If you go into central Lausanne, Switzerland, you’re likely to pass the Palais du Remine, and if you do, I recommend you go inside. I was happy I did while visiting Lausanne for the AMAM2019 conference. A luxurious palace has been given over to house five (!) free (!) museums on science and culture. These include the canton’s (~state’s) museums of palaeontology and zoology, which I’ll showcase here (also a little of geology and archaeology museums). Tripadvisor’s reviews were good but not as glowing as I’d make mine, so I will remedy that. I’m a sucka for old-school museums, and that’s what these are. So if that sounds right for you, journey onward!

It’s nice.
As you may be expecting by now if you’ve been here before, it’s time for another museum photo blog!
Stomach-Churning Rating: 5/10 for bones, preserved organs, taxidermy aplenty, and animal developmental deformities.

Nice cathedral nearby, w/great view of the city.

Nice interior architecture. There’s lots of nice to behold.

Posters That Get You Excited 101. But you must wait. Like I did.

Quadrupedal human at Zoology museum entry.

Tomistoma, false gharial.

Not a bad collection of taxidermied Crocodylia!

Visually arresting cobra display.

I’ve never seen three Draco gliding lizards on display together!

Bipedal lizard taxidermy displays, freezing the dynamic in the static, are no easy feat.

Plenty of stuffed animals like these raptors/other large birds. Classical zoology museum style. Minimal signage. Just specimen labels, mainly.

Coelacanth!

Sperm whale jaw.

Open space with big specimens. A ~4m long great white shark included.

Second zoology hall: bones!

Gorilla standing tall next to human.

Ostrich skeleton up close, amongst the mammalz.

Cassowary skeleton.

Emu shoulder/arm bones in right side view.

Walrus skeleton in what seems like an odd pose to me, but then they are odd on land.

Alligator skeleton in repose.

Giant anteater, “knuckle-walking”.

Pangolin skeleton! And mounted digging into a nest– very well done!

Bernard Heuvelmans display, about the (in)famous cryptozoologist. This was quite a surprise to me. I’m sure I’d read his English-translated book “On the Track of Unknown Animals” as a kid, during my long stint as an avid reader of much zoology, crypto- and otherwise. He bequeathed a lot of his work to the museum.

Bernard’s handwritten CV!? With a “sea serpent” sketch.

A “sea serpent” vertebra… but if you know any anatomy, it’s not a snake’s vertebra at all but a fish’s, such as a basking shark‘s.

Are you ready for more weirdness? How about some “mutants”- congenital deformities of animals? Fascinating errors of developmental anatomy… somehow this two-headed calf survived awhile. Plenty more where that came from, as follows:





And then there’s all kinds of wonderful comparative anatomy. To be a student of this subject in Lausanne would be a lucky thing, with this museum’s collection at hand. These are valuable specimens, made with love and skill.

