Our special guest post this week comes from Dr. Liz Clark of Yale University in New Haven, Connecticut, USA. She continues to bring biomechanics-fu to echinoderms– the weird marine critters like seastars and sea urchins. Including fossils, as you’ll see today! You may remember her from blog posts such as “Guest Post: Brittle Star Arms Are Weird“.
Stomach-Churning Rating: 1/10; echinoderms are inoffensive.
Imagine that you’re stuck in a cardboard box on the beach, holding a small stick. Could you use the stick to move yourself forward? What would you do? You could try digging into the sediment ahead of you to pull yourself along. You could try rowing side to side, as if you were in a rowboat. Or maybe it’s not possible and you’d give up, decide to stay put, and wave your stick in the air for help.
Believe it or not, this is a strange-but-important dilemma that some paleobiologists- like me!- have been wrestling with for generations. My research specialty is in the biomechanics of locomotion– how organisms use their bodies to get from one place to the next (through walking or swimming, for instance). We can learn a lot about an animal by studying their locomotion, such as why their body is shaped the way that it is, or what role they occupy in their ecosystem. Animal motion is a major inspiration for robotic design, and I work with engineers to apply the novel insights on animal locomotion from my research to create new kinds of devices.
Studying the biomechanics of motion in living organisms is (relatively) straightforward. We can use high-speed cameras, motion capture software, and 3D imaging tools to visualize and understand how organisms move in real-time, informing our inferences about how they perform certain tasks. Inferring locomotion in fossil organisms, on the other hand, is tricky since we can’t observe the organism’s behavior like we could if the organism were alive. Instead of being able to watch the organism move, we’re left with a snapshot of the animal frozen in place in a rock. We’re also missing a lot of physical information: locomotion in most animals requires soft tissue and hard skeletal structures, but typically with fossils, only the hard structures get preserved.
However, we can often garner some insights from living organisms to determine the locomotion strategies that fossil organisms use. Most organisms in the fossil record look at least somewhat similar to organisms alive today. If our fossil has four legs, for instance, we can study locomotion in living tetrapods (four-legged animals) to help us create a framework for deriving inferences about locomotion in our extinct tetrapod fossil animal. But for some really strange-looking animals- ones without obvious modern analogues- we’re not so lucky. For me, this is where the fun begins.
Figure 1: Stylophorans! Here are four fossilized stylophorans from the Helderberg Group of the Early Devonian (YPM 036413)
So getting back to the cardboard box and the stick. These are metaphorical examples of the different locomotion strategies that have been proposed for a group of fossil animals known as stylophorans (Figure 1). Stylophorans are extinct organisms related to sea stars and sea urchins, but with a body structure unlike any organism on the planet today. They have a large, relatively flat body called a theca (i.e., the cardboard box), and a long, thin segmented tail known as the aulacophore (i.e. the stick) (Figure 2). They’re known in the paleontological community as some of “the strangest-looking animals of all time.”
Figure 2: Stylophoran anatomy. The “theca” is the body cavity, and the “aulacophore” comprises of the proximal aulacophore, the stylocone, and the distal aulacophore.
By reconstructing stylophoran locomotion, we can unlock the mechanics of a unique system for motion and its potential applications to engineering. We can also understand more about how this organism lived and functioned in its ancient ecosystem. And, by developing a new approach to understand locomotion in stylophorans, we can apply this strategy to analyze locomotion and movement in other unusual fossil animals as well!
For years, scientists have been documenting the incredible array of stylophoran diversity in the fossil record and making their best predictions about how they would have been able to move (or not!). These predictions are based on their morphology– the structure of an organism’s body. For stylophorans, that means the shape and structure of the theca and aulacophore. There are a variety of stylophoran thecal shapes, ranging from ovoid in Enopleura to trapezoidal in Ceratocystis to almost crescent-shaped in Cortnurnocystis. There’s a similarly wide array of aulacophore morphologies as well.
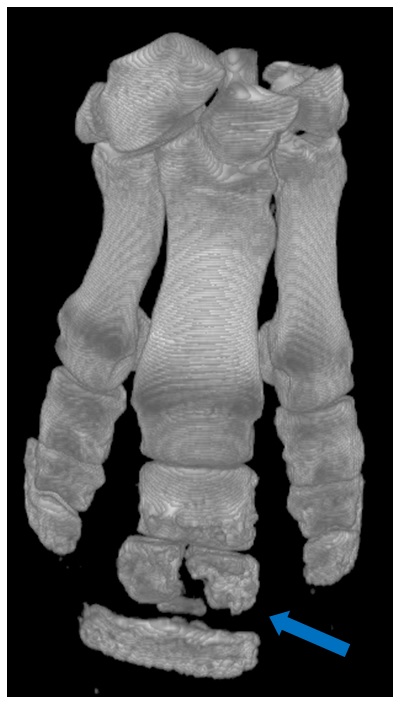
Figure 3: Left: One half of the concretion within which the stylophoran fossil we analyzed is preserved. Right: The 3D digital image of the stylophoran fossil, created by micro-CT scanning the fossil specimen.
We developed a new approach using 3D imaging (Figure 3) to create a digital model of a stylophoran specimen. We used the model to test if several different locomotion strategies that had been proposed before were physically possible or impossible for a stylophoran to actually perform.
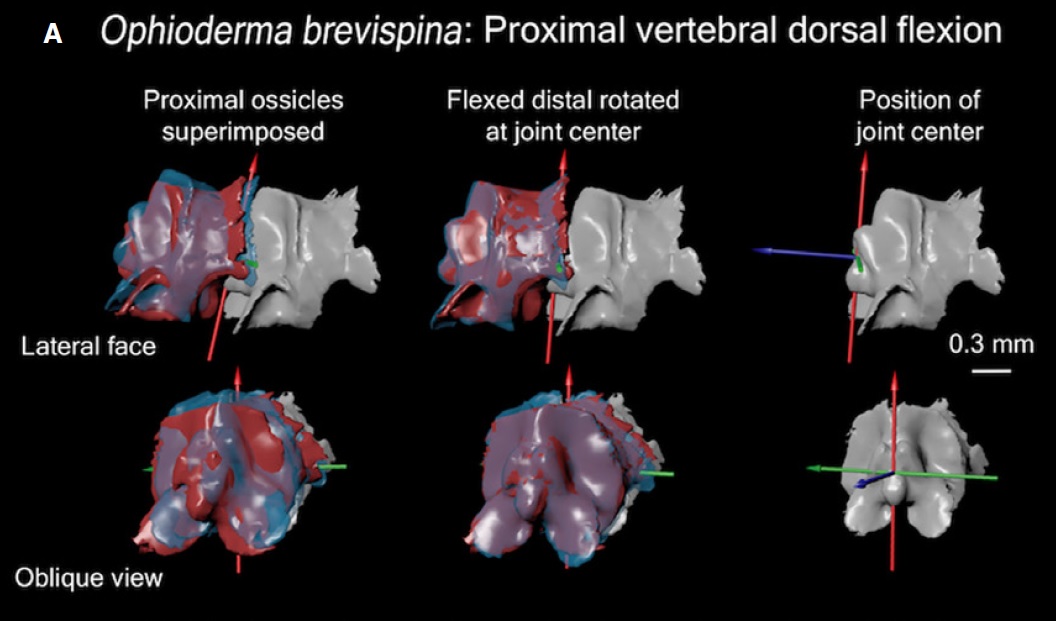
First, we used a micro-CT scanner to image a fossil stylophoran. This outputs a digital 3D picture of the stylophoran fossil that we can look at and analyze on a computer. Next, we developed a program to calculate the joint centers- the point at which one skeletal structure rotates relative to another-within the digitized stylophoran’s aulacophore (Figure 4). This created a digital marionette– a rig of our stylophoran fossil that flexes at the junctures between aulacophore segments as it would have in life. We then rotated each segment at the joint center to calculate the aulacophore’s total range of motion– a reconstruction of how far the aulacophore could flex in each direction (Figure 5).
Figure 4: A look into some of the nuts and bolts of the 3D model we created. Tri-colored axes demarcate where the joint centers are in the proximal aulacophore.
We used this 3D range of motion model to evaluate several locomotion strategies that had been previously hypothesized for this group of stylophorans. One hypothesis suggested that these stylophorans dug their aulacophores into the substrate– sediment on the ocean floor- to pull themselves forward. Another suggested that they moved the aulacophore side to side in order to push themselves along. We found that the first hypothesis would have been impossible to conduct based on the range of motion we calculated, but the second strategy was theoretically possible! We’ll need to do more work to see how likely it was that stylophorans would have actually used this technique. Nevertheless, through this investigation, our team produced the first objective, data-driven methodology for analyzing locomotion in fossil invertebrates, which is a big step in the right direction for the study of fossil invertebrate biomechanics! Our technique can be applied to study other organisms with rigid skeletons as well, like crabs, insects, or sea stars, for instance, and we’re looking forward to seeing our technique used to uncover more interesting locomotion strategies!
Figure 5: A snapshot of the 3D model where we can observe how dorsal and ventral range of motion compare to the originally preserved orientation of the aulacophore (highlighted in green).
Eversible Stomach-Churning Rating: 2/10; no Uni sushi here.
