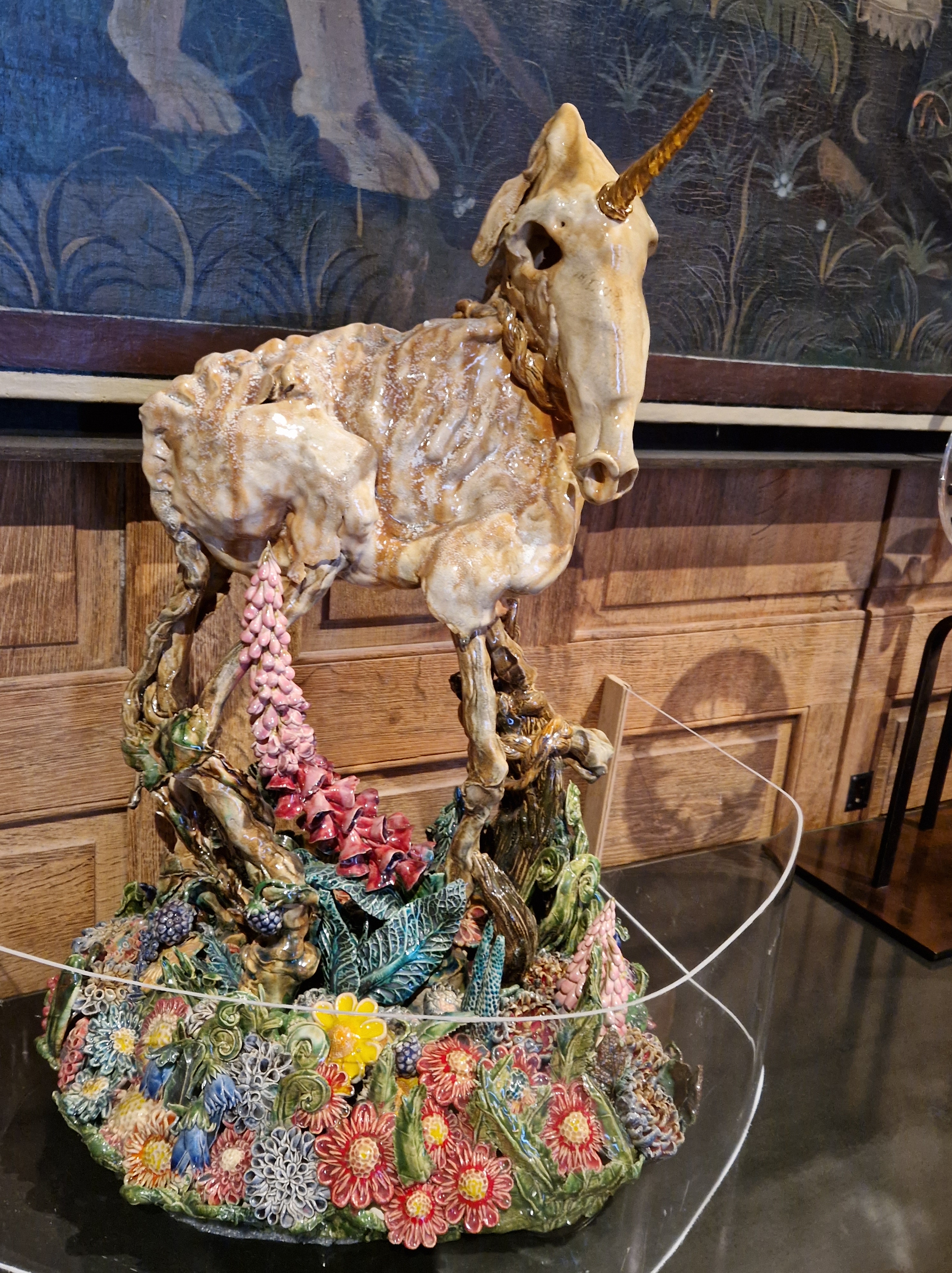I’ve been in Paris twice recently, for PhD defenses of co-supervised students, and had a great time. Which included doing one of my favourite things, wandering that amazing city and exploring its streets — and museums! I’ve raved about the perfection of la Galerie de Paléontologie et d’Anatomie comparée before, and that love will never die. But this time, in one of my wanderings I stumbled across a museum that surprised me with how fun it is. (I also got to explore the sewers, which I tweeted about in a thread here.) This is post what it’s about. Désolé, I cannot write this in French.
Stomach-Churning Rating: 3/10; I don’t know what to say, it’s bizarre stuff. But not gory, per se.
This is Le Musée de la Chasse et de la Nature à Paris. The name sounds innocent enough, except for the hunting (chasse) poor beasties part. Tripadvisor said it was a good overlooked destination, so I thought OK, it probably just has some stuffy Victorian-style rooms with dusty old hunting trophies. It had plenty of that, but so, so much more! This museum trip was TRIPPY.
First, there was a special gallery of ceramics, which caught me off guard as I expected boring, fussy relics that my grandparents might have kept in a cabinet. Oh no grandpa and grandma, these would not be for you, not at all. The stuff of nightmares!
As usual, click to embiggen images– and I recommend this, to fully savour their delights!
You could play a scavenger hunt throughout the rest of the galleries, spotting more of these tucked amongst the other displays:
Then at the back of that special gallery there was a fresco-like display, it seemed made out of paper (or was it ceramics like paper?) and it took my breath away. Tons of detail and ghoulish creativity.
Next, lots of just surreal, wacky, disturbing, WTF-why-is-this-here stuff, almost randomly distributed amongst more staid (or sometimes rather naughty/disturbing) artworks and taxidermy of hunting glories as I walked through the labyrinth of galleries:
Like, why a boar festooned with male pheasants?
It’s a hunting museum, and when it went in for hunting imagery and artifacts, it went in with all guns blazing:
And it wasn’t shy about the human figure, either:
Some highlights for me: (1) canned critters!
(2) Kissing apes!
(3) bear… shoes…
How about a 3D view of a weird sculpture of snaky tentacles and… somebody.
As a closer, let’s laugh our way out of the museum. Turn your volume up and have a nice, friendly, entirely not creepy chat with the boar’s head. Remember a certain scene in “Evil Dead 2”? Well I do! One of the greatest scenes in cinema ever, IMO. The boar would fit right in.
Until we meet again, go to the museum if you’re in Paris, and keep up the good fight.
If you go into central Lausanne, Switzerland, you’re likely to pass the Palais du Remine, and if you do, I recommend you go inside. I was happy I did while visiting Lausanne for the AMAM2019 conference. A luxurious palace has been given over to house five (!) free (!) museums on science and culture. These include the canton’s (~state’s) museums of palaeontology and zoology, which I’ll showcase here (also a little of geology and archaeology museums). Tripadvisor’s reviews were good but not as glowing as I’d make mine, so I will remedy that. I’m a sucka for old-school museums, and that’s what these are. So if that sounds right for you, journey onward!