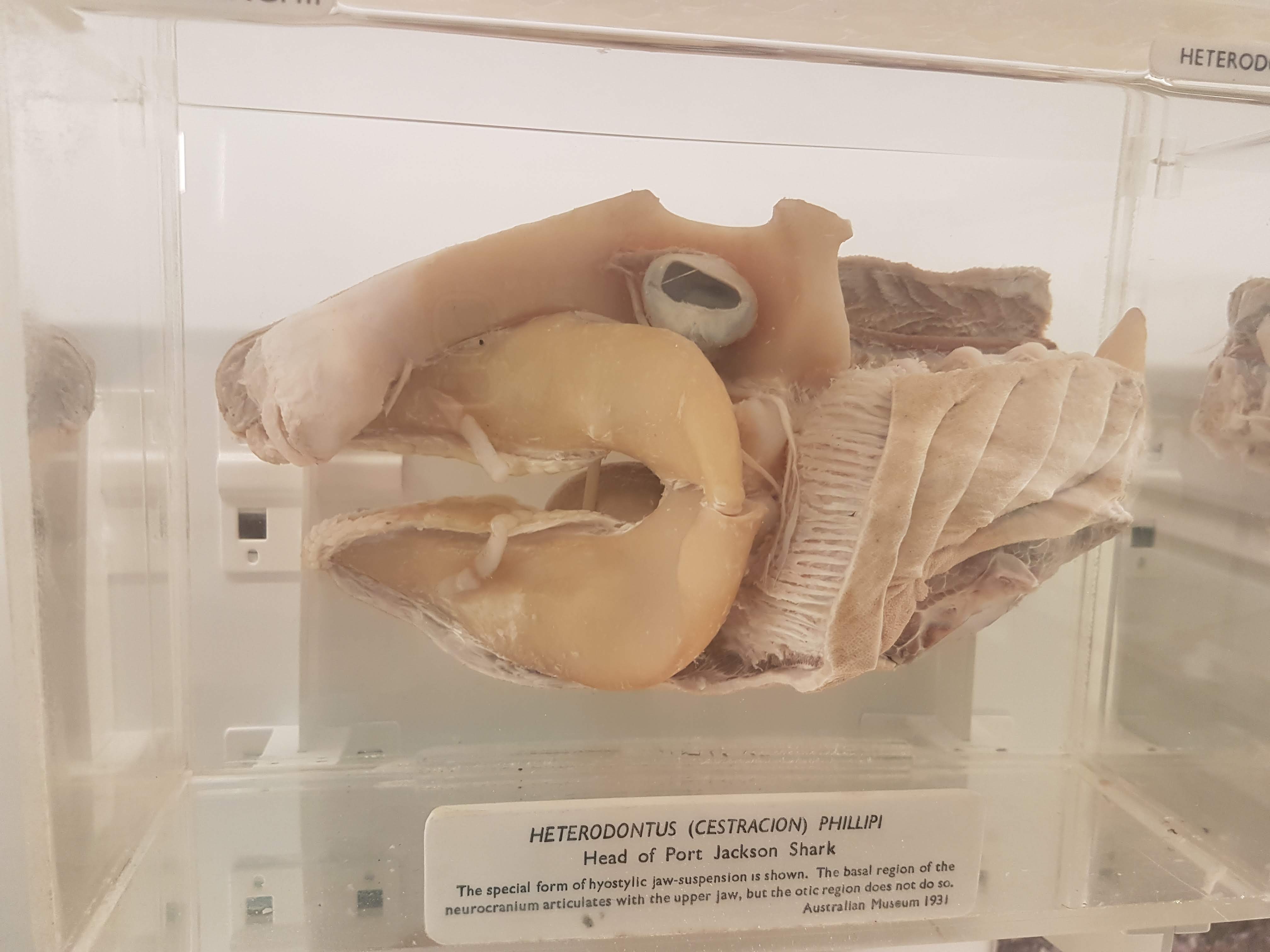
Jaws

Fish head anatomy. Some vertebrae on the left, too.

Developmental regions of the head: a lovely wax(?) model of an Echidna skull. A treasure.

Brains: alligator vs. pigeon.

Salamander muscles.

Pigeon muscles.

More spotted felids than you can shake a jar of catnip at.

Another pangolin!

Giant armadillo.

Petaurus: flying phalanger (a gliding marsupial).

Second zoology hall open area: left side.

Second zoology hall open area: right side.

A final hall with a more new-fangled display, on the topic of evolution and extinction. Attractive phylogeny graphic here. Birds at the “top”, of course. Poor lowly mammals!

Taxidermied giant auk- not a common sight! (Extinct)

The extinct southern pig-footed bandicoot. Also a rare sight of a whole specimen- in a Swiss museum, too.
NOW ON TO THE FOSSILS!
You’ve been very patient. Here, have a Toblerone.

Palaeo museum entry. Already there are cool things visible. Inside, we find it just like I prefer my zoo/palaeo museums (as above): stuffed with specimens and leaving plenty for you to wonder about and investigate. Not frilly; a well-stocked museum that mostly lets its specimens speak for themselves.

Sauriermuseum (Aathal) specimen of Plateosaurus: sculpt/cast. A very good, big skeleton of this common dinosaur, rearing up.

Rear view of same.

Real bones of same; vertebrae and pelvic (this is the “Frick specimen”).

Metaxytherium (current name), an ancient and large fossil dugong/seacow. Skull is in left side view. (that may help, as their skulls are odd!)

Anthracotherium upper jaw: ancient hippo-cousin.

Prolagus: the “Sardinian hare” (recently extinct; old lineage).

Potamotherium: to some an early otter-like mammal, more recently thought to be an ancient seal.

“Broke-ulum”: a walrus broke its penis bone (baculum) and was surely not pleased about it, but lived to heal— physically if not mentally. Yeesh!

Glyptodont tail club and armour.

Aepyornis elephant bird legs!

A partial/reconstructed skeleton of the dodo.

Velociraptor preparing to pounce from above. It’s too late for you!

Rhamphorhynchus fossil (2D slab) and sculpt/cast coming alive in 3D– good stuff.

Anhanguera pterosaur watches the chaos from above, fish snagged in its teeth.

Not-shabby metriorhynchid marine croc fossils, from Britain.

Lovely 3D plesiosaur bones (flippers, neck, etc.) from near RVC: Peterborough!

Mesosaur; early reptile.

The museum clearly is proud of its excellent “Mammoth of Brassus” skeleton, essentially complete.

Ice Age elk/moose, a 10,000 year old skeleton in fine shape.

Cave bear skull rawr

Purty ammonites!

Spiky ammonite!

Cretaceous sponge colony from France. I hadn’t seen something like this before, so here it is.

Trilobites, brittlestars and friends.
Well I did wander through the geology and archaeology museums too, and while I liked them I did not take so many photos. My non-human organismal bias is apparent. But check these final ones out:

Splendid cross-section of the stratigraphy of the Alps around Lausanne. I gazed at this for quite a few minutes, trying to figure out what was where in the landscape I’d seen and how old, how deformed, etc.

Slab of “dinosaur” tracks but it was not clear to me what dinosaurs/archosaurs/whatever made them. I wish my French was better. Closeup below shows two footprints superimposed.


At last, the coup de grace! What museum would be complete without a diorama!? (I love them) This one, with a goat sacrifice and early Stone Age people praying to heathen deities/spirits at an elaborate petroglyph array rocked my world. And so it makes a perfect final image. Enjoy, and conduct the proper rites. \m/