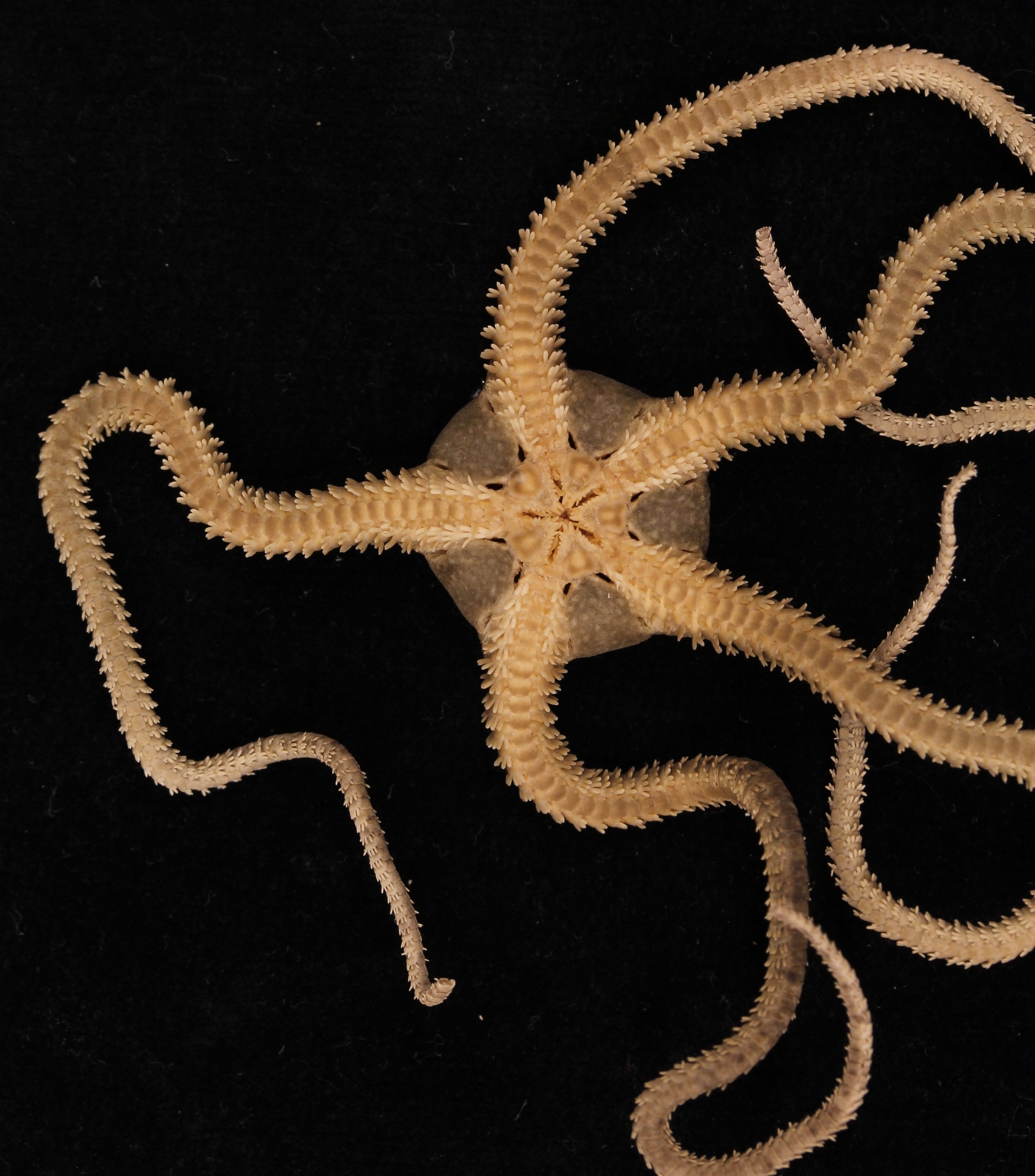
I remember the first time I saw one. I was at the Duke Marine Lab staring at a chunk of dredged-up oyster shells in a glass dish, when all of a sudden a mass of big, black spines obscured my view. I looked up from the microscope to see a creature with a round body the size of a nickel and a flurry of long, skinny, spiny arms skulking hurriedly across the dish. It wasn’t quite a spider- the five-fold symmetry gave its echinoderm affinity away- but it wasn’t quite a starfish, either. Starfish appear graceful as their tiny tube-feet make hurried and unseen movements underneath them to transport them slowly across the sand- appearing nearly motionless to the naked eye. This animal, on the other hand, was making rapid, whip-like strikes with its arms so that it clambered forward, rapidly and fearlessly scaling the uneven terrain of the shells in a bold attempt to escape the dish. I was hooked. I had to know who this monster was, and learn as much about it as I could.
Brittle star arm set up to study its ossicle-joint mobility with CT scanning (below).
That was the day I was introduced to the brittle star. The name “brittle star” is a bit of a misnomer, since they are really anything but. Brittleness implies rigidity and stiffness, suggesting they have a delicate nature with the impossibility of repair or to adapt, which couldn’t be farther from the truth. Their long arms are incredibly flexible, each made of around 100 tiny segments that allow them to bend in any direction or loop them around in circles. I bet that their name comes from the ease at which they can cast off their arms, which they do intentionally to escape predators or pesky researchers trying to grab them, which deceitfully suggests fragility when in fact their arms are incredibly sturdy and packed with powerful muscles. They can flawlessly regenerate their arms, and, in the meantime, even after they lose several of them, they adjust their strategy for locomotion so that they keep prowling across the seafloor unphased. Their physical flexibility and ability to repair and adapt in the face of damage makes them anything but brittle. The Japanese name for brittle star roughly translates to “spider-human-hand,” which I think much more accurately captures the ethos of this group.
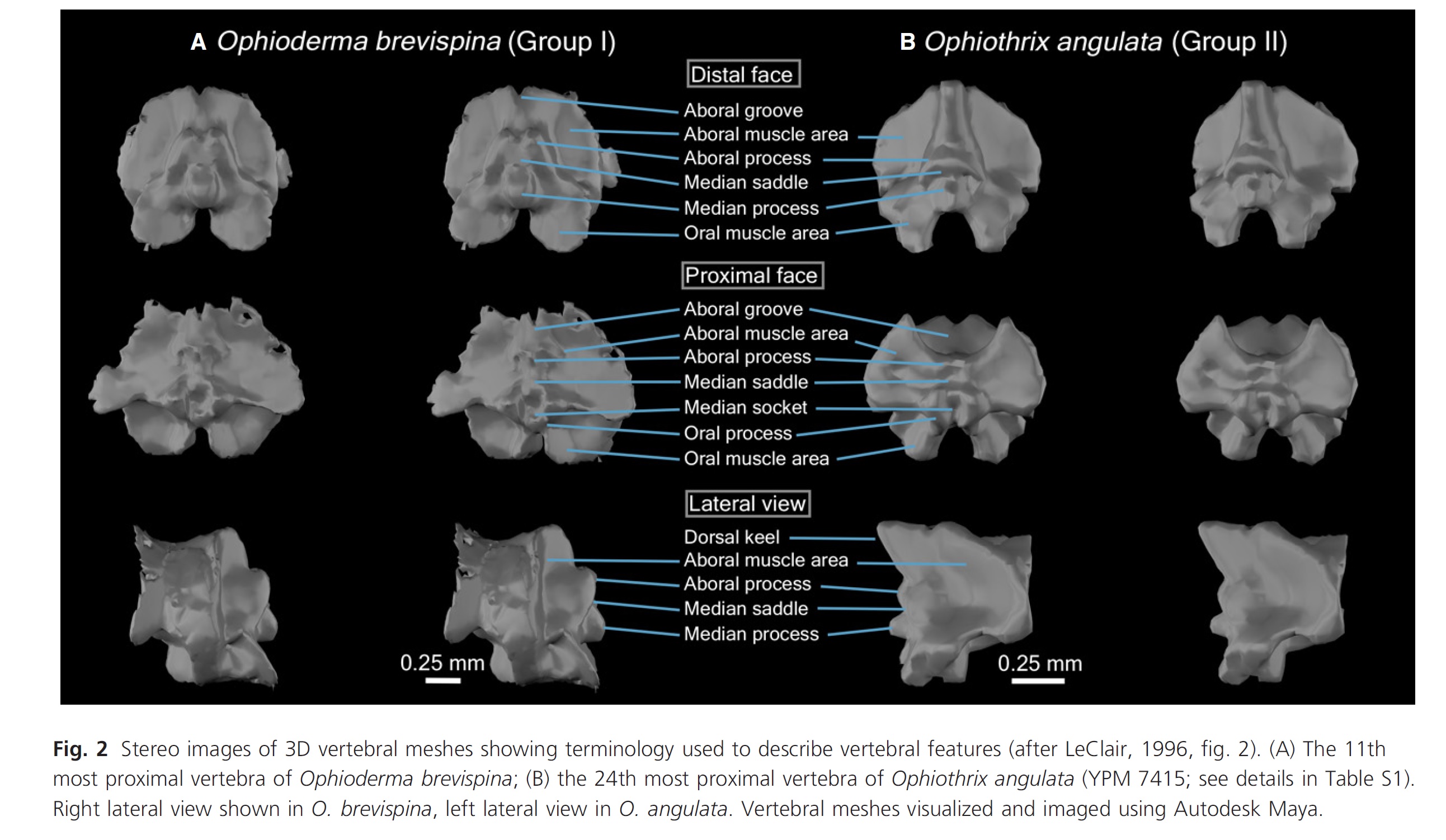
Brittle stars have internal skeletons, and each segment of their arms are made of a cluster of small skeletal elements (ossicles). Researchers in the past have made the assumption that differences in the shape of these ossicles between species change how they move, but I wasn’t so sure. So, John and I decided to work together to figure it out.