It’s nice.
As you may be expecting by now if you’ve been here before, it’s time for another museum photo blog!
Stomach-Churning Rating: 5/10 for bones, preserved organs, taxidermy aplenty, and animal developmental deformities.
Nice cathedral nearby, w/great view of the city.
Nice interior architecture. There’s lots of nice to behold.
Posters That Get You Excited 101. But you must wait. Like I did.
Quadrupedal human at Zoology museum entry.
Tomistoma, false gharial.
Not a bad collection of taxidermied Crocodylia!
Visually arresting cobra display.
I’ve never seen three Draco gliding lizards on display together!
Bipedal lizard taxidermy displays, freezing the dynamic in the static, are no easy feat.
Plenty of stuffed animals like these raptors/other large birds. Classical zoology museum style. Minimal signage. Just specimen labels, mainly.
Coelacanth!
Sperm whale jaw.
Open space with big specimens. A ~4m long great white shark included.
Second zoology hall: bones!
Gorilla standing tall next to human.
Ostrich skeleton up close, amongst the mammalz.
Cassowary skeleton.
Emu shoulder/arm bones in right side view.
Walrus skeleton in what seems like an odd pose to me, but then they are odd on land.
Alligator skeleton in repose.
Giant anteater, “knuckle-walking”.
Pangolin skeleton! And mounted digging into a nest– very well done!
Bernard Heuvelmans display, about the (in)famous cryptozoologist. This was quite a surprise to me. I’m sure I’d read his English-translated book “On the Track of Unknown Animals” as a kid, during my long stint as an avid reader of much zoology, crypto- and otherwise. He bequeathed a lot of his work to the museum.
Bernard’s handwritten CV!? With a “sea serpent” sketch.
A “sea serpent” vertebra… but if you know any anatomy, it’s not a snake’s vertebra at all but a fish’s, such as a basking shark‘s.
Are you ready for more weirdness? How about some “mutants”- congenital deformities of animals? Fascinating errors of developmental anatomy… somehow this two-headed calf survived awhile. Plenty more where that came from, as follows:
And then there’s all kinds of wonderful comparative anatomy. To be a student of this subject in Lausanne would be a lucky thing, with this museum’s collection at hand. These are valuable specimens, made with love and skill.
Jaws
Fish head anatomy. Some vertebrae on the left, too.
Developmental regions of the head: a lovely wax(?) model of an Echidna skull. A treasure.
Brains: alligator vs. pigeon.
Salamander muscles.
Pigeon muscles.
More spotted felids than you can shake a jar of catnip at.
Another pangolin!
Giant armadillo.
Petaurus: flying phalanger (a gliding marsupial).
Second zoology hall open area: left side.
Second zoology hall open area: right side.
A final hall with a more new-fangled display, on the topic of evolution and extinction. Attractive phylogeny graphic here. Birds at the “top”, of course. Poor lowly mammals!
Taxidermied giant auk- not a common sight! (Extinct)
The extinct southern pig-footed bandicoot. Also a rare sight of a whole specimen- in a Swiss museum, too.
NOW ON TO THE FOSSILS!
You’ve been very patient. Here, have a Toblerone.
Palaeo museum entry. Already there are cool things visible. Inside, we find it just like I prefer my zoo/palaeo museums (as above): stuffed with specimens and leaving plenty for you to wonder about and investigate. Not frilly; a well-stocked museum that mostly lets its specimens speak for themselves.
Sauriermuseum (Aathal) specimen of Plateosaurus: sculpt/cast. A very good, big skeleton of this common dinosaur, rearing up.
Rear view of same.
Real bones of same; vertebrae and pelvic (this is the “Frick specimen”).
Metaxytherium (current name), an ancient and large fossil dugong/seacow. Skull is in left side view. (that may help, as their skulls are odd!)
Anthracotherium upper jaw: ancient hippo-cousin.
Prolagus: the “Sardinian hare” (recently extinct; old lineage).
Potamotherium: to some an early otter-like mammal, more recently thought to be an ancient seal.
“Broke-ulum”: a walrus broke its penis bone (baculum) and was surely not pleased about it, but lived to heal— physically if not mentally. Yeesh!
Glyptodont tail club and armour.
Aepyornis elephant bird legs!
A partial/reconstructed skeleton of the dodo.
Velociraptor preparing to pounce from above. It’s too late for you!
Rhamphorhynchus fossil (2D slab) and sculpt/cast coming alive in 3D– good stuff.
Anhanguera pterosaur watches the chaos from above, fish snagged in its teeth.
Not-shabby metriorhynchid marine croc fossils, from Britain.
Lovely 3D plesiosaur bones (flippers, neck, etc.) from near RVC: Peterborough!
Mesosaur; early reptile.
The museum clearly is proud of its excellent “Mammoth of Brassus” skeleton, essentially complete.
Ice Age elk/moose, a 10,000 year old skeleton in fine shape.
Cave bear skull rawr
Purty ammonites!
Spiky ammonite!
Cretaceous sponge colony from France. I hadn’t seen something like this before, so here it is.
Trilobites, brittlestars and friends.
Well I did wander through the geology and archaeology museums too, and while I liked them I did not take so many photos. My non-human organismal bias is apparent. But check these final ones out:
Splendid cross-section of the stratigraphy of the Alps around Lausanne. I gazed at this for quite a few minutes, trying to figure out what was where in the landscape I’d seen and how old, how deformed, etc.
Slab of “dinosaur” tracks but it was not clear to me what dinosaurs/archosaurs/whatever made them. I wish my French was better. Closeup below shows two footprints superimposed.
At last, the coup de grace! What museum would be complete without a diorama!? (I love them) This one, with a goat sacrifice and early Stone Age people praying to heathen deities/spirits at an elaborate petroglyph array rocked my world. And so it makes a perfect final image. Enjoy, and conduct the proper rites. \m/
I heard that the UMZC has some new exhibits open, so back I went! For the prior posts see here (mammals/basement) and here (everything else). Another photo tour! There’s a special (art) exhibit, too, so stick around to the end.
All images can be clicked to mu-zoom in on them.
Stomach-Churning Rating: 3/10 mainly skeletons, some preserved critters in jars.
The first new section is an elaborated display on reptiles.
Clevosaurus, a Triassic relative of the living tuatara reptile, Sphenodon. Nice fossil hindlimbs!
Tuataras (Sphenodon), skeletal and preserved.
Tuatara embryos!
