreaking work in the 1990s on the CFL muscle, we understand now how it works in living reptiles like crocodiles. It mainly serves to retract the femur (extend the hip joint), drawing the leg backwards. This also helps support the weight of the animal while the foot is on the ground, and power the animal forwards. So we call the CFL a “stance phase muscle”, referring to how it mainly plays a role during ground contact and resisting gravity, rather than swinging the leg forwards (protracting the limb; i.e. as a “swing phase muscle”).
The “tendon of Sutton” probably helps to begin retracting the shank once the thigh has moved forward enough, facilitating the switch from stance to swing phase, but someone really needs to study that question more someday.
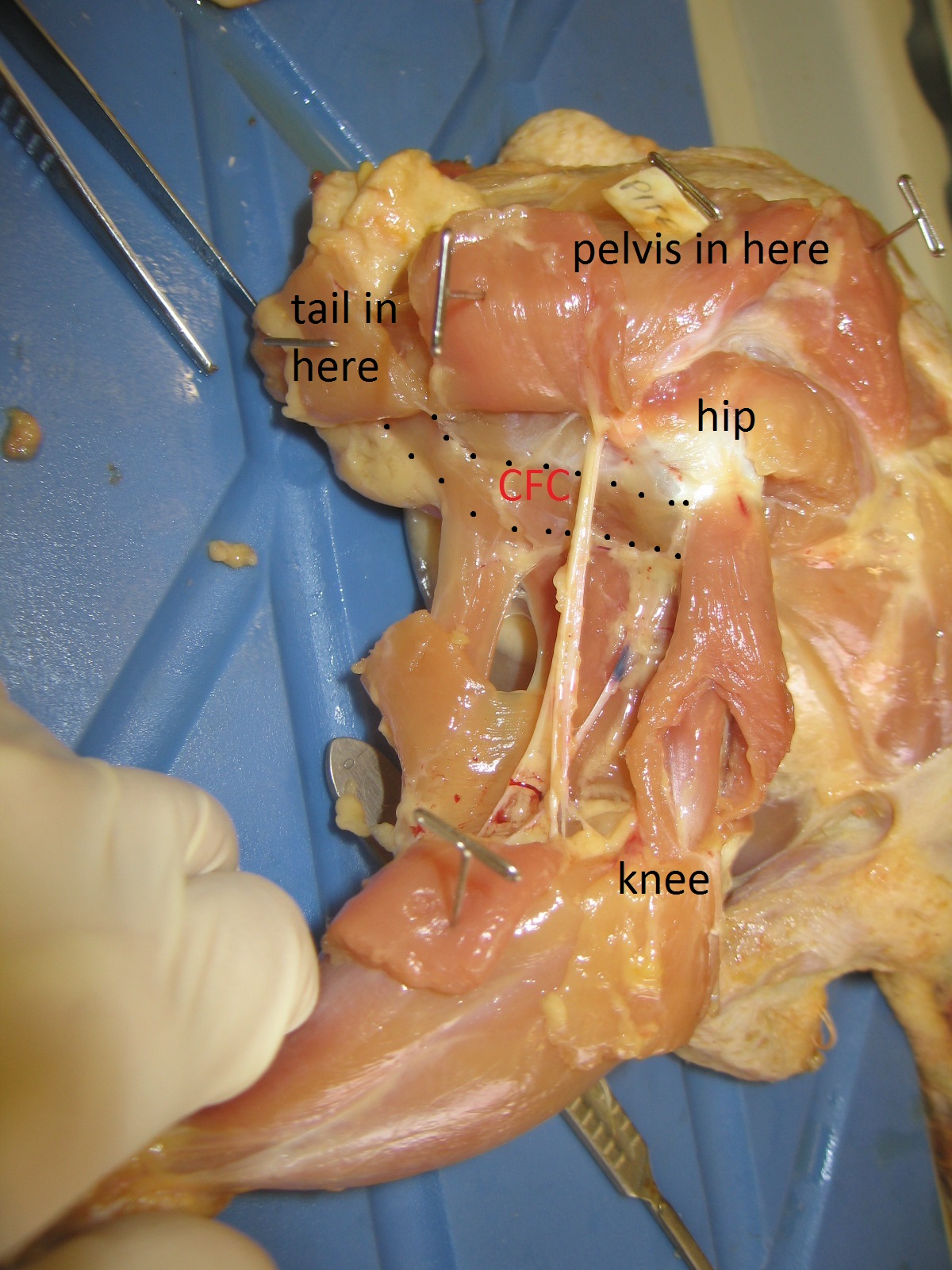
And thanks again to that same body of work by Gatesy (and some others too), we also understand how the CFL’s anatomy relates to the underlying anatomy of the skeleton. There is a large space for the CFL to originate from on the bottom of the tail vertebrae, and a honking big crest (the fourth trochanter) on the femur in most reptiles that serves as the major attachment point, from which the thin “tendon of Sutton” extends down past the knee.