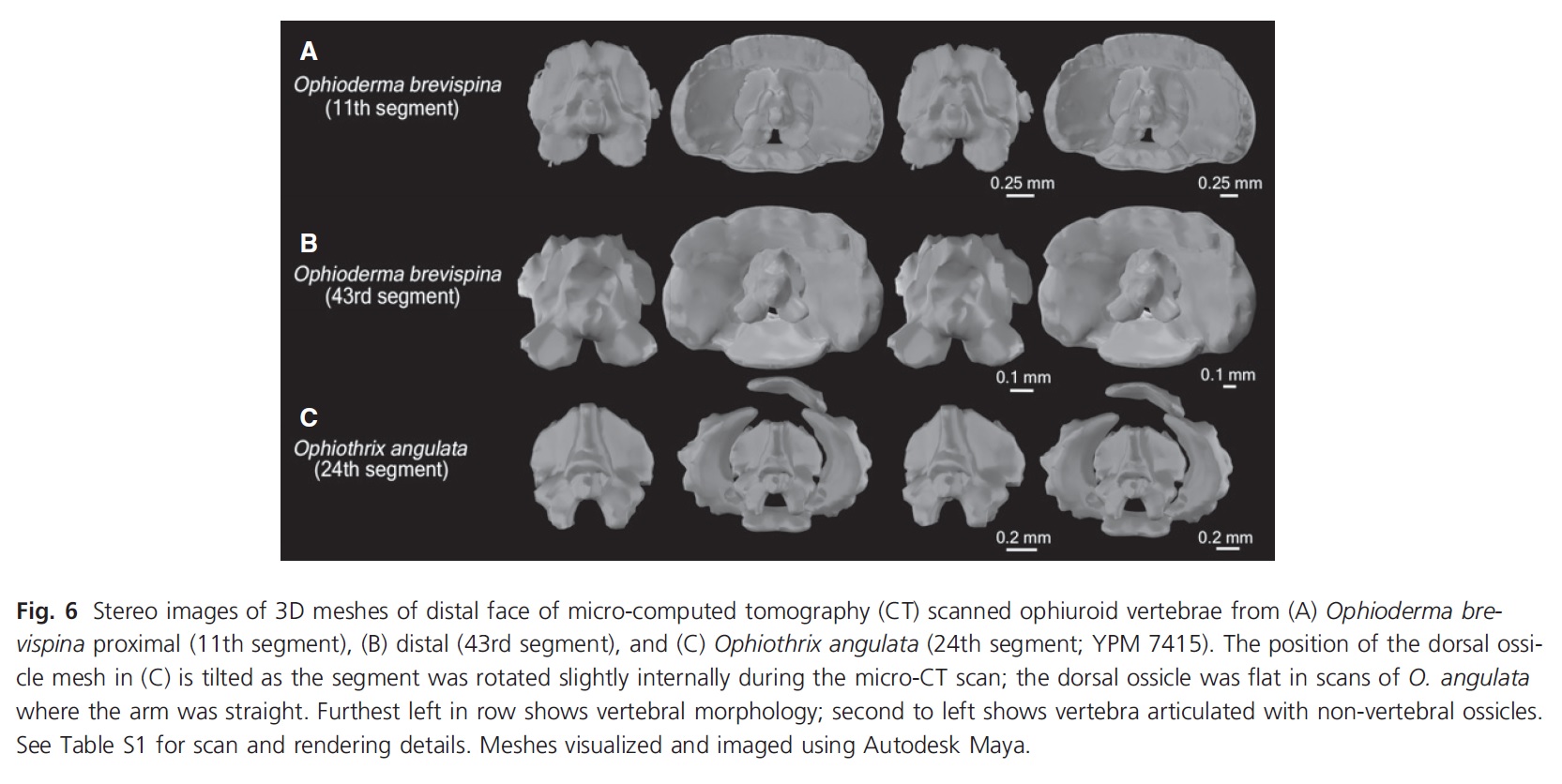
We didn’t dive into the freezer for this one- sorry to disappoint all of the diehard fans of John’s freezer out there (but in my defense can you imagine how tough it would have been to even find them in the sea of rhinos, giraffes, and crocs?!). [JOHN: awwwwwww!! It’s more of a wall keeping in the wildlings, than a sea right now though!] Instead we ordered some brittle stars off the internet! The first thing we did was make some measurements of how flexible the arms of brittle stars are when they’re alive. Then we digitized their skeletons by micro-CT scanning them so we could see the articulations between the ossicles and the segments in 3D. We scanned them in a few different positions so we could see the articulations between the ossicles as their arms bend. Then we incorporated all of that data into a 3D model that allowed us to visualize what’s going on in the inside of brittle star arms as they move them around.
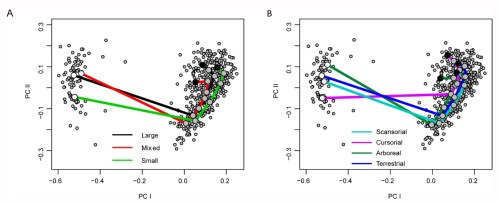
We made several different models using this strategy to see if different ossicle shapes change how their arms move. We looked at the differences between arm ossicles in two different species– Ophioderma brevispina and Ophiothrix angulata, which represent two of the three different major morphologies of brittle star arms. We also looked at the difference in the movement mechanics at the tip and base of the arms in O. brevispina, since the ossicles at the tip are thin and elongated compared to wide and flat at the base.
We found that the tip of the arm of Ophioderma brevispina was more flexible than the base due, at least in part, to the shape of the ossicles. We also found several major differences between the two species, including the location of their joint center and the degree to which they could laterally flex. However, none of these differences were easily attributable to any specific morphological feature that set Ophiothrix angulata and O. brevispina apart, which cautions against making assumptions of brittle star functional capabilities by only looking at the shape of the ossicles. We also found that some of the smaller ossicles within each segment shift their position to accommodate arm flexion, when they were originally thought to limit the motion of the arm! We only looked at a few individuals of two species, but the methods for model-building we developed provide a framework to incorporate a broad sample of brittle star species in the future. We’re curious if the results we found stand when more brittle stars are brought into the mix!
It was incredible to take the journey from initially being surprised and captivated by the movement of these animals to eventually building 3D digital models to discover how they are able to do so. It made me realize that opportunities to be inspired by the natural world are around every corner, and that there are so many interesting questions out there that are still unanswered. Thanks to John and our other team members Derek Briggs, Simon Darroch, Nicolás Mongiardino Koch, Travis Brady, and Sloane Smith for making this project happen!
Stomach-Churning Rating: 1/10; just bones and data plots!
It is often said that all cats are very similar in terms of their skeletal morphology (“a cat is a cat is a cat”). But is this really the case? It may be if only gross, qualitative anatomy is taken into consideration, i.e., if you just eyeball the skeletons of tigers and lions you might find yourself not knowing which one is which. But with huge advances in technology that allows for extracting detailed shape information off a structure (e.g., a skull) and for analysing this information (‘Geometric Morphometrics’), it has become more and more possible to distinguish between relatively similar forms – which may be from distinct species, separate sexes, or even just different populations of the same taxon.
And it is reasonable to think that cat skeletons might be a lot more different than what meets the eye, as for a lineage of apparently similarly built animals, with not that much variation in diet (all cats are hypercarnivores) there is substantial variation in body mass (over 300-fold just in living species!) and in ecology across cat species. From the cursorial cheetah to the arboreal clouded leopard, felids present a wide range of locomotory adaptations. Yes, all cats can climb, but some do it better than others: think lion versus margay (yes, they do descend trees head-first). As hypercarnivores, all cats are meat specialists, but they also change with regards to how big their prey is, with a general and sometimes-not-so-black-and-white three-tier classification into small, mixed and large prey specialists. The rule of thumb is ‘if you are lighter than ~20-25 kg, hunt small stuff. If you are heavier than that, hunt BIG BIG things; bigger than yourself. And if you are in the middle ground, hunt some small-ish things, some big-ish things, and things about your size. Well, -ish’ – their prey size preference has a lot to do with energetic constraints (have a look at Carbone et al. 1999; and Carbone et al. 2007, if you’re interested in this). But the fun bit here is that form sometimes correlates quite strongly with function, so we should be able to find differences in some of their bones that carry this ecological signal.
Indeed, for a while now, we have known that the shape of the skull and limbs of felids can tell us a lot about how they move and how big their prey is (Meachen-Samuels and Van Valkenburgh 2009, 2009), but a large proportion of their skeleton has been largely ignored: we don’t know half as much about ecomorphology and evolution of the vertebral column. Well, it was time we changed this a bit! As the PhD student in the Leverhulme-funded ‘Walking the cat back’ (or more informally, “Team Cat”) project, I’ve spend a big chunk of my first two years travelling around the world (well, ok, mainly to several locations in the USA) carrying a heavy pellet case containing my working tool, a Microscribe, to collect 3-D landmarks (Fig. 1) across the presacral vertebral column of several cat species. And some of first results are just out! Check them out by reading our latest paper, “Regional differentiation of felid vertebral column evolution: a study of 3D shape trajectories” in the Organisms Diversity and Evolution journal (Randau, Cuff, et al. 2016).