Nice chameleon mount w/tongue extended.
Thorny devil (Moloch), de-thorned and in the flesh.
Skull (cast) of Ninjemys, the giant turtle.
Pipe snakes! Snakes with vestigial hindlegs.
Istiodactylus pterosaur snout-tip (real fossil) from the Isle of Wight, UK. Nice 3D fossil.
The gharial (Gavialis), male with protuberance on snout (mating-related).
I dub thee Dinosaur Corner! For dinosaurs, the Sedgwick Museum across the street (also free; also classic and awesome) is the place to go but this corner does a good job fighting for the scientific conclusion that birds are dinosaurs.
And now a change of pace. On to the special exhibit!
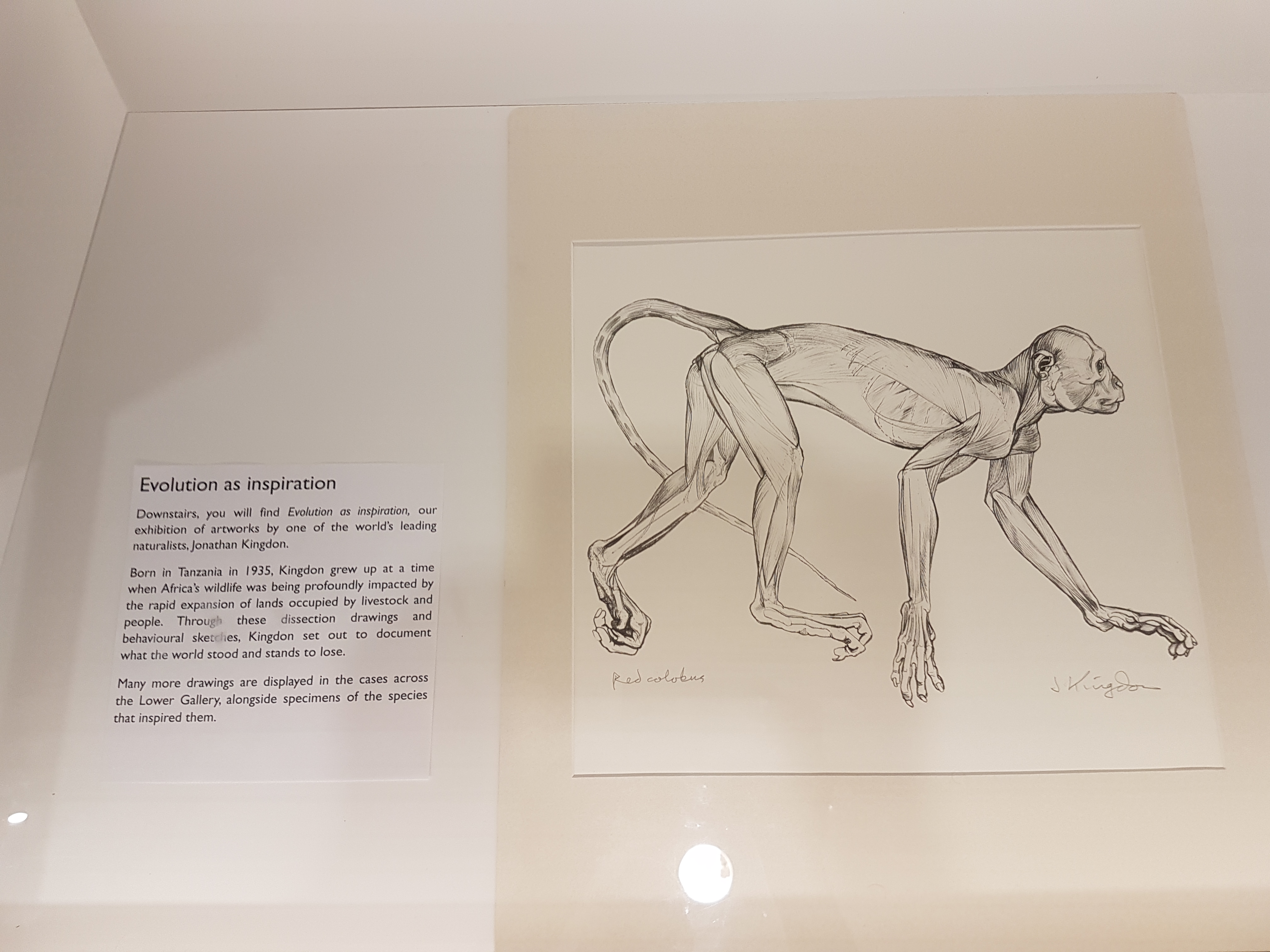
A nice surprise to see naturalist superstar Jonathan Kingdon‘s scientific illustrations and nature-inspired artwork displayed here. I’ve added photos of ones I liked the most.
As the caption explains, Kingdon used art to explain the value of nature; via realistic images of life, dissections, and creative abstractions drawn from them.
Hammerhead bats: even freakier when skinned.
Begone if ye find not joy in aardvarks!
White-toothed shrew looking extra-ghoulish with flensed face.
Skinned sengis in action.
More sengis (elephant shrews); with a note explaining that they are not rodents/insectivores but afrotheres, cousins of aardvarks, elephants and kin.
Bronze Jackson’s chameleon bust.
Asian barbet faces: this was fascinating. Kingdon used the paintings to explain how barbet faces vary across species as recognition devices to aid in territorial defense, especially of their nest-holes in trees, in which they face outwards to display their coloured faces. The middle image shows one lone species that has no such territorial competitors and has evolved back into brown colour, perhaps due to relaxed selective pressure for colour. Neat!
Oh my, this took my breath away! Mixed media depicting the varied forms of facial ornaments in vultures; soft tissues used in communcation. And here mounted on a butcher’s rack. Do vulture bits mimic their grisly food?
To me, there is no question that the Galerie de Paléontologie et d’Anatomie comparée of Paris’s Muséum national d’Histoire naturelle (MNHN) is the mecca of organismal anatomy, as their homepage describes. Georges Cuvier got the morphological ball rolling there and numerous luminaries were in various ways associated with it too; Buffon and Lamarck and St Hiliaire to name but a few early ones. It is easy to think of other contenders such as the NHMUK in London (i.e., Owen), Jena in Germany, the MCZ at Harvard (e.g. Romer) and so forth. But they don’t quite cut the dijon.
As today is John’s Freezer’s 7th blogoversary, and I was just at the MNHN in Paris snapping photos of their mecca, it’s time for an overdue homage to the magnificent mustard of that maison du morphologie. The exhibits have little signage and are an eclectic mix of specimens, but this adds to its appeal and eccentricity for me. I’ve chosen some of my favourite things I saw on exhibit on this visit, with a focus on things that get less attention (NO MESOZOIC DINOSAURS! sorry), are just odd, or otherwise caught my fancy. It’s a photo blog post, so I shall shut up now, much as I could gush about this place. I could live here.