Femur bones (left side; rear view) from an adult ostrich (left) and Nile crocodile (right). Appropriate scale bar is appropriate. The fourth trochanter for the CFL is visible in the crocodile almost midway down the femur. Little is left of it in the ostrich but there is a bumpy little muscle scar in almost the same region as the fourth trochanter, and this is where the same muscle (often called the CFC; but it is basically just a small CFL) attaches.
That relationship of the CFL’s muscular anatomy and the underlying skeleton’s anatomy helps us a lot! Now we can begin to look at extinct relatives of crocodiles; members of the archosaur group that includes dinosaurs (which today we consider to include birds, too), and things get even more interesting! The “tendon of Sutton”, hinted at by a “pendant” part of the fourth trochanter that points down toward the knee, seems to go away multiple times within dinosaurs. Bye bye! Then plenty more happens:
A large duckbill dinosaur’s left leg, with a red line drawn in showing roughly where the CFL would be running, to end up at the fourth trochanter. Many Mesozoic dinosaurs have skeletal anatomy that indicates a similar CFL muscle.
We can even go so far as to reconstruct the 3D anatomy of the CFL in a dinosaur such as T. rex (“Sue” specimen here; from Julia Molnar’s awesome illustration as part of our 2011 paper), with a fair degree of confidence. >180kg steak, anyone?
As we approach birds along the dinosaur lineage, the tail gets smaller and so does the fourth trochanter and thus so must the CFL muscle, until we’re left with just a little flap of muscle, at best. In concert, the hindlimbs get more crouched, the forelimbs get larger, flight evolves and voila! An explosion of modern bird species!
Left femur of an ostrich in side view (hip is toward the right side) showing many muscles that attach around the knee (on the left), then the thin strap of CF muscle (barely visible; 2nd from the right) clinging near the midshaft of the femur.
Another adult ostrich’s CF muscle complex, removed for study. Not enough ostrich myology for you yet? Plenty more in this old post!Or this one!Or this one… hey maybe I need to write less about ostriches? The CF muscle complex looks beefy but it’s no bigger than any other of the main hindlimb muscles, unlike the CFL in a crocodile or lizard, which puts everything else to shame!
STILL not enough ostrich for you yet? Take a tour of the major hindlimb muscles in this video:
And check out the limited mobility of the hip joint/femur here. No need for much femur motion when you’re not using your hip muscles as much to drive you forwards:
But I must move on… to the remainder of avian diversity! In just a few photos… Although the CF muscles are lost in numerous bird species, they tend to hang around and just remain a long, thin, unprepossessing muscle:
Chicken’s right leg in side view. CFC muscle (equivalent of CFL; the ancestral CFB is confusingly called the CFP in birds, as it entirely resides on the pelvis) outlined and labelled.
A jay (species? I forget) dissected to show some of the major leg muscles, including the CFL-equivalent muscle; again, smallish. [Photo by Vivian Allen]
Finally, what’s up with mammals‘ tail-to-thigh CF-y muscles? Not much. Again, as in birds: smaller tail and/or femur, smaller CF muscles. Mammals instead depend more on their hamstring and gluteal muscles to support and propel themselves forward.
But many mammals do still have something that is either called the M. caudofemoralis or is likely the same thing, albeit almost always fairly modest in size. This evolutionary reduction of the CF muscle along the mammal (synapsid) lineage hasn’t gotten nearly as much attention as that given to the dinosaur/bird lineage’s CFL. Somebody should give it a thoroughly modern phylogenetic what-for! Science the shit outta that caudofemoralis…
Yet, oddly, to give one apparent counter-example, cats (felids) have, probably secondarily, beefed up their CF muscle a bit:
Cats have a pretty large CF muscle in general, and this jaguar is no exception! But mammals still tend to have fairly wimpy tails and thus CF muscles, or they even lose them (e.g. us?). [photo by Andrew Cuff, I think]
In summary, here’s what happened (click to embeefen):
Better Know A Muscle: the evolution of M. caudofemoralis (longus).
I hope you enjoyed the first BKAM episode!
I am willing to hear requests for future ones… M. pectoralis (major/profundus) is a serious contender.
P.S. It was Freezermas this week! I forgot to mention that. But this post counts as my Freezermas post for 2016; it’s all I can manage. Old Freezermas posts are here.