Fig. 1:Different vertebral morphologies and their respective three-dimensional landmarks. Vertebral images are from CT scans of Acinonyx jubatus (Cheetah, USNM 520539)
Building from results based on our linear vertebral data from the beginning of the year (Randau, Goswami, et al. 2016), the 3-D vertebral coordinates carry a lot more information and we were able to describe how this complex shape-function relationship takes place throughout the axial skeleton (in cats at least) in much better detail than our prior study did. One of the difficulties in studying serial structures such as the vertebral column is that some clades present variation in vertebral count which makes it less straightforward to compare individual vertebrae or regions across species. However, mammals are relatively strongly constrained in vertebral count, and Felidae (cats; living and known fossils) show no variation at all, having 27 presacral vertebrae. So adaptation of the axial skeleton in mammals has been suggested to happen by modification of shape rather than changes in vertebral number.
Using a variety of geometric morphometric analyses, under a phylogenetically informative methodology, we have shown that there is clear shape and functional regionalisation across the vertebral column, with vertebrae forming clusters that share similar signal. Most interestingly, the big picture of these results is a neck region which is either very conservative in shape, or is under much stronger constraints preventing it from responding to direct evolutionary pressures, contrasting with the ‘posteriormost’ post-diaphragmatic tenth thoracic (T10) to last lumbar (L7) vertebral region, which show the strongest ecological correlations.
We were able to analyse shape change through functional vertebral regions, rather than individual vertebrae alone, by making a novel application of a technique called the ‘Phenotypic Trajectory Analysis’, and demonstrated that the direction of vertebral shape trajectories in the morphospace changes considerably between both prey size and locomotory ecomorphs in cats, but that the amount of change in each group was the same. It was again in this T10-L7 region that ecological groups differed the most in vertebral shape trajectories (Fig. 2).
Figure 2:Phenotypic trajectory analysis (PTA) of vertebrae in the T10 – L7 region grouped by prey size (A) and locomotory (B) categories.
So in the postcranial morphology of cats can be distinguished, changing its anatomy in order to accommodate the different lifestyles we see across species. But the distinct parts of this structure respond to selection differently. The next step is figuring out how that might happen and we are working on it.
Carbone, C., Mace, G. M., Roberts, S. C., and Macdonald, D. W. 1999. Energetic constaints on the diet of terrestrial carnivores. Nature 402:286-288.
Carbone, C., Teacher, A., and Rowcliffe, J. M. 2007. The costs of carnivory. PLoS biology 5 (2):e22.
Meachen-Samuels, J. and Van Valkenburgh, B. 2009. Craniodental indicators of prey size preference in the Felidae. Biol J Linn Soc 96 (4):784-799.
———. 2009. Forelimb indicators of prey-size preference in the Felidae. Journal of morphology 270 (6):729-744.
Randau, M., Cuff, A. R., Hutchinson, J. R., Pierce, S. E., and Goswami, A. 2016. Regional differentiation of felid vertebral column evolution: a study of 3D shape trajectories. Organisms Diversity and Evolution Online First.
Randau, M., Goswami, A., Hutchinson, J. R., Cuff, A. R., and Pierce, S. E. 2016. Cryptic complexity in felid vertebral evolution: shape differentiation and allometry of the axial skeleton. Zoological Journal of the Linnean Society 178 (1):183-202.
Greetings Freezerinos, and Happy New Year! I have been quiet on this blog for health and other reasons but those will pass and there will be new posts in 2016. However, behind the scenes there have been super-cool things afoot. I am very happy to bring one of them to you now:
(but first: Stomach-Churning Rating: 6/10; video below shows a dissected sea turtle foot in motion)
We have just debuted our new social media “presence” (for lack of a better word) that is a sister blog to this one. It is called Anatomy To You (http://anatomytoyou.com/), as its intent is to bring a wide array of science about animal anatomy to “you”, the general public. This John’s Freezer blog will continue with it’s style of rambling longer posts targeted at a fairly geeky scientifically literate audience and focusing on my team’s research and my own disparate thoughts about science and related issues. Anatomy To You will bring you shorter posts, even just images, completely focused on celebrating the structure of organisms, and not just presenting my team’s research but also a wide array of anatomical science from around the globe. It will also be much more regular and frequent in its posts. We’ll welcome guest posts and I encourage you to get in touch with us if you want to jump on the bandwagon early, or have us feature your research for you!
Anatomy To You will continue to evolve over this coming year, so please stay with us and give us feedback; join in the morphological conversations with us. I am SUPER excited to see where this goes– it is an experiment that has a lot of potential, we think.
Sea turtle from our ATY dissection, foot muscles in action (found dead in the wild; don’t be ridiculous, we don’t kill sea turtles for our research)
Seeking adaptations for running and swimming in the vertebral columns of ancient crocs
A guest post by Dr. Julia Molnar, Howard University, USA (this comes from Julia’s PhD research at RVC with John & colleagues)
Recently, John and I with colleagues Stephanie Pierce, Bhart-Anjan Bhullar, and Alan Turner described morphological and functional changes in the vertebral column with increasing aquatic adaptation in crocodylomorphs (Royal Society Open Science, doi 10.1098/rsos.150439). Our results shed light upon key aspects of the evolutionary history of these under-appreciated archosaurs.
Stomach-Churning Rating: 5/10; a juicy croc torso in one small photo but that’s all.
Phylogenetic relationships of the three crocodylomorph groups in the study and our functional hypotheses about their vertebrae. * Image credits: Hesperosuchus by Smokeybjb, Suchodus by Dmitry Bogdanov (vectorized by T. Michael Keesey) http://creativecommons.org/licenses/by-sa/3.0
As fascinating as modern crocodiles might be, in many ways they are overshadowed by their extinct, Mesozoic cousins and ancestors. The Triassic, Jurassic, and early Cretaceous periods saw the small, fast, hyper-carnivorous “sphenosuchians,” the giant, flippered marine thalattosuchians, and various oddballs like the duck-billed Anatosuchus and the aptly named Armadillosuchus. As palaeontologists/biomechanists, we looked at this wide variety of ecological specializations in those species, the Crocodylomorpha, and wanted to know, how did they do it?
Of course, we weren’t the first scientists to wonder about the locomotion of crocodylomorphs, but we did have some new tools in our toolbox; specifically, a couple of micro-CT scanners and some sophisticated imaging software. We took CT and micro-CT scans of five fossil crocodylomorphs: two presumably terrestrial early crocodylomorphs (Terrestrisuchus and Protosuchus), three aquatic thalattosuchians (Pelagosaurus, Steneosaurus, and Metriorhynchus) and a semi-aquatic modern crocodile (Crocodylus niloticus). Since we’re still stuck on vertebrae (see, e.g., here; and also here), we digitally separated out the vertebrae to make 3D models of individual joints and took measurements from each vertebra. Finally, we manipulated the virtual joint models to find out how far they could move before the bones bumped into each other or the joints came apart (osteological range of motion, or RoM).
Our methods: get fossil (NHMUK), scan fossil, make virtual fossil and play with it.
Above: Video of a single virtual inter-vertebral joint from the trunk of Pelagosaurus typus (NHMUK) showing maximum osteological range of motion in the lateral direction (video). Note the very un-modern-croc-like flat surfaces of the vertebral bodies! (modern crocs have a ball-and-socket spinal joint with the socket on the front end)
While this was a lot of fun, what we really wanted to find out was whether, as crocodylomorphs became specialized for different types of locomotion, the shapes of their vertebrae changed similarly to those of mammalian lineages. For example, many terrestrial mammals have a lumbar region that is very flexible dorsoventrally to allow up-and-down movements during bounding and galloping. Did fast-running crocodylomorphs have similar dorsoventral flexibility? And did fast-swimming aquatic crocodylomorphs evolve a stiffer vertebral column like that of whales and dolphins?
Above: Video of how we modelled and took measurements from the early crocodylomorph Terrestrisuchus gracilis (NHMUK).
Our first results were puzzling. The Nile croc had greater RoM in side-to-side motions, which makes sense because crocodiles mostly use more sprawling postures and are semi-aquatic, using quite a bit of side-to-side motions in life. The part that didn’t make sense was that we found pretty much the same thing in all of the fossil crocodylomorphs, including the presumably very terrestrial Terrestrisuchus and Protosuchus. With their long limbs and hinge-like joints, these two are unlikely to have been sprawlers or swimmers!