Need plus-grand images? Clic!
Stomach-Churning Rating: 7/10 for some potentially disturbing anatomical images such as viscera, preserved bits, models of naughty bits etc.
Greetings. Note the stomach-churning rating above, please.
Right. We’ll get the amazing first view as one steps into the gallery done first. Mucho mecca. Anatomy fans simply must go here at least once in their life to experience it, and one cannot ever truly absorb all the history and profound, abundant details of morphology on exhibit.
Less-often-seen views from the balcony; one more below.
Indian Rhinoceros from Versailles’s royal menagerie; came to the MNHN in 1792.
Brown bear hindlimb bones.
Brown bear forelimb bones and pelvis.
Two baby polar bears; part of the extensive display of ontogeny (too often missing in other museums’ exhibits).
Asian elephant from Sri Lanka.
Lamb birth defect. Like ontogeny, pathology was a major research interest in the original MNHN days.
Wild boar birth defect.
Fabulous large Indian gharial skull + skeleton.
“Exploded” Nile crocodile skull to show major bones.
Let’s play name-all-the-fish-skull-bones, shall we?
Rare sight of a well-prepared Mola mola ocean sunfish skeleton.
Diversity of large bird eggs.
Asian musk deer (male), with tooth roots exposed.
Freaky gorilla is here to say that now the really odd specimens begin, including the squishy bits.
Freaky tamandua, to keep freaky gorilla company. Displaying salivary glands associated with the tongue/pharynx. These are examples of anatomical preparations using older analogues of plastination, such as papier-mâché modelling. I’m not completely sure how the preservation was done here.
Simpler stomach of a wolf. Not much room for Little Red Riding Hood, I’m afraid.
Expansive surface area of a hippo’s stomach; but not a multi-chambered ruminant gut.
Cervical air sacs of a Turquoise-fronted Amazon parrot.
Heart and rather complex pulmonary system of a varanid lizard.
It’s pharynx time: Keratinous spines of a sea turtle’s throat. All the better to grip squids or jellies!
Pharynx convergent evolution in a giraffe: keratinous spines to help grip food and protect the pharynx from spiny acacia thorns while it passes down the long throat.
Tongue/hyoid region of the pharynx of a varanid, showing the forked tongue mechanism.
Palaeontological awesomeness on the upper floor (the 2nd part of the gallery’s name). Here, the only Siberian woolly mammoth, I’m told, to have left Russia for permanent display like this. Frozen left side of face, here, and 2 more parts below.
Extinct rhino Diaceratherium, with a pathological ankle (degenerative joint disease). I love spotting pathologies in specimens- it makes them stand out more as individuals that lived a unique life.
Glyptodont butt and thagomizer, to begin our tour of this business-end weaponry.
Eutatus leg bones, from a large fossil armadillo; Argentina. Really odd morphology; Xenarthrans are so cool.
Stomach-Churning Rating: 10/10 may be too high (it’s all plastinated anatomy; not gooey bloody stuff) but I’m being wary. There are graphic images of humanity and opinions will vary on the tastefulness; I think they are beautiful. (And to me, Bodyworlds plastination leaves specimens looking more like puppets or statues than disturbing undead) There are images of reproductive anatomy that are not appropriate for children unless parental guidance is along for a “birds and the bees” chat. Got it? OK.
I had a spare hour in Cambridge this weekend so I dared the crowds in the revamped UMZC’s upper floor. In my prior visit and post I’d experienced and described the lower floor, which is almost exclusively mammals. This “new” floor has everything else that is zoological (animal/Metazoa) and again is organized in an evolutionary context. And here is my photo tour as promised!
Inviting, soft lighting perfuses the exhibits from the entryway onwards.
All images can be clicked to mu-zoom in on them.
Stomach-Churning Rating: 5/10 for spirit animals, by which I mean dissected/ghostly pale whole specimens of animals in preservative fluids.
The exhibits are on a square balcony overlooking the lower floor, so you can get some nice views. It does make the balcony crowded when the museum is busy, so take that in mind if visiting. Strollers on this upper floor could be really difficult. But the ceiling is very tall so it is not cramped in a 3D sense. The lower floor is more spacious.
Like phylogenies? You got em! Tucked away at the beginning of each major group; not occupying huge valuable space or glaringly obvious like AMNH in NYC but still noticeable and useful. To me, it strikes a good balance; gives the necessary evolutionary context for the displayed specimens/taxa.
Introductory panels explain how names are given to specimens, how specimens are preserved and more.
The exhibits give due focus to research that the UMZC is doing or has been famous for. Hey I recognize that 3D tetrapod image in the lower left! 🙂
There is ample coverage of diversity throughout Metazoa but my camera tended to be drawn to the Vertebrata. Except in some instances like these.
Some larger chelicerates.
Some smaller, shadowy sea scorpion (eurypterid) fossils.
Watch here for more about ophiuroids (brittlestars) in not too long!
A BIG fish brain! Interesting! Before I go through specimens in evolutionary “sequence”, I will feature another thing i really liked: lots of dissected spirit-specimens that show off cool anatomy/evolution/adaptation (and technical skills in anatomical preparation). Mostly heads; mostly fish.
Salps and other tunicates! Our closest non-vertebrate relatives- and some insight into how our head and gut came to be.
Salp-reflection.
Lamprey head: not hard to spot the commonalities with the salps; but now into Vertebrata.
Hagfish head: as a fellow cyclostome/agnathan, much like a lamprey but never forget the slime glands!
Shark head. Big fat jaws; all the better to bite prey with!
Lungfish (Protopterus) head showing the big crushing tooth plates (above).
Sturgeon vertebrae: tweak some agnathan/shark bits and here you are.
Worm (annelid) anatomy model, displaying some differences from/similarities to Vertebrata. (e.g. ventral vs. dorsal nerve cord; segmentation)
Dissected flipper from a small whale/other cetacean. Still five fingers, but other specializations make it work underwater.
Wonderful diversity of tooth and jaw forms in sharks, rays and relatives. I like this display a lot.
More of the above, but disparate fossil forms!
On with the evolutionary context! Woven throughout the displays of modern animals are numerous fossils, like these lovely placoderms (lineage interposed between agnathans, sharks and other jawed fish).
Goblin shark head.
I seem to always forget what ray-finned fish this is (I want to say wolffish? Quick Googling suggests maybe I am right), but see it often and like its impressive bitey-ness.
Bichir and snakefish; early ray-finned fish radiations.