Tonight is the debut of the ballyhooed BBC1 programme “Attenborough and the Giant Dinosaur“, featuring Sir David and The Titanosaur-With-No-Name, at 6:30pm. Furthermore, this week I presided over a showing of “T. rex Autopsy” to our RVC undergrad Zoological Society, with a very enjoyable Q&A afterwards. So it seemed timely for me to do a post about a theme that links these two documentaries that I helped with, my own research, and science communication and palaeontological research more generally. But first let’s get this out of the way:
It was great. I could gush more but that’s not what this post is about.
Stomach-Churning Rating: ~7/10; mainly the elephant leg dissection that’s not far below, which is a bit messy.
For the titanosaur documentary with Sir David, and the incomparable Ben Garrod as well, we had an old elephant “friend” of mine (subject of many biomechanics studies we’d done) walk across a pressure pad to demonstrate how the elephant locomotion works and some of the basic similarities with how a giant sauropod dinosaur might walk:
A broader feature of that documentary, which elephants are linked into, is how we can use the skeleton to reconstruct some general aspects of the soft tissue anatomy, and thereby the physiology or even behaviour, of a giant titanosaur. Which brings me to this post’s subject: We dig up dinosaurs all the time, but what about digging into dinosaurs and using what’s preserved to reconstruct what isn’t?
Some of the “G-suit” compressive stocking that the fascia wrapped around elephant, and other large mammals, creates, and the underlying, interwoven muscles and tendons (hindlimb of a young Asian elephant that sadly died in captivity). Did some larger dinosaurs have something like this? I’d expect so.
Another view, more superficially, of that G-suit/stocking under the thick, tight skin of an elephant’s leg. You’ll hear more about this in the Attenborough show…
Once the documentary airs, I may be able to share some more images from it showing what they did for the titanosaur, but this BBC photo gives a good idea. Here, blood vessels and other tissues surrounding the skeleton. How would a titanosaur pump blood around its body? A good question.
I wanted to focus more on the end product; the imagery or even physical object; and how it conveys what we think we know about dinosaur anatomy. I’ll do that via examples, using photos of dinosaur anatomy that I’ve collected over the years from museums or other media. There won’t be any profound points or long musings; it’s mainly a photo blog:
What your (inner?) child most needs is a dinosaur to dissect yourself! Why not a T. rex toy like this?
For the price (~$30 in USA), the 4D Vision dinosaurs deliver a pretty good bargain, and the anatomy is satisfactory. I’ve been collecting this series. I could quibble, but hey: it’s a dinosaur you get to build/dissect yourself, and with many major organs in reasonable positions! Not so easy to put/keep together, but it’s fine. I would not pay a ton for it, though.
Poster of Velociraptor’s anatomy I’ve had since grad school, adorning my office. For ~1996 (no feathers; “zombie hands“), it’s damn good, mostly… Closer views below (sorry, photo quality is crap– taking photos of wall poster turned out to be harder than I expected! Bad lighting.) :
Closeup of the leg muscles- hey, not bad! Pretty much the right muscles in the right places more or less, and plausible proportions. No air sacs in the torso, but again, this is mid-1990’s science shown. BUT…
I was happy with this poster until I got it home from the western-USA museum I bought it at and read this final bit of text… Oh, America! You silly place. (And unfortunately, these dinosaurs are not from the very end of the Cretaceous anyway, so “global catastrophe” is not implicated.)
Looking down onto the opened torso of the Barcelona ornithomimid. Strikingly realistic! Breastbone, heart, liver, intestines; not unreasonable positions and sizes. No feathers, but again hey– this was made in the earlier days.
Albertosaurus from the Drexel Academy of Sciences. I forget where I got this pic but I like the display– this is an impressive full-scale physical model. The transition from skeleton-only on the left to skinned in the middle to fully-fleshed and popping out atcha on the right is clever.
?T. rex? leg, photo that I took ages ago as a PhD student, if memory serves. Can anyone remind me where this was? California Academy of Sciences? I am embarrassed that I cannot recall. I remember geeking out about it. It has a toy-ish look, but I reckon they had to build this to withstand kids touching it.
Perhaps the best dino-dissection example I’ve seen in a museum: the AMNH’s sauropod Mamenchisaurus with internal organs and their functions projected onto it, in the “World’s Largest Dinosaurs” exhibit. Bravo! I stood and watched it for quite a while.
This is far from comprehensive– just several kinds of imagery that I mostly like. There’s the tension between showing too much realism, which science simply can’t back up, and being too cartoonish, losing the viewer’s immersion in the time-travelling fantasy. I do, however, like other kinds of more abstract, schematic depictions of dinosaur anatomy that simplify the details to focus on the basics of what organs should have been where and how they may have worked, such as this depiction from T. rex Autopsy, which also took the other extreme favouring ultra-realism (but with physical models, not so much with the CGI):
Air flow through a T. rex: simplified but clear. CGI used to explain, not abused. The real air sac anatomy would be too complex to show. You may see something similar with the titanosaur show.