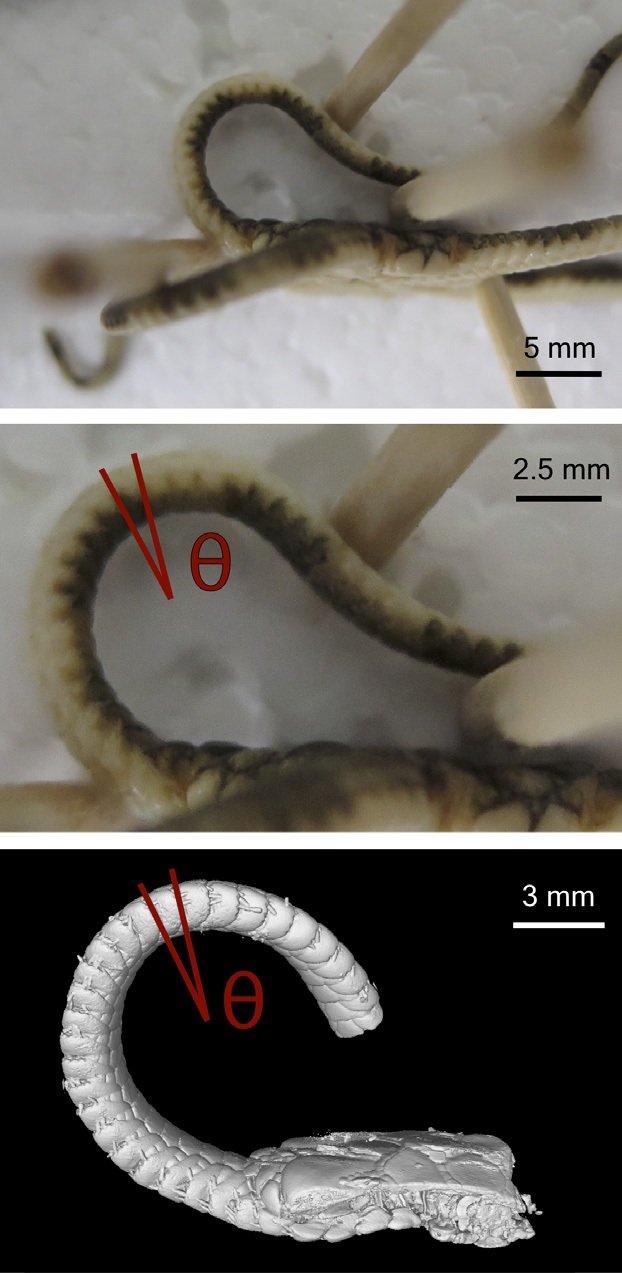
So we started looking for other parts of the croc that might affect RoM. The obvious candidate was osteoderms, the bony scales that cover the back. We went back to John’s Freezer and got out a nice frozen crocodile to measure the stiffness of its trunk and found that, sure enough, it was a lot stiffer and less mobile without the osteoderms. If the fairly flexible arrangement of osteoderms in crocodiles had this effect on stiffness, it seemed likely that (as previous authors have suggested; Eberhard Frey and Steve Salisbury being foremost amongst them) the rigid, interlocking osteoderms running from head to tail in early crocodylomorphs would really have put the brakes on their ability to move their trunk in certain ways.
Testing the stiffness of (Nile) crocodile trunks to learn the effects of osteoderms, skin, muscles, and ribs. We hung metric weights from the middle of the trunk and measured how much it flexed (Ɵ), then removed bits and repeated. Click to em-croccen.
Another cool thing we found was new evidence of convergent evolution to aquatic lifestyles in the spines of thalattosuchians. The more basal thalattosuchians, thought to have been near-shore predators, had stiffness and RoM patterns similar to Crocodylus. But Metriorhynchus, which probably was very good at chasing down fast fish in the open ocean, seems to have had greater stiffness. (The stiffness estimates come from morphometrics and are based on modern crocodiles; see here again, or just read the paper already!) A stiff vertebral column can be useful for a swimmer because it increases the body’s natural frequency of oscillation, and faster oscillation means faster swimming (think tuna, not eel). The same thing seems to have happened in other secondarily aquatic vertebrate lineages such as whales, ichthyosaurs, and mosasaurs.
So, our results were a mixed bag of adaptations particular to crocs and ones that seem like general vertebrate swimming specializations. Crocodylomorphs are important because they are the only group of large vertebrates other than mammals that has secondarily aquatic members and has living members with a reasonably similar body plan, allowing us to test hypotheses in ways that would arguably be impossible for, say, non-avian dinosaurs and birds. The take-home message: crocodylomorphs A) are awesome, and B) can teach us a lot about how vertebrates adapt to different modes of life.
I am reposting a blog post that I co-authored with Anne Osterrieder in 2012. I’ve always liked this post and been proud that we did it. A colleague brought it up to me yesterday, and I was sad to hear that the blog had been killed by hackers, with the original post lost, but Anne and I reconstructed it and I’ve decided to put it up on my blog, as I still feel strongly about its main points and Anne concurred.
Stomach-Churning Rating: 1/10; just words and ideas.
This blog is about freezer-promotion.
Here we present two views on public engagement (PE) or public relations (PR) and the thorny issue of “self promotion” in scientific research, from two scientists who might on the surface seem to be as different as scientists can be in regards to PE/PR. Yet we hope to convey the common ground that lies between these “extremes” and use it to explore, and spark discussion in, what self-promotion is and when it is a good vs. bad thing for scientists. Similar points came up in another blog post at around the same time, linked here.
Professor John R. Hutchinson (here, simply John will do!) does research on dinosaurs and elephants and other “celebrity species” (well, some of them anyway; some others aren’t so sexy but he doesn’t care). Thus getting PE/PR is often all too easy. It is often said that “dinosaur” (or fossil) is among the “holy trinity” of media story subjects; space and health being two others. That status lubricates the gears of a science PE/PR machine. Sometimes, even, the problem is keeping a lid on the “sexy” research until it is “thoroughly cooked” and ready for PE/PR, rather than releasing it prematurely. A flip side to this issue is that this easy success with PE/PR means that almost everyone is doing it, albeit with varying aplomb. So it takes some extra effort to achieve relative excellence at PE/PR in John’s line of research, but he’s not complaining. In contrast, many (indeed, most!) scientists might not have it so easy getting PE/PR and hence need to actively engage in it to draw audiences in. However, when they are successful at PE/PR it might be easier for them to then stand out from the crowd.
Dr. Anne Osterrieder (again, let’s stick with Anne for short) is a Research and Science Communication Fellow, doing research on plant cells – – hello? Hello?! Are you still there? Nine out of ten people will react to this revelation with the question: ‘Why do you work on plants? Plants are boring, they don’t really do anything, do they?’ Most plant scientists agree that the apathy or even contempt displayed towards our poor plants stems from a lack of proper engagement, starting with the way plants are taught in schools. As such plant scientists need to make a conscious effort to engage the public with current plant research and highly topical issues such as food security or plant pathology. Cells have a higher ‘fascination potential’, as the huge success of BBC’s ‘The Hidden Life of the Cell’ showed. Communicating current cell biology becomes more challenging however the deeper we go.
With those introductions done, let’s see what our two scientists think about self-promotion and PE/PR:
You might have spotted John and collaborator James Proffitt on the BBC or in the New Yorker lately, engaging in penguin-promotion.
John:
While self-promotion among scientific researchers could be a slippery slope that leads to a spiral of egomaniacal aggrandizements and delusions of grandeur, how justifiable is this seemingly common perception? In extreme instances, namely the stereotyped – but perhaps relatively rare– “media whore” or “press hound” committing the faux pas of science-by-press-release, perhaps it is. But more commonly among scientists it may just be healthy behaviour. Almost every scientist probably does research because it brings them profound joy and satisfaction, indulging their curiosity. Is it selfish to share that positive, personal message? By turning the issue around like this, one might instead wonder, what’s the problem? Put it all out there, fly your science banner high! Screw the cynics.
But as in much of life, there probably is a happy medium of moderation: a middle ground, because both selfish and generous reasons might underlie “self promotion”. Such reasons can and probably do coexist not only in perfectly non-pathological, but highly PE/PR-committed, researchers, but perhaps even in most scientists. The problem is, self-promotion has taken on bad connotations to some, or even many, scientists. It can frequently be seen couched as “shameless self-promotion” when a person promotes their science, as if to apologize for the promotion and commit it in one fell swoop. Why apologize? Just do it?! If you’re having fun with it, someone else probably will too, and that’s reason enough.
And a second issue is what kind of self-promotion is being performed– is it about the individual and their self-perceived, self-appointed glory? Or is it about the science, even in a detached third person view? Or is not even self-promotion, but team-promotion, if we consider that so many scientists these days are vital parts of a team, not lone wolves? Such a distinction of self “vs.” science is too artificial a dichotomy because scientists, as human beings, tend to feel personally enmeshed in their research. Without it, they would lack the drive to do it, even though every good supervisor is “supposed” to warn us to stay objective as researchers. And the subtext behind that “stay objective” is to stay impersonal; i.e. detached, inhuman, drained of character, passive voice and all that. Boring! But there is still some merit in considering both (and other?) sides of the matter, because it is not unreasonable to predict that the first kind of promotion (selfish; aggrandizing) is more dangerous than the second (generous; celebratory), because it is the ego taking the stage rather than the science. At the same time, we need both sides: the human, fallible, witty, emotive ego and the dry, objective, methodical, taciturn science. Without the former; warts and all; science could be too frigid to be fun.
Many researchers probably find it healthy to reflect on how much self-promotion is too much, whatever the reasons (and to some degree the reasons may not matter!). But it is not just the promoters who deserve introspection about their own practice. Those perceiving others’ “self-promotion”, especially in a negative light, could benefit from scrutiny of their own perceptions. What makes them presume that the motivation behind self-promotion is a malignant one, or not? And is the reasoning behind their judgement as sound as they’d apply to other scientific judgements they make on a daily basis– what behaviour are they reading into and how?
Alternatively, why worry about it? Isn’t a good scientist one who celebrates good science, yours, your team’s, or someone else’s? Again, this comes back to how much self-promotion is too much, but from an external perspective. Researchers are likely to judge others’ promotional activities by their own standards, not those of the promoter. They may be making value judgements with no objective basis, or (with colleagues that are not well known to the individual, all too common on the internet) no empirical evidence to go by except a brief press release, blog post, tweet or news article. Indeed, a case could be made that there is no objective basis to such a value judgement, by definition. Semantics and slippery slopes toward postmodernism aside, perhaps there is even no point to judging others’ self-promotions– and why does one wish to judge? An inward look at our own motivations for judging others’ can be salutary.