Armoured and similar fish today.
Armoured fish of the past; some convergent evolution within ray-fins.
Convergence- and homology- of amphibious nature in fish is another evolutionary pattern exemplified here.
Gorgeous fossils of ray-finned fish lineages that arose after the Permian extinctions, then went extinct later in the Triassic.
Note the loooooong snout on this cornetfish but the actual jaws are just at the tip.
Flying fish– those ray-fins are versatile.
Diversity of unusual ray-finned fish, including deep-water and bottom-dwelling forms.
Can you find the low-slung jaws of a dory?
Recent and fossil perch lineage fish.
It’s hard to get far into talking about evolution without bringing up the adaptive radiation of east African cichlid fish, and UMZC researchers are keen on this topic too.
Lobe-fins! Everybody dance!
Rhizodonts & kin: reasons to get out of Devonian-Carboniferous waters.
A Cretaceous fossil coelacanth (skull); not extremely different from living ones’.
Let’s admire some fossil and modern lungfish skulls, shall we? Big platey things (here, mainly looking at the palate) with lots of fusions of tiny bones on the skull roof.
Eusthenopteron fossils aren’t that uncommon but they are still great to see; and very important, because…
OK let’s stop messing around. The UMZC has one of the best displays of fossil stem-tetrapods in the world! And it should.
Another look at the pretty Acanthostega models.
Acanthostega vs. primate forelimb: so like us.
Ichthyostega parts keep Acanthostega company.
A closer look at the “Mr. Magic” Ichthyostega specimen, which takes some unpacking but is incredibly informative and was a mainstay of our 2012 model. Back of skull, left forelimb, and thorax (from left to right here).
Eucritta, another stem-tetrapod.
Closer look at Eucritta‘s skull.
Weird stem-tetrapod Crassigyrinus, which we’re still trying to figure out. It’s a fabulous specimen in terms of completeness, but messy “roadkill” with too many damn bones.
The large skull of Crassigyrinus, in right side view.
Early temnospondyl (true amphibian-line) skulls and neck.
Nectrideans or the boomerangs of the Palaeozoic.
Cool fossil frogs.
Giant Japanese salamander!
Fire salamanders: not as colourful as the real thing, but here revealing their reproductive cycle in beautiful detail.
Closeup of oviduct in above.
Sexual dimorphism in Leptodactylus frogs: the males have bulging upper arms to (I am assuming) help them hold onto females during amplexus (grasping in mating competitions).
Did I forget that Leptodactylus has big flanges on the humerus in males, to support those muscles? Seems so.
An early stem-amniote, Limnoscelis (close to mammals/reptiles divergence); cast.
Grand sea turtle skeleton.
One of my faves on display: a real pareiasaurian reptile skeleton, and you can get a good 3D look around it.
Details on above pareiasaurian.
Mammals are downstairs, but we’re reminded that they fit into tetrapod/amniote evolution nonetheless.
Let there be reptiles! And it was good.
Herps so good. (slow worm, Gila monster, glass lizard)
A curator is Dr Jason Head so you bet Titanoboa is featured!
Crocodylia: impressive specimens chosen here.
It ain’t a museum without a statuesque ratite skeleton. (There are ~no non-avian dinosaurs here– for those, go to the Sedgwick Museum across the street, which has no shortage!)
Avian diversity takes off.
Glad to see a tinamou make an appearance. They get neglected too often in museums- uncommon and often seemingly unimpressive, but I’m a fan.
I still do not understand hoatzins; the “cuckoo” gone cuckoo.
Dodo parts (and Great Auk) near the entrance.
Wow. What an oilbird taxidermy display! :-O
There we have it. Phew! That’s a lot! And I left out a lot of inverts. This upper floor is stuffed with specimens; easier there because the specimens are smaller on average than on the lower floor. Little text-heavy signage is around. I give a thumbs-up to that– let people revel in the natural glory of what their eyes show them, and give them nuggets of info to leave them wanting more so they go find out.
Now it’s in your hands– go find out yourself how lovely this museum is! I’ve just given a taste.
If you’re in London, you still have almost one week left to hurry to the Valence House in Dagenham and see a great exhibit on Ray Harryhausen’s dinosaurs and other cool “Dynamation” stop-motion models and art!
This blog post is a photo tour of what I saw, in case you cannot go.
I loooooooooooove Ray Harryhausen’s work, ever since I was a child and saw “Jason and the Argonauts” and many other films, plus “Clash of the Titans” once it came into theatres. There is the attention to detail in anatomy and locomotion, and the wondrous fantastic nature of even the more mundane creatures he animated, and the rich mythology that he drew from to inspire his creations. Modern CGI is great in a different way, but nothing I can think of in recent special effects truly beats (1) the skeleton battle in ‘Jason, and (2) the Medusa encounter in ‘Clash (to name what might be my top two faves). And so when I learned that several of the original (restored) models from those films were on exhibit in northeastern London, I requested to go there with my family for Fathers Day. Results:
Boom! Ole’ stony-gazed, snaky-haired gorgon of yore.
No deadly bow here, but the rattlesnake tail is.
Medusa concept art by Harryhausen; the “bra” was there for American censors but Ray thought it looked wrong and removed it in the final version.
Look out, Jason! Here come the Children of the Hydra! Yep, original (restored) articulated models. Joints are visible. They look ready to kick some Iolcusian butt!
Context of the exhibit- local chap befriended Harryhausen and convinced him to let him restore his models; and so here we are. On with the dinosaurs! (and other palaeo-things)
Gwangi model made in resin; non-poseable but made around time of the “Valley of Gwangi” film to help design the poseable models.
Gwangi climactic scene in church; concept art by Harryhausen.
Other ‘Gwangi characters: “Eohippus” (Hyracotherium), Ornithomimus and boy.
Cowboy lassoing an Ornithomimus as per the movie scene in ‘Gwangi? Yes please. (Harryhausen original) Jurassic Park had its T. rex lurching out of a forest to grab a Struthiomimus, intentionally mirroring the scene in ‘Gwangi where the titular Allosaurus–Tyrannosaurus hybrid chomps the Ornithomimus.
Poseable “Eohippus” original- with real fur! Great Dynamation too; very lifelike in the film.
Original Harryhausen concept art of the “Eohippus” show demo.
Suddenly, Ceratosaurus! (from “One Million Years BC”)
Styracosaurus original resin model. (from “One Million Years BC”)
Old school Polacanthus art by Alan Friswell. SPIKEY!
Old school Iguanodon art by Alan Friswell. MUSCLEY!
Panoply of archosaurs by Alan Friswell: pterodactyl, Tenontosaurus (made for the Frame Store special effects company in 2001) and tyrannosaur head (made at age 9).