That’s enough for now. I’ve stuck with relatively recent examples; of course in my particular field I also think back to Romer’s wonderful 1920’s drawings, which I covered in this post.
So, blog readers, help me out here: what examples of dinosaur internal, squishy anatomy from museums, documentaries or other not-entirely-done-by-nitpicky-scientists venues do you like, or not like so much? What works for you, or at least is memorable in some way?
I posted the above photo once before, but didn’t explain any of the fun details of artist-designer Thomas Thwaites‘s visit to the RVC to dissect a goat with us. Now his show has just finished in London, celebrating the end of his project and the near-completion of his book about his experience trying to live life as a goat. This week, I went to his east side gallery and had some time to chat with Thomas about his transhuman experiences. Because the project has a strong biomechanics, anatomy, art and science theme to it, I’m posting a photo-blog post about all of that. It’s goat to be seen to be believed! I for one wouldn’t mind being a goat right now; I could use a break from my decrepit body…
Stomach-Churning Rating: Too late, there’s the goat pic above and more like it below. I’d give those a 8/10; no kidding. The puns make it worse, too.
The context. Thomas never did get to gallop (sorry, spoiler!) but he did manage a trot, and some other capricious behaviours. I forgot to ask him if he’d tried the Goat Simulator. I have; it’s good for an hour of fun hircosity.
Starting the dissection at the RVC, to get inside a goat.
Hide.
Fore- and hindlimbs; comparative design for inspiring prosthetics.
Dissections on display!
Prototype goat-suits. Their mobility was too limited.
The prototype in the foreground could not move without falling down.
Inhabited-goat-suit shots.
The final Goat-Suit: custom prosthetics, a helmet, and some form-fitting casts.
Thomas Thwaites with the goat-suit.
The forelimb prosthesis. I was worried it would hurt his wrists but apparently it transferred the loads mainly to the forearms. It was made by a prosthetics clinic up in Salford.
Photos from rambling around the Swiss Alps in the goat-suit with goats.
Trip-trap-trip-trap… (but no trolls)
Goat-suit in action! With Goat-Pro camera, I see.
Acceptance?
And the goat that we had dissected, skeletonized at RVC and re-articulated by Thomas:
Stomach-Churning Rating: 5/10: cheesy songs vs. fatty chunks of tissue; there are no better Crimbo treats!
Today is a special day for palaeognath publications, principally pertaining to the plethora of published PeerJ papers (well, three of them anyway) released today, featuring my team’s research! An early Crimbo comes this year in the form of three related studies of hind limb anatomy, development, evolution and biomechanics in those flightless feathered freaks of evolutionary whimsy, the ratites! And since the papers are all published online in PeerJ (gold open access), they are free for anyone with internet access to download and use with due credit. These papers include some stunning images of morphology and histology, evolutionary diagrams, and a special treat to be revealed below. Here I’ll summarize the papers we have written together (with thanks to Leverhulme Trust funding!):
1) Lamas, L., Main, R.P., Hutchinson, J.R. 2014. Ontogenetic scaling patterns and functional anatomy of the pelvic limb musculature in emus (Dromaius novaehollandiae). PeerJ 2:e716 http://dx.doi.org/10.7717/peerj.716
My final year PhD student and “emu whisperer” Luis Lamas has published his first paper with co-supervisor Russ Main and I. Our paper beautifully illustrates the gross anatomy of the leg muscles of emus, and then uses exhaustive measurements (about 6524 of them, all done manually!) of muscle architecture (masses, lengths, etc.) to show how each of the 34 muscles and their tendons grew across a more than tenfold range of body mass (from 6 weeks to 18 months of age). We learned that these muscles get relatively, not just absolutely, larger as emus grow, and their force-generating ability increases almost as strongly, whereas their tendons tend to grow less quickly. As a result, baby emus have only about 22% of their body mass as leg muscles, vs. about 30% in adults. However, baby emus still are extremely athletic, more so than adults and perhaps even “overbuilt” in some ways.
This pattern of rapidly growing, enlarged leg muscles seems to be a general, ancestral pattern for living bird species, reflecting the precocial (more independent, less nest-bound), cursorial (long-legged, running-adapted) natural history and anatomy, considering other studies of ostriches, rheas, chickens and other species close to the root of the avian family tree. But because emus, like other ratites, invest more of their body mass into leg muscles, they can carry out this precocial growth strategy to a greater extreme than flying birds, trading flight prowess away for enhanced running ability. This paper adds another important dataset to the oft-neglected area of “ontogenetic scaling” of the musculoskeletal system, or how the locomotor apparatus adapts to size-/age-related functional/developmental demands as it grows. Luis did a huge amount of work for this paper, leading arduous dissections and analysis of a complex dataset.