A major point here is: it is easy to conflate or confuse selfish promotion and unselfish sharing-the-joy-of-science, and to a degree it does not matter. This is because inevitably it is what is presented that matters: the content, not so much as the intent, in addition to the feedback one gets from engaging the public with research. That content-with-feedback is what almost everyone outside of academia says we should be doing—who are we to argue? Maybe we should try harder to put self esteem and other internal issues aside, and enjoy good science promotion for what it is, not what we might fear it could be. Whether a scientist is a lone wolf or team wolf, there’s no big bad wolf’s huffing and puffing to fear from good self-promotion of science. Let’s focus on building a strong house of science, brick by brick; one that lasts, and one that people hear of and care about.
Anne’s great Vacuole Song; plant organnelle-promotion!
Anne:
Whenever I write something about science communication, I feel like I am treading on an extra-slippery slope. Science communication, outreach, public engagement, PR and promotion all can have very different meanings depending on who you talk to. When I was a full-time researcher, I’d never even have thought about that they could mean different things. To me they all were synonyms of ‘Hey, let’s tell the world how amazing our research and science is!’ Since I became involved in science communication, I have realised that promoting our research isn’t necessarily the same as engaging non-expert audiences. While promotion certainly has its place and benefits (for example institutions highlighting their groups’ research achievements in external newsletter and online), real engagement is not so much broadcasting but two-way communication. I would like to point to an excellent article by Steve Cross, Head of Public Engagement at University College London in a recent issue of British Science Association magazine ‘People & Science’. Steve writes: ‘I don’t tell members of the public that ‘science is fun’ or that ‘science has the answers’. I don’t even treat science as one great big unified thing. Instead I help researchers to share what they do. The message is less ‘We’re great!’ and more ‘Here’s what we’re doing. What do you think?’
Participating in this dialogue-centred way of public engagement means however that, invariably, our specific research project will be the centre of attention. Most likely our person would be as well, since science isn’t (yet) carried out by autonomously working nano-robots. I would be very surprised if our audience saw such activities as self-promotion. I predict that they’d rather appreciate researchers ‘stepping out of the tower’ into the public and interact with non-experts. Would our peers see it as self-promotion? Probably not. What if we promoted our activities beforehand on Twitter and other online or offline channels? What if we wrote a summary of the event and reflections on it afterwards? What if we posted links to our content at different times during the day to make sure that different audiences saw it? What if we had several projects running in parallel and did this for all of them? The problem becomes apparent now and I am certain that at this point some peers would drop cynic remarks about ‘self-promotion’ or ‘attention whores’.
So, self-promotion is frowned upon. But if you think about it, our wole current academic system is based on self-promotion. When we submit a manuscript, we need to state in the cover letter why our research is novel and interesting. Even though scientific conferences are supposed to be about disseminating scientific results and initiate collaborations, they also serve the purpose of self-promotion. I don’t recall many talks with mainly negative, confusing or boring results (except maybe if a well established principal investigator was talking about their newest project and asking for feedback). Most early-career scientists would rather not submit an abstract if they haven’t got good data and wait until they can show nice results. Fact is, conferences are a big job interview for PhD students and post-docs. What about grants? Each proposal has dedicated sections for promoting yourself, your research group and your institute to increase your chances of getting a grant. Early-career researchers quickly have to learn how to write these bits, as otherwise they quickly will be at a disadvantage compared to those who can sell themselves well. I believe that there is a certain double standard around the issue of self-promotion in academia. On the one hand researchers accept it as a necessity to climb up the career ladder. On the other hand they might sneer at peers who put all of their Nature and Science references on slides in their talk. ‘What a complete showoff!’
If I follow someone on Twitter whose work I admire, say science writer Ed Yong or blogger Prof. Athene Donald, or who does cool research I am interested in, I want to read everything they publish. I appreciate them linking to their articles and papers, repeatedly, since I am bound to miss it otherwise. I loved seeing John’s BBC clip of rhino foot pressure experiments because I wanted to learn more about his research – and I loved seeing him talk about it in ‘real life’ rather than only reading his words! But if someone at my professional level, who I am competing with for fellowships or grants, was constantly posting links to their achievements, I would probably be less tolerable of them. I’d roll my eyes and think “show-off”! But I admit honestly that this would be based on a less-than-noble notions: envy, feeling threatened and insecurity about my own achievements being sufficient to succeed.
When I talked about Twitter and enhancing your online profile at our departmental Away Day someone said: “Our generation has been brought up as being humble, as not showing off, as not shouting out our achievements. So where is the border between self-promotion and being a complete d***?” I don’t think that this is a generational thing, as many senior academics have no difficulties promoting themselves. At that time I bounced the question back to the audience and asked: ‘What do the younger ones think?’ There was silence and one PhD student said: ‘I think it’s OK. You have to do it – who else would do it otherwise?’ I suspect that being willing and able to sell yourself might be a personality rather than an age thing and that the line between ‘selling yourself’ and ‘showing off’ subjectively lies in the eye of the beholder. Whatever you think, times have changed and academic positions are getting scarce. Maybe we need another motto next to ‘publish or perish’ – ‘self-promote or perish?’ Having a decent publication record won’t guarantee a research job anymore, as the competition is fierce. ‘Getting your name out there’, enhancing your profile, building a network and being engaged however will make you stand out of the crowd – as long as your self-promotion activities build upon solid achievements and not on hot air. In that case, you might deserve eye-rolling.
Self-promotion is often frowned upon in academic circles. Generally it seems to be all right to promote ‘science’ or a whole field. Numerous times I have seen blogging scientists state – defend themselves! – that in many years of writing they never blogged about their own paper. But why not? If we follow the two-way model of public engagement described above, it would be perfectly fine to write a non-expert summary about one’s latest publication and say: ‘This is what I just published, and the story behind it. What do you think?’ Similarly, the benefit of open access papers embedded in a social media site structure is that it allows discussions with non-experts. This will work significantly quicker and efficient if the authors alert and direct potential audiences to their paper through as many communication channels as possible- an act that again can be seen as self-promotion. Is our academic culture with its subtle or open contempt of self-promotion maybe inadvertently hindering effective engagement?
What do you think? Chime in on the poll below.
If the poll does not show up above in your browser, click the link here to go directly to it (new window):
Some context, first. As we finished this post together, Anne and John reflected on what got us working on it, back in August 2012:
Anne: “You wrote that you had these thoughts on self-promotion after you returned from the [British] Science Festival. Was there a specific incident that raised these thoughts, or just general thinking?
John: “I often think about what I tweet and the amount of it, and whether “me-tweeting” is such a bad thing as some on Twitter say it is. I was me-tweeting a bunch of responses to my BSF talk and I thought I should, much as I do the same when people post stories about my research papers etc. But in particular this BSF event, which was heavy PE, got me thinking on the train ride home about why some people would (cynically, in my view) see that as PR and shameful self-promotion.”
While the two views we presented above are from different backgrounds and perspectives and such, our thoughts reveal many elements common to both. Perhaps these commonalities apply to most scientists, but, but… There is a hulking science-gorilla in the room: cultural similarities and differences. We cannot neglect the HUGE issue of Western scientific culture that John and Anne and others have in common! In other cultures, self-promotion might be seen very differently; indeed in UK it seems to sneered at more than in the USA, as Brits tend to be less comfortable tooting their own horn (easy, now!). Some other cultures might have no problem with it at all. Others might find it abominable. However, how culture factors into self-promotion and PE/PR perceptions is a huge kettle of fish that we’re not quite ready to tackle, so we will turn that over for discussion in the comments here! How does your culture, whether very local (department?) or very broad (country/ethnicity) factor into this?
Or, if you prefer, please contribute your thoughts on how you handle or perceive the self-promotion vs. science-promotion (false) dichotomy as a scientist, science communicator and/or layperson? How do you determine what is a tolerable level of promotion?
Stomach-Churning Rating: 1/10 for ambiguous sacks.
I mainly post here about my team’s research and interests, but today I felt like sharing something special and concrete: the contents of our freezers. They are not just John’s and there’s more than one freezer; thus there is room to share, within reason. So if you’re a researcher, especially in the UK/EU, needing unusual research specimens/tissue, you might want to contact me to use them. This blog’s posts summarize most of what I have available, and for security/other reasons I don’t want to get into deep detail here, but we sport a respectable collection of limbs/bodies of animals like:
Birds: ostriches, emus, broiler chickens, guineafowl, assortment of others.
Crocodiles: Nile, Osteolaemus, Morelet’s and some others (1 Melanosuchus, 1 normal Caiman).
Squamates: a monitor lizard or two and some other random lizards.
Amphibians: a few fire salamanders and such.
Mammals: of course, plenty of elephant bits (no ivory!), rhinos too (no horns!), giraffes, a dwarf forest buffalo, alpacas, deer, pieces of camels and zebras (feet etc.), wild cat species (no penises!) and a few other things. And then the usual assortment of veterinary species like cows and horses. A heavy focus on limb material– very few if any heads, torsos, etc.
This is in addition to a nice little comparative skeletal collection, focused on cleaned members of the above groups and a smattering of others. Nothing on the scale of RVC’s marvellous Anatomy Museum, but we’re young.
And two African land snail shells (inhabited) I was reminded of during a recent inventory… Here are some of my helpful helpers in that inventory extravaganza!