Pterodactyl made at age 12, so don’t laugh.
Back to the fantastic beasts– original poseable hydra from ‘Jason!
Original Pegasus from ‘Clash! What a seamless blend of fur and feathers.
Original R2, I mean Bubo, from ‘Clash!
I forget the scene (the 1-eyed fates in ‘Clash?) but I like it. Original Harryhausen concept art.
Lunar leader from “First Men in the Moon.” (original)
Non-original (but based thereon) model by Alan Friswell, of nautiloid thingy from “Mysterious Island”.
Fiji mermaid by Alan Friswell.
“Hand of Glory” by Alan Friswell.
Pithecanthropus by Alan Friswell. Very Harryhausen in spirit.
Oddly, but somehow appropriately, there are ?350 year old whale bones on display in the hall next door, with a mysterious history.
WW2 bomb shelter in a “Victory Garden” outside the House. And the house is supposedly haunted. So take care when you visit…
What can I say? I loved it! Almost a religious experience; like seeing holy relics. Awesome in every sense of awesome.
Downside: you cannot grab the precious Dynamation models and play with them hands-on. I wanted to enact a furious Hydra-Gwangi battle. But alas, only in my imagination…
One of my favourite museums in the world, and certainly one of the best natural history museums in the UK, is Cambridge’s Museum of Zoology, AKA “University Museum of Zoology at Cambridge” (UMZC). It is now nearing a lengthy completion of renovations; the old museum exhibits and collections were excellent but needed some big changes along with the re-fabbed “David Attenborough Building” that houses them. As a longtime fan of the exhibits and user of the collection (and microCT scanner), I hurried to see the new museum once it officially opened.
And that makes a great excuse to present a photo-shoot from my visit. This focuses on the “mammal floor” below the entrance- the upper floor(s?) are still being completed and will have the birds, non-avian tetrapods, fish, etc. But the UMZC is strong in mammals and so it is natural for them to feature them in this chock-full-o-specimens display. Less talk, more images. Here we go!
All images can be clicked to mu-zoom in on them.
Stomach-Churning Rating: 3/10; bones and taxidermy and innocuous jars.
The building. The whale skeleton that hung outside for years is now cleaned up and housed right inside; you walk under it as you enter.
Entrance.
First view past the entryway: lots of cool specimens.
View from the walkway down into the ground/basement level from the entry. As specimens-per-unit-volume goes, the UMZC still scores highly and that is GOOD!
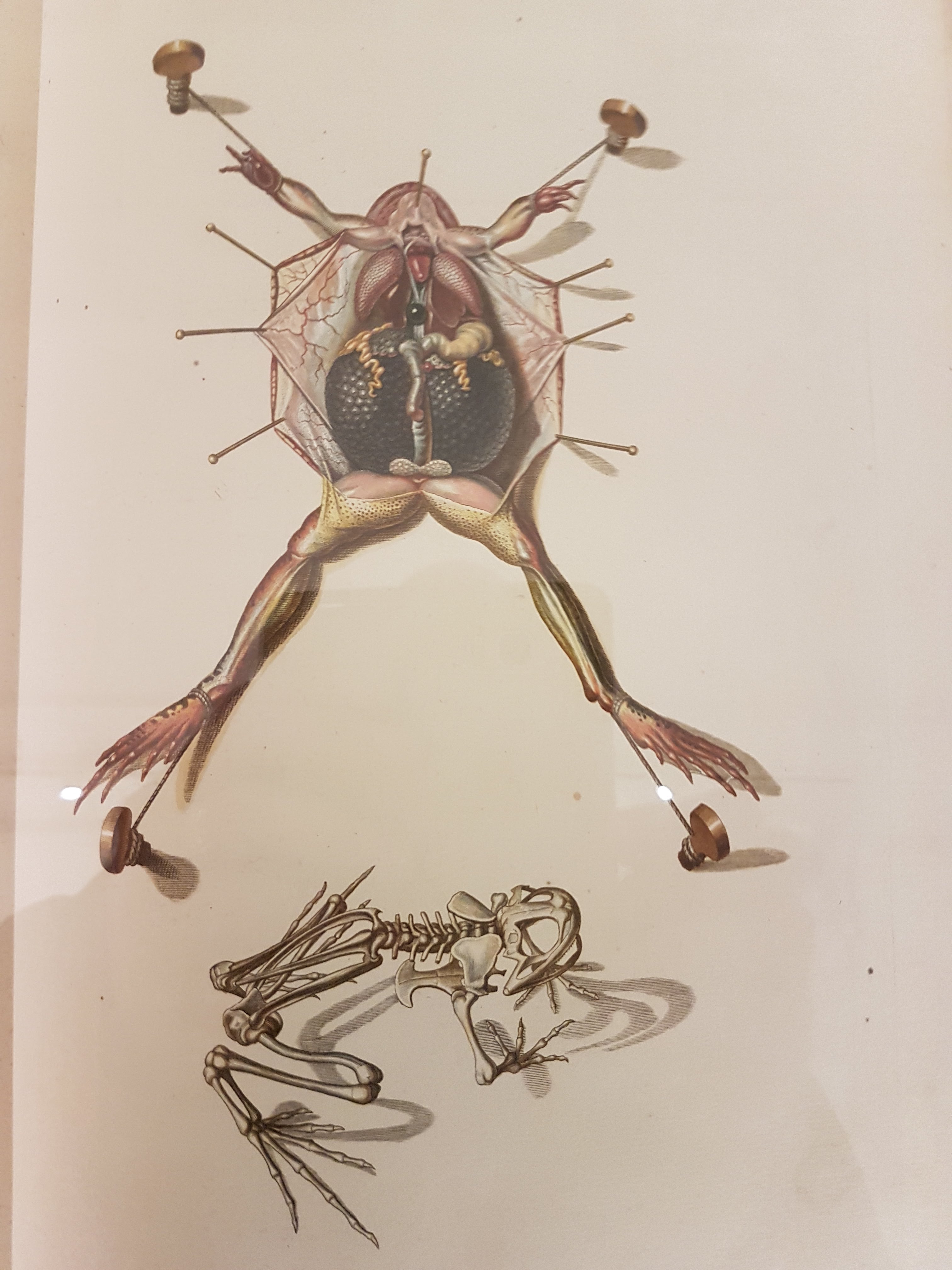
Explanation of frog dissection image below.
Gorgeous old frog dissection illustration; such care taken here.
Leeuwenhoek’s flea woodcut; I think from Arcana Naturae Detecta (1695). There is an impressive display of classic natural history books near the entryway.
Dürer/other rhino art image and info.