Superficial layer of leg muscles in an emu, in right side view. Click any image here to emu-biggen. The ILPO and IC are like human rectus femoris (“quads”); ILFB like our biceps femoris (“hams”); FL, GM and GL much like our fibularis longus and gastrocnemius (calf) muscles, but much much bigger! Or, perhaps FL stands for fa la la la la?
Data for an extra set of emus studied by coauthor Russ Main in the USA, which grew their muscles similarly to our UK group. The exponents (y-axis) show how much more strongly the muscles grew than isometry (maintaining the same relative size), which is the dotted line at 1. The numbers above each data point are the # of individuals measured. Muscle names are partly above; the rest are in the paper. If you want to know them, we might have been separated at birth!
2) Regnault, S., Pitsillides, A.A., Hutchinson, J.R. 2014. Structure, ontogeny and evolution of the patellar tendon in emus (Dromaius novaehollandiae) and other palaeognath birds. PeerJ 2:e711http://dx.doi.org/10.7717/peerj.711
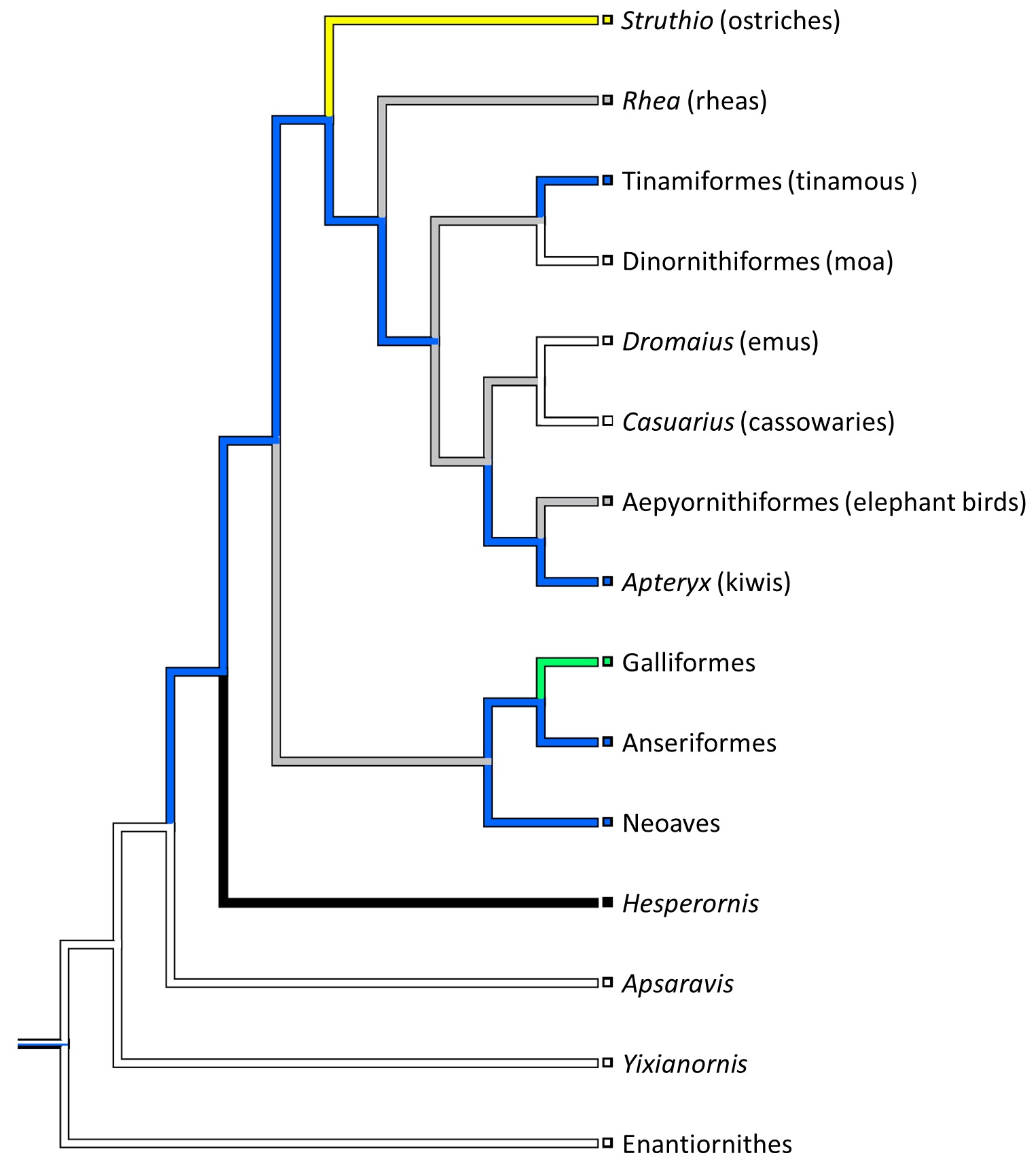
My second year PhD student Sophie Regnault (guest-blogger here before with her rhino feet post) has released her first PhD paper, on the evolution of kneecaps (patellae) in birds, with a focus on the strangeness of the region that should contain the patella in emus. This is a great new collaboration combining her expertise in all aspects of the research with coauthor Prof. Andy Pitsillides‘s on tissue histology and mine on evolution and morphology. This work stems from my own research fellowship on the evolution of the patella in birds, but Sophie has taken it in a bold new direction. First, we realized that emus don’t have a patella– they just keep that region of the knee extensor (~human quadriceps muscle) tendon as a fatty, fibrous tissue throughout growth, showing no signs of forming a bony patella like other birds do. This still blows my mind! Why they do this, we can only speculate meekly about so far. Then, we surveyed other ratites and related birds to see just how unusual the condition in emus was. We discovered, by mapping the form of the patella