Especially if you’re searching for CT scan data (sooner or later these data will appear online; I want it to happen!), tissue samples for genetics or cell biology (if frozen is OK!), comparative anatomical specimens to inspect, or other uses of frozen anatomy (photography? other art? We’ve helped artists before!), the freezers might be able to help you! The less destructive, the better, but even some destructive analysis might be OK. We regularly accommodate visitors, either independent ones or collaborators, and I aim to provide good hospitality when I can accommodate them!
Get in touch with me if the above description is you. It’s not an open invitation to everyone, but for valid research purposes I can and should try to help. But I’m limited by time and other human factors, so I can’t do everything and help everyone. Our ability to host others to come work on our specimens here in-house is very limited, I’m sorry to say. The primary purpose of all the hard work we’ve done accumulating these specimens remains to support our research, but there’s room to help others too, and we want to maximize the impact of our research collection, including potentially on teaching and public engagement with science where feasible. So I’ve put it out there, and that ends this post.
UPDATE May 2016:
I am planning more freezer cleaning; under more pressure due to dwindling space; so if specimens here interest you and you’re EU-based, I am possibly up for loaning out material. Conditions:(1) you do all transport pickup and return; (2) skeletal material is kept intact unless I approve in advance; (3) 2 year-ish loan at most, informal. We are curating material, so it must return– and it must stay in the EU (ideally UK) if on loan. Everything gets CT-scanned before it goes out. Not everything is up for grabs, but there is room for negotiation as to what’s available or not. For scientific usage only! Best to email me if interested.
Is there something in the “Non-Elephant Freezer” for you?
(John: here’s a guest post from my former PhD student, soon to be 100% legit PhD, Dr., and all that jazz, Julia Molnar!)
This is my first guest post, but I have been avidly following what’s in John’s freezer (and the blog too) for quite a while. I joined the lab in 2009 and left a month ago on the bittersweet occasion of surviving my PhD viva (oral exam/defense), so I’d like to take a moment here to thank John and the Structure & Motion Lab for a great 4 years!
Moving on to freezer-related matters; specifically, a bunch of frozen crocodile spines. It was late 2011, and the reason for the spines in John’s freezer was that John, Stephanie Pierce, and I were trying to find out more about crocodile locomotion. This was anticipated to become my first major, first-author research publication (but see my Palaeontologia Electronica paper on a related subject), and I was about to find out that these things seldom go as planned; for example, the article would not be published for more than three years (the research took a long time!). Before telling the story of how it lurched and stumbled toward eventual publication, I’ll give you some background on the project.
Stomach-Churning Rating: 3/10; x-ray of dead bits and nothing much worse.
A stumbly sort-of-bounding crocodile. They can do better.
First of all, why crocodiles? For one thing, they’re large, semi-terrestrial animals, but they use more sprawling postures than typical mammals. Along with alligators and gharials, they are the only living representatives of Crocodylomorpha, a 200+ million year-old lineage that includes wolf-like terrestrial carnivores, fish-like giants with flippers and a tail fin, even armored armadillo-like burrowers. Finally, crocodiles are interesting in their own right because they use a wide variety of gaits, including bounding and galloping, which are otherwise known only in mammals.
Nile crocodile skeletal anatomy
OK, so why spines? Understanding how the vertebral column works is crucial to understanding locomotion and body support on land, and inter-vertebral joint stiffness (how much the joints of the backbone resist forces that would move them in certain directions) in particular has been linked to trunk movements in other animals. For this reason, vertebral morphology is often used to infer functional information about extinct animals, including dinosaurs. However, vertebral form-function relationships have seldom been experimentally tested, and tests on non-mammals are particularly scarce. So we thought the crocodile spines might be able to tell us more about the relationship between vertebral morphology, mechanics, and locomotion in a broader sample of vertebrate animals. If crocodile spine morphology could be used to predict joint stiffness, then morphological measurements of extinct crocodile relatives would have some more empirical heft to them. Several skeletal features seem to play roles such as levers to mechanically stiffen crocodile spines (click to emcroc’en):
Anatomy of a crocodile vertebra
We decided to use a very simple technique that could be replicated in any lab to measure passive stiffness in crocodile cadavers. We dissected out individual joints were and loaded with known weights. From the movement of the vertebrae and the distance from the joint, we calculated how much force takes to move the joint a certain number of degrees (i.e. stiffness).
Me with crocodile vertebra and G-clamp
X-ray of two crocodile vertebrae loaded with a metric weight to calculate their joint’s stiffness
Afterwards, we boiled the joints to remove the soft tissues – the smell was indescribable! We took 14 measurements from each vertebra. All of these measurements had been associated with stiffness or range of motion in other studies, so we thought they might be correlated with stiffness in crocodiles also.
Some of the vertebral measurements that were related to stiffness
Despite my efforts to keep it simple, the process of data collection and analysis was anything but. I recall and exchange with Stephanie Pierce that went something like this:
Stephanie: “How’s it going?”
Me: “Well, the data are messy, I’m not seeing the trends I expected, and everything’s taking twice as long as it was supposed to.”
Stephanie: “Yes, that sounds like science.”
That was the biggest lesson for me: going into the project, I had been unprepared for the amount of bumbling around and re-thinking of methods when the results were coming up implausible or surprising. In this case there were a couple of cool surprises: for one thing, crocodiles turn out to have a very different pattern of inter-vertebral joint stiffness than typical mammals: while mammals have stiff thoracic joints and mobile lumbar joints, crocodiles have stiffer lumbar joints. Many mammals use large lumbar movements during bounding and galloping, so crocodiles must use different axial mechanics than mammals, even during similar gaits. While that’s not shocking (they did evolve their galloping and bounding gaits, and associated anatomy, totally independently), it is neat that this result came out so clearly. Another unexpected result was that, although several of our vertebral measurements were correlated with stiffness, some of the best predictors of stiffness in mammals from previous studies were not correlated with stiffness in crocodiles. The study tells a cautionary tale about making assumptions about extinct animals using data from only a subset of their living relatives or intuitive ideas about form and function.
Finally, the experience of doing the experiments and writing the paper got me interested in other aspects of crocodilian functional anatomy. For instance, how does joint stiffness interact with other factors, such as muscle activity and properties of the ribs, skin, and armor in living crocodiles? Previous studies by Frey and Salisbury had commented on this, but the influence of those factors is less tractable to experiment on or model than just naked backbones with passively stiff joints. In the future, I’d like to study vertebral movements during locomotion in crocodiles – especially during bounding and galloping – to find out how these patterns of stiffness relate to movement. In the meantime, our study shows that, to a degree, crocodile backbone dimensions do give some clues about joint stiffness and locomotor function.
To find out more, read the paper! It was just featured in Inside JEB.
Julia Molnar, Stephanie Pierce, John Hutchinson (2014). An experimental and morphometric test of the relationship between vertebral morphology and joint stiffness in Nile crocodiles (Crocodylus niloticus). The Journal of Experimental Biology 217, 757-768 link here and journal’s “Inside JEB” story
Today, to help thaw you poor Americans out of that Arctic Vortex, we have a guest post bringing the heat, by my PhD student Sophie Regnault! This relates to some old posts about rhinos, which are a mainstay here at the WIJF blog- I’ve posted a lot about the rhino extinction crisis, feet, skin, big and bigger bones, and more, but this is our first rhinoceros-focused, actual published scientific paper! Take it away, Sophie! (We’re planning a few more “guest” blog posts from my team, so enjoy it, folks!)
Almost a year ago to the day, I submitted my first paper written with John Hutchinson and Renate Weller at the RVC and it has (finally!) just been published. To celebrate, I have been allowed to temporarily hijack ‘What’s in John’s Freezer?’ for my first foray into the world of blogging. I started the paper back as an undergraduate veterinary student. It was my first experience of proper research, and so enjoyable that I’m now doing a PhD, studying sesamoid bones like the patella!
We wanted to discover more about the types of bony disease rhinos get in their feet, of which there isn’t much known. Rhinos, of course, are big, potentially dangerous animals – difficult enough to examine and doubly difficult to x-ray clearly because of their thick skin. Unlike diseases which are fairly easy to spot (like abscesses or splitting of the nails and footpad), there is hardly anything out there in the scientific literature on bony diseases in rhino feet. It’s no small issue, either. When your feet each need to support over 900kg (typical for a large white rhino), even a relatively minor problem can be a major pain. Progressing unseen under their tough hide, lesions in the bone can eventually become so serious than the only solution is euthanasia, but even mild conditions can have negative consequences. For example, foot problems in other animals are known to have knock-on effects on fertility, which would be a big deal for programs trying to breed these species in captivity.