Darwin was famed for collecting beetles when he should have been studying theology at Cambridge as a youth, and here is some of his collection. Dang.
Darwin’s finches!
Darwin kicked off some of his meticulous work with volumes on barnacles; specimens included here; which helped fuel insights into evolution (e.g. they are “retrograde” crustaceans, not mollusks).
Darwin’s voyage: fish & other preserved specimens.
I think this is a solitaire weka (flightless island bird; see Comment below). I’ve never seen them displayed w/skeleton + taxidermy; it’s effective here.
Eryops cast. More early tetrapods will surely be featured on the upper floor; this one was on the timeline-of-life-on-Earth display.
I LOVE dioramas and this seabird nesting ground display is very evocative, especially now that I’ve visited quite a few such islands.
Mammal introduction; phylogenetic context.
Monotreme glory.
UMZC is well endowed with thylacines and this one is lovely.
“TAZ FEEL NAKED!”
Narwhal above!
Rhinocerotoidea past, present, and fading glory. 😦
Ceratotherium white rhino. The horn is not real; sadly museums (and even zoos) across the world have to worry about theft of such things, given that some people think these horns are magic.
Ceratotherium staring match. You lose.
Ceratotherium stance.
Foot of a Sumatran rhino juxtaposed with a horse’s for Perissodactyla didaction.
A tapir. As a kid, I used to wander around the house pretending to be a tapir but I did not know what noise they’d make so I’d say “tape tape tape!”.
Big Southern Elephant Seal.
Squat little fur seal.
Hippopotamus for the lot of us. (baby included)
Hippo facedown.
Skull of a dwarf Madagascar hippo.
Cave bear and sabretooth cat make an impressive Ice Age demo.
It’s a wombat.
Ain’t no don like a Diprotodon! (also note its modern miniature cousin the wombat, below)
Diprotodon facial.
Diprotodon shoulder: big clavicles bracing that joint region.
Diprotodon knee: even in big marsupials, the “parafibula”/lateral sesamoid of the knee is still generally present. And why it is there/what it does deserves much more study.
Diprotodon hip. I just find this animal’s anatomy fascinating head-to-tail.
Diprotodon front foot. Absolutely freakish.
Diprotodon hind foot. Even weirder.
Your view after having been trampled in a supine position by a Diprotodon. Not a good way to go.
Diprotodon got back.
Elephant seal’s butt continues my series of photos of big animals’ bottoms.
Asian elephant’s butt view.
African elephant butt.
Sectioned elephant skull to show pneumatic resonating chambers.
Paenungulates: hyraxes, Sirenia, elephants & kin (evolutionary demo).
AND MY HYRAX! Sorry. Had to.
Megatherium side view.
Megatherium. Yeah!
Megatherium hindlegs fascinate me. Well-heeled.
Tamandua duo.
Silky anteater; wonderful.
Armadillos.
Anteaters round out a fab display on Xenarthra.
The UMZC has everything from aardvarks to zebus. Here, conceptualized with other Afrotheria.
Golden moles: the more I read about them, the more they fascinate me.
We can all use some more solenodons in our lives!
Example of the phylogenetic context used throughout exhibits.
If you’ve got a good Okapi taxidermy, you’d better use it.
It’s a giraffe. Did you guess right?
Gerenuk showing off its bipedal capacity.
Warthogs have an inner beauty.
Pangolin. Glad to see it back on exhibit.
Nice little brown bear.
Double-barrelled shot of hyenas.
Colugo!
Nice to see some Scandentia featured.
My brain says this is a springhare (Pedetes) so I am going with what my brain says and anyway I really like this display.
When I saw this I thought, “That’s a nice… rodent thingy.” And so “rodent thing” it shall be labelled here. Enjoy the rodent thingy. Some serious taxidermy-fu in action.
Moonrats– now there’s something you seldom see a full display of. Well done!
That’s part I of this sneak peek at the evolving exhibits- I will put up a part II once the upper floor exhibits open. I highly encourage a visit!
Back in November 2016 I got an exciting email from colleague Dr. Richard Thomas, who was building a team of experts for a proposed documentary on Jumbo the elephant; the famed proboscidean of the Victorian era (and arguably most famous elephant of all time, first international celebrity animal, etc.). I knew him from social media and from our mutual interests in chicken anatomy and evolution. And that exciting email, for once, worked out! Over coming months I chatted with the film producers and they could see a place for me in the programme, contributing my expertise in elephant (postcranial) anatomy, locomotion, health/welfare etc. Lo and behold, in May 2017 I met Sir David Attenborough at Heathrow and we flew out to New York City to film with the skeleton at the American Museum of Natural History. And to cap it off, I got to meet another childhood science communication hero: Professor David Suzuki of CBC’s “The Nature of Things“– my adrenaline levels were sky high!
Brooklyn neighborhood by our hotel. Lots to do!
The show has aired in the UK and is coming very soon to Canada and the world (details below). Here’s my part of the story.
Stomach-Churning Rating: 3/10– bad bones but no blood.
We filmed from 15-19 May 2017 at the AMNH’s warehouse of mammalian skeletal remains, which is housed deep in the Brooklyn Army Terminal; a picturesque site in and of itself. And it is a site with a lot of history— WWI and II, Elvis and more.
It was a hectic week of the usual documentary stuff: repeat the same lines and motions again and again from different angles and with different paces and intonations (I cannot help in these cases but think about the Simpsons “Fallout Boy” episode), from ~9am-5pm, with plenty of downtime watching setup or other bits being filmed. I’m used to all that. But having the time to peer around the collection and chat to Richard and colleague Dr. Holly Miller (handling the tissue isotopes side of the story) about Jumbo’s skeleton was a lot of fun during downtime and filming itself. Not to mention the utter joy of studying one of the most famous museum specimens ever, and an animal widely held to be one of the largest of its kind, with much mystery surrounding its history despite its fame. (Wikipedia does a fair job of summarizing some of this)
Here are some photos to tell the story:
Photo of the team, courtesy of Infield Fly Productions (CBC production, “Jumbo: The Life of An Elephant Superstar”.
The Brooklyn Army Terminal, with a view of the harbour beyond.
Inside the terminal: old army staging area and an evocative wooden Liberty/tank artwork.
Army terminal cat. Shipping still comes through the terminal so I guess there are plenty of rats and handouts from cat-lovers to keep it going. I miss our cats when I travel so this moment was appreciated.
Whale skulls and other specimens inside the AMNH warehouse.
First view of Jumbo’s remains.
Photo opp with Sir David.
Photo opp with Prof Suzuki.
That’s the setup. I’ve done ~15 other documentary episodes/shows but this was like nothing else– simply an awesome experience.