across an avian family tree, that this fatty tendon seems to be a thing that some ratites (emus, cassowaries and probably the extinct giant moas) do, whereas ostriches go the opposite direction and develop a giant double-boned kneecap in each knee (see below), whereas some other relatives like tinamous and kiwis develop a more “normal”, simple flake-like bit of bone, which is likely the state that the most recent common ancestor of all living birds had.
There’s a lot in this paper for anatomists, biomechanists, palaeontologists, ornithologists, evo-devo folks and more… plenty of food for thought. The paper hearkens back to my 2002 study of the evolution of leg tendons in tetrapods on the lineage that led to birds. In that study I sort of punted on the question of how a patella evolved in birds, because I didn’t quite understand that wonderful little sesamoid bone. And now, 12 years later, we do understand it, at least within the deepest branches of living birds. What happened further up the tree, in later branches, remains a big open subject. It’s clear there were some remarkable changes, such as enormous patellae in diving birds (which the Cretaceous Hesperornis did to an extreme) or losses in other birds (e.g., by some accounts, puffins… I am skeptical)– but curiously, patellae that are not lost in some other birds that you might expect (e.g., the very non-leggy hummingbirds).
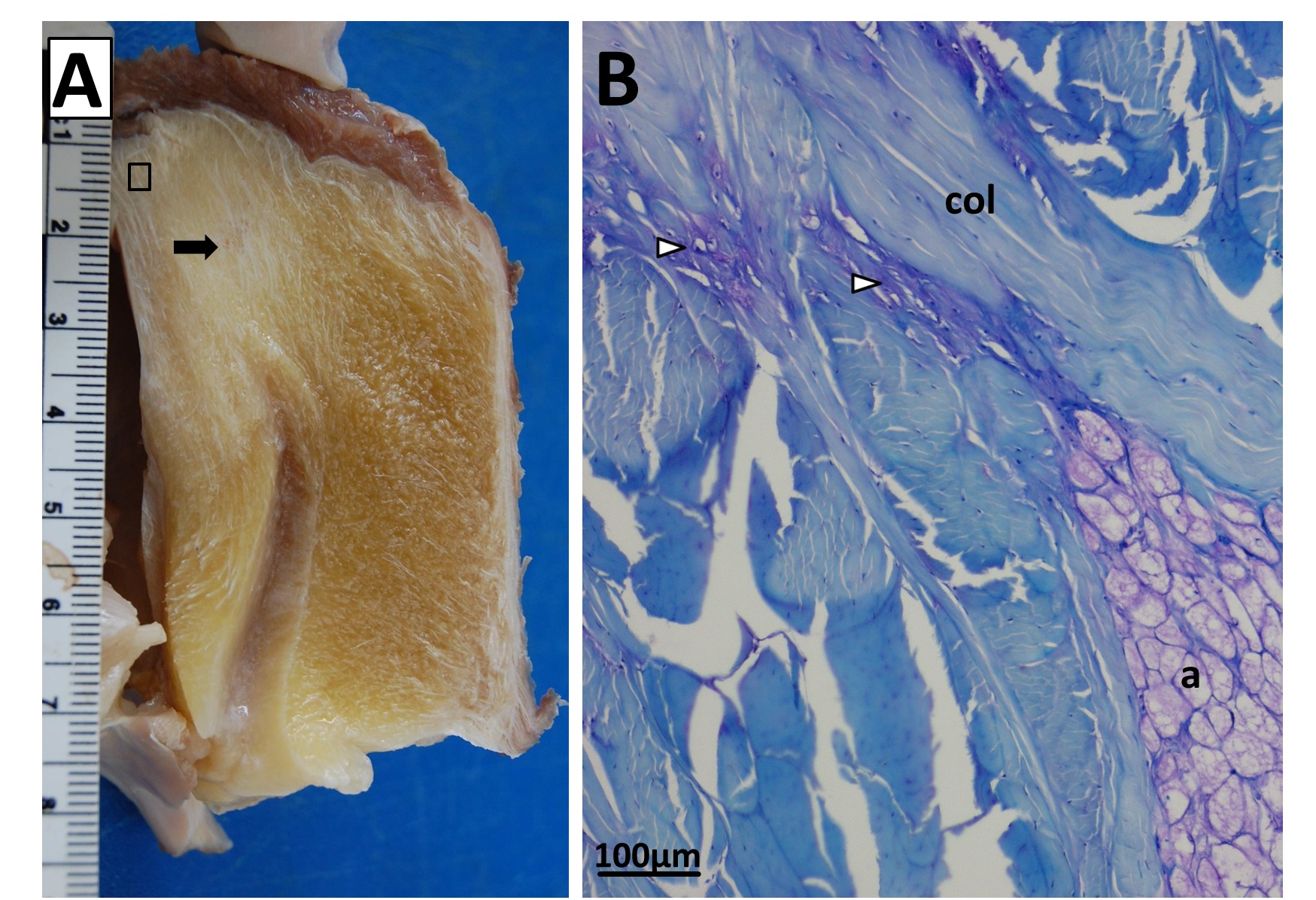
Fatty knee extensor tendon of an emu, showing the absence of a patella. The fatty tissue is split into superficial (Sup) and deep regions, with a pad corresponding to the fat pad in other birds continuous with it and the knee joint meniscus (cushioning pad). The triceps femoris (knee extensor) muscle group inserts right into the fatty tendon, continuing on over it. A is a schematic; B is a dissection.
Sectioning of a Southern Cassowary’s knee extensor tendon, showing: A, Similar section as in the emu image above. revealing similar regions and fibrous tissue (arrow), with no patella, just fat; and B, With collagen fibre bundles (col), fat cells (a), and cartilage-like tissue (open arrows) labelled.