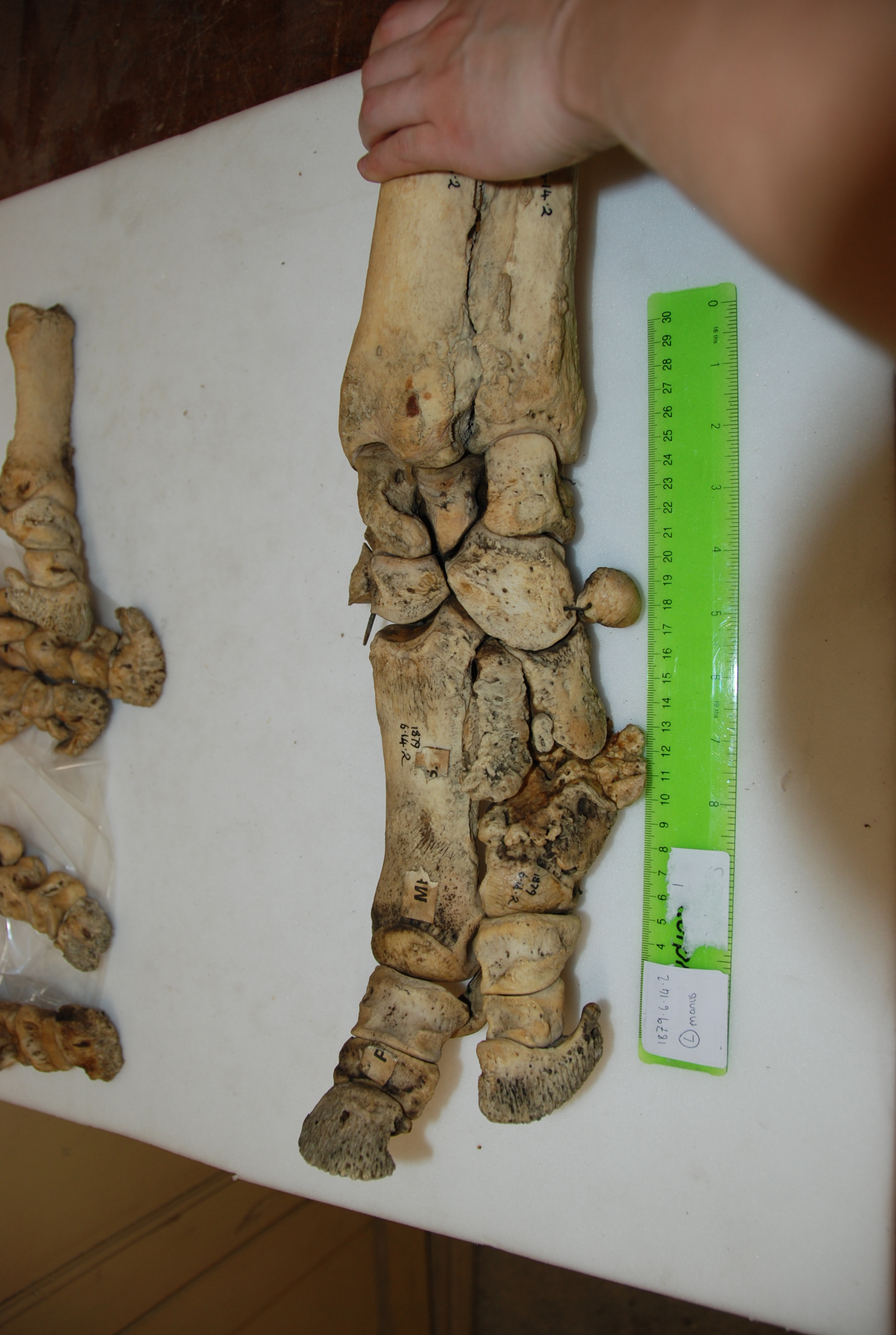
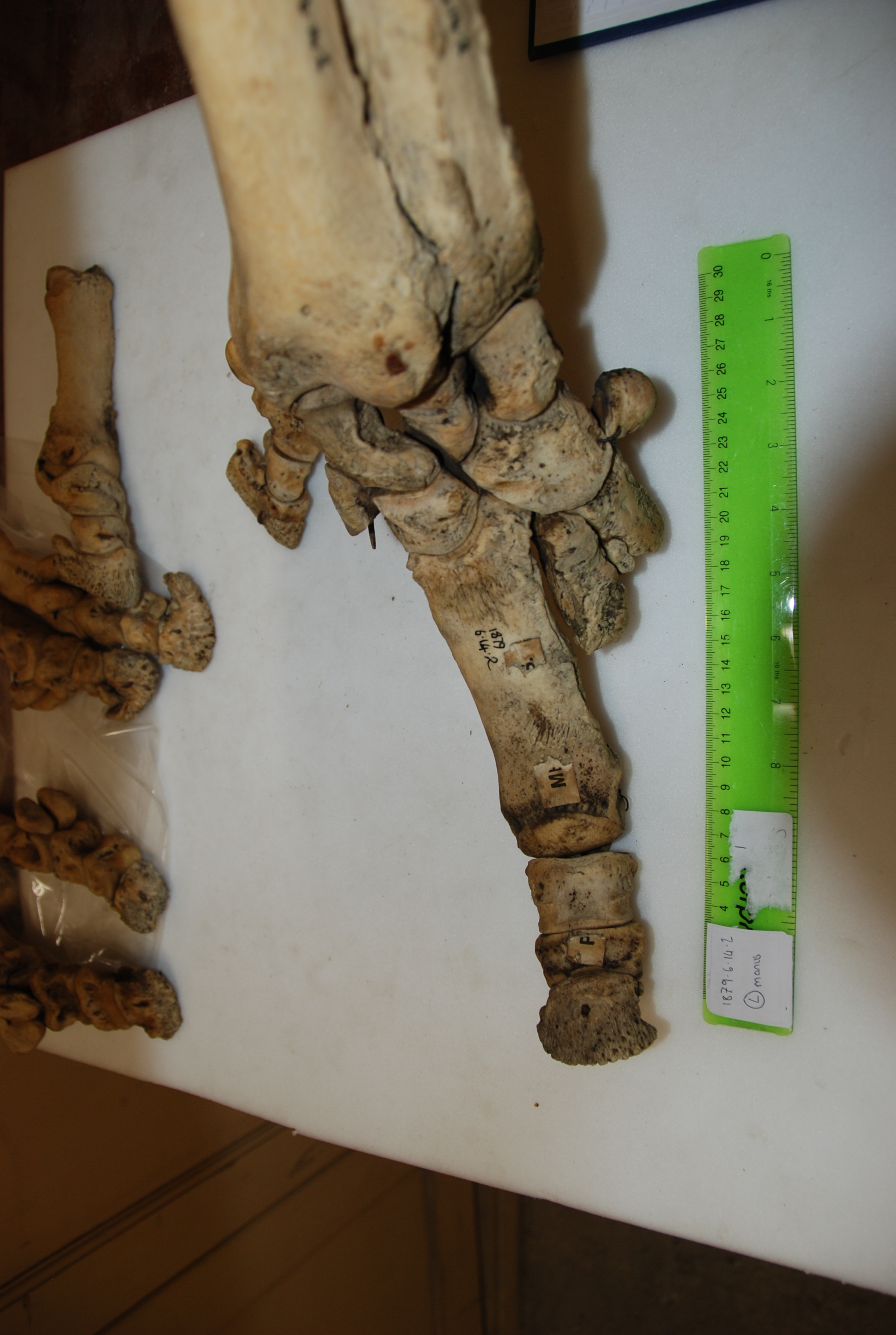
Hidden treasures abound! (Photos can be clicked to embiggen)
Data gathering was a blast. I got to travel to Cambridge, Oxford, and London during one of England’s better summers, and these beautiful old museums were letting me snoop around their skeleton collections. I’d been there often as a visitor, but it was anatomy-nerd-heaven to go behind the scenes at the Natural History Museum, and to be left alone with drawers and drawers of fantastic old bones. Some of the specimens hadn’t been touched for decades – at Cambridge University Museum of Zoology, we opened an old biscuit tin filled with the smallest rhinoceros foot bones, only to realise they were wrapped in perfectly preserved 1940’s wartime Britain newspaper.
Osteomyelitis… (3 clickable pics above) the toe’s probably not meant to come off like that!
In addition to my museum studies, I had another fun opportunity to do hands-on research. John (of course!) had freezers full of rhino legs (looking disconcertingly like doner kebabs, but maybe that’s just me!), which we CT scanned to see the bones. Although it is a pretty standard imaging technique, at this point I had only just started my clinical studies at the vet hospital, and being able to flick through CT scans felt super badass. Most vet students just get to see some horse feet or dog/cat scans, at best.
Another osteomyelitis fracture, visible in a CT scan reconstruction.
We expected to find diseases like osteoarthritis (a degenerative joint disease) and osteomyelitis (bone infection and inflammation). Both had previously been reported in rhinoceroses, although it was interesting that we saw three cases of osteomyelitis in only 27 rhinos, perhaps making it a fairly common complication. It’s an ugly-looking disease, and in two of the cases led to the fat, fluffy bones fracturing apart.
We also had several unexpected findings, like flakes of fractured bone, mild dislocations, tons of enthesiophytes (bone depositions at tendon/ligament attachments) and lots of holes in the bones (usually small, occasionally massive). For me, writing up some of these findings was cool and freaky paranoid in equal measures. They hadn’t been much described before, and we were unsure of their significance. Was it normal, or pathological? Were we interpreting it correctly? Discussions with John and Renate (often involving cake) were reassuring, as was the realisation that in science (unlike vet school at the time, where every question seemed to have a concrete answer) you can never be 100% sure of things. Our study has a few important limitations, but has addressed a gap in the field and found some neat new things. Six months into my PhD, I’m enjoying research more than ever, and hoping that this paper will be the first of many (though I promise I won’t keep nicking John’s blog for my own shameless self-promotion if that happens! EDIT BY JOHN: Please do!).
Nasty osteoarthritis wearing away the bone at the joint surface. Most cases occurred in the most distal joint.
Deep holes in some of the bones: infection, injury?
The paper:
Sophie Regnault, Robert Hermes, Thomas Hildebrandt, John Hutchinson, and Renate Weller (2013) OSTEOPATHOLOGY IN THE FEET OF RHINOCEROSES: LESION TYPE AND DISTRIBUTION. Journal of Zoo and Wildlife Medicine: December 2013, Vol. 44, No. 4, pp. 918-927.