Evolution of patellar form in birds. White branches indicate no patella, blue is a small flake of bone for a patella, green is something bigger, yellow is a double-patella in ostriches, black is a gigantic spar of bone in extinct Hesperornis and relatives, and grey is uncertain. Note the uncertainty and convergent evolution of the patella in ratite birds (Struthio down to Apteryx), which is remarkable but fits well with their likely convergent evolution of flightlessness and running adaptations.
Finally, Kyle Chadwick came from the USA to do a technician post and also part-time Masters degree with me on our sesamoid grant, and proved himself so apt at research that he published a paper just ~3 months into that work! Vivian Allen (now a postdoc on our sesamoid bone grant) joined us in this work, along with Sophie Regnault. We conceived of this paper as fulfilling a need to explain how the major tissues of the knee joint in ostriches, which surround the double-patella noted above, all relate to each other and especially to the patellae. We CT and MRI scanned several ostrich knees and Kyle made a 3D model of a representative subject’s anatomy, which agrees well with the scattered reports of ostrich knee/patellar morphology in the literature but clarifies the complex relationships of all the key organs for the first time.
This ostrich knee model also takes Kyle on an important first step in his Masters research, which is analyzing how this morphology would interact with the potential loads on the patellae. Sesamoid bones like the patella are famously responsive to mechanical loads, so by studying this interaction in ostrich knees, along with other studies of various species with and without patellae, we hope to use to understand why some species evolved patellae (some birds, mammals and lizards; multiple times) and why some never did (most other species, including amphibians, turtles, crocodiles and dinosaurs). And, excitingly for those of you paying attention, this paper includes links to STL format 3D graphics so you can print your own ostrich knees, and a 3D pdf so you can interactively inspect the anatomy yourself!
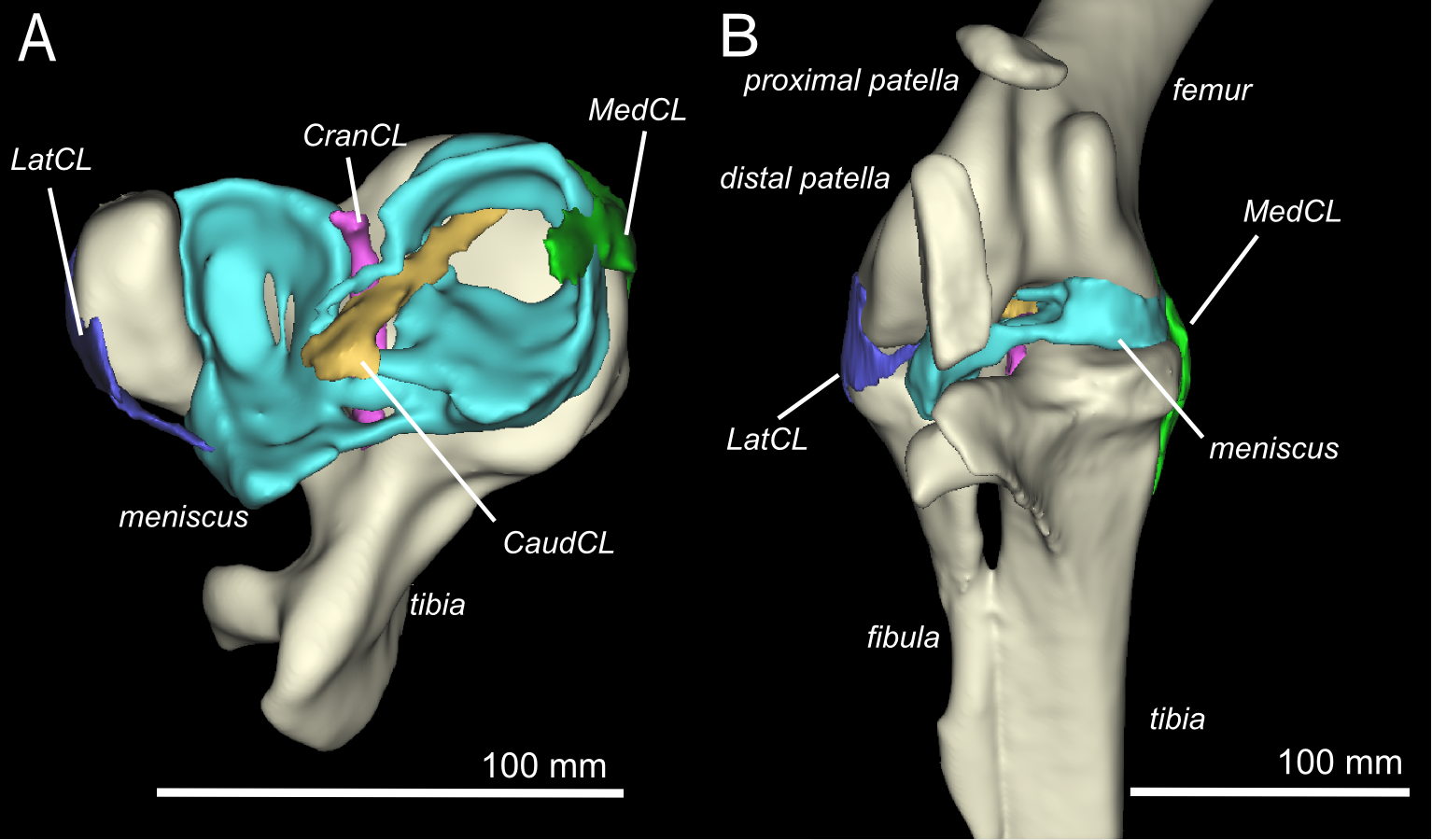
Ostrich knee in side view: A, X-ray, and (B) labelled schematic.
3D model of an ostrich knee, showing: A, View looking down onto the top of the tibia (shank), with the major collateral ligaments (CL), and B, View looking straight at the front of the knee joint, with major organs of interest near the patella, sans muscles.
You can view all the peer review history of the papers if you want, and that prompts me to comment that, as usual at PeerJ (full disclosure: I’m an associate editor but that brings me £0 conflict of interest), the peer review quality was as rigorous at a typical specialist journal, and faster reviewing+editing+production than any other journal I’ve experienced. Publishing there truly is fun!
Merry Christmas and Happy Holidays — and good Ratite-tidings to all!
And stay tuned- the New Year will bring at least three more papers from us on this subject of ratite locomotion and musculoskeletal anatomy!
♬Should auld palaeognathans be forgot, And never brought for scans? Should publications be soon sought, For auld ratite fans!♪
Identify (1) the animal shown in the four-panel top images (CT scan/reconstruction), and (2) the DIFFERENT animal (and/or the main central, pink structure) shown in the big, gooey bottom image (Dissection). No special rules. Potential for double points!
And someone will get these, I am sure. This might be the final round of 2014’s Mystery Anatomy game.