Now the delivery: we set to studying those bones. We’d seen photos before, and Henry Fairfield Osborn had illustrated the specimen as his type of “Elephas africanus rothschildi” (Sudanese elephant; no longer valid but those were different times– it’s now just a nicely preserved Loxodonta africana africana), so we knew some of what to expect.
Looking at Osborn’s classic monograph. Oddly he didn’t address the GLARING MASSIVE PROBLEMS WITH THE TEETH!
Skull with terrible tooth pathologies– and let’s play spot Mumbo, my daughter’s toy elephant! He might even appear in some TV footage!
We had noted some serious issues with some bones (pathologies). I won’t spoil the message here but will show some images. I know some experts have voiced issues with how the tooth pathologies/growth were explained in some footage but I can’t address that here; it’s not my expertise. The important point to me is that the teeth are incredibly messed up and that can easily be linked to bad diet and other management/health issues, as the documentary explains.
Jumbo’s torso in left side view. Glorious preservation.
Right forelimb, showing that the “growth plates” (epiphyses”) were not all fused, consistent with Jumbo still growing– as expected for an African male elephant in his 20’s.
Right elbow with some pathologies consistent with degenerative joint disease.
Surprisingly, Jumbo’s feet were not in nasty condition in terms of pathologies. I’d expected to see that. They’d been painted and drilled for mounting, but were not riddled with arthritic changes that I could see.
Strange bony plaque on the left pelvis (hip) region; something I’d never seen before in any elephant (and I’ve seen many). Why? The programme offers a reasonable explanation.
Jumbo’s right hip, with bad erosion of the bone and thus presumably the overlying cartilage. Ouch!
Strange extra prong on one right rib in Jumbo- we didn’t figure that out. It could conceivably be natural variation.
So, poor Jumbo suffered some jumbo-sized problems, and in complex ways. That’s just scratching the surface of what his skeleton tells us, and there’s plenty more in the show plus plenty more we can say later– there’s real science that came out of this programme! I was surprised to find how little had been stated anywhere in the scientific literature about Jumbo’s pathologies.
Sad as Jumbo’s skeletal story is, the broader story of his life and death is sadder still. For purposes of time I don’t think any of the three versions of the show will get to delve into how Jumbo’s mother may have been slashed to death by a broadsword, as the story below describes was the ancient practice:
I’d hate to be “so pestered by a popinjay”, too.
Adding insult to injury, we can reflect on how Jumbo was taken from the Sudan to the east (across the Suez), then on boat to Italy and then overground to Paris, where he lived for a little while until the zoological garden sold him to London. Luckily Jumbo avoided becoming a meal to starving Parisians during the Prussian siege of 1870-1. So he did not become elephant consommé like some of his co-captives did. The more one learns about Jumbo’s life and the life of elephants in captivity in the 1800s, the more harrowing the tale becomes.
Jumbo is THE celebrity elephant. His name has come to mean ‘big’ and ‘bombastic’, from applications to jumbo jets to hot dogs and other (darkly ironic) forms of consumption and extravagance. He has had a jumbo effect on Western culture, but also symbolizes the complex human-elephant relationship, such as the inspiration for “Dumbo’s” own sad story. We love elephants but our fascination with them can also be their undoing, such as poaching for the ivory trade or mistreatment in captivity. Jumbo’s story writ large is also the story of elephants, and our story to learn from. If anything comes out of my participation in the Jumbo documentary for the public’s benefit, I hope it is increased empathy for how we interact with elephants. They are like us in many ways (maybe over-emphasized with anthropomorphism in many accounts), but also unlike us (maybe even unfathomable) in not only their size and anatomy but also in aspects of their prodigious intellect, emotions and social structure. Elephants aren’t just jumbo spectacles. They are jumbo responsibilities for humans now that we dominate the planet so much.
Want to catch a version of the Jumbo show? I’ll try to keep this list up to date:
BBC iplayer now: https://www.bbc.co.uk/iplayer/episode/b09jcxrj/attenborough-and-the-giant-elephant
It has been almost three months since my last post here, and things have fallen quiet on our sister blog Anatomy to You, too. I thought it was time for an update, which is mostly a summary of stuff we’ve been doing on my team, but also featuring some interesting images if you stick around. The relative silence here has partly been due to me giving myself some nice holiday time w/family in L.A., then having surgery to fix my right shoulder, then recovering from that and some complications (still underway, but the fact that I am doing this post is itself evidence of recovery).
Stomach-Churning Rating: 4/10; semi-gruesome x-rays of me and hippo bits at the end, but just bones really.
X-ray of my right shoulder from frontal view, unlabelled
Labelled x-ray
So my priorities shifted to those things and to what work priorities most badly needed my limited energy and time. I’ve also felt that, especially since my health has had its two-year rough patch, this blog has been quieter and less interactive than it used to be, but that is the nature of things and maybe part of a broader trend in blogs, too. My creative juices in terms of social media just haven’t been at their ~2011-2014 levels but much is out of my control, and I am hopeful that time will reverse that trend. Enough about all this. I want to talk about science for the rest of this post.
My team, and collaborators as well, have published six recent studies that are very relevant to this blog’s theme- how about we run through them quickly? OK then.
Panagiotopoulou, O., Pataky, T.C., Day, M., Hensman, M.C., Hensman, S., Hutchinson, J.R., Clemente, C.J. 2016. Foot pressure distributions during walking in African elephants (Loxodonta africana). Royal Society Open Science 3: 160203.
Our Australian collaborators got five African elephants together in Limpopo, South Africa and walked them over pressure-measuring mats, mimicking our 2012 study of Asian elephants. While sample sizes were too limited to say much statistically, in qualitatively descriptive terms we didn’t find striking differences between the two species’ foot pressure patterns. I particularly like how the centre of pressure of each foot (i.e. abstracting all regional pressures down to one mean point over time) followed essentially the same pattern in our African and Asian elephants, with a variable heelstrike concentration that then moved forward throughout the step, and finally moved toward the outer (3rd-5th; especially 3rd) toes as the foot pushed off the ground, as below.
African elephant foot COP traces vs. time in red; Asian elephant in orange-yellow. Left and right forefeet above; hindfeet below.
Gradually, this work is moving the field toward better ability to use similar techniques to compare elephant foot mechanics among species, individuals, or over time– especially with the potential of using this method (popular in human clinical gait labs) to monitor foot (and broader musculoskeletal) health in elephants. I am hopeful that a difference can be made, and the basic science we’ve done to date will be a foundation for that.
Panagiotopoulou, O., Rankin, J.W., Gatesy, S.M., Hutchinson, J.R. 2016. A preliminary case study of the effect of shoe-wearing on the biomechanics of a horse’s foot. PeerJ 4: e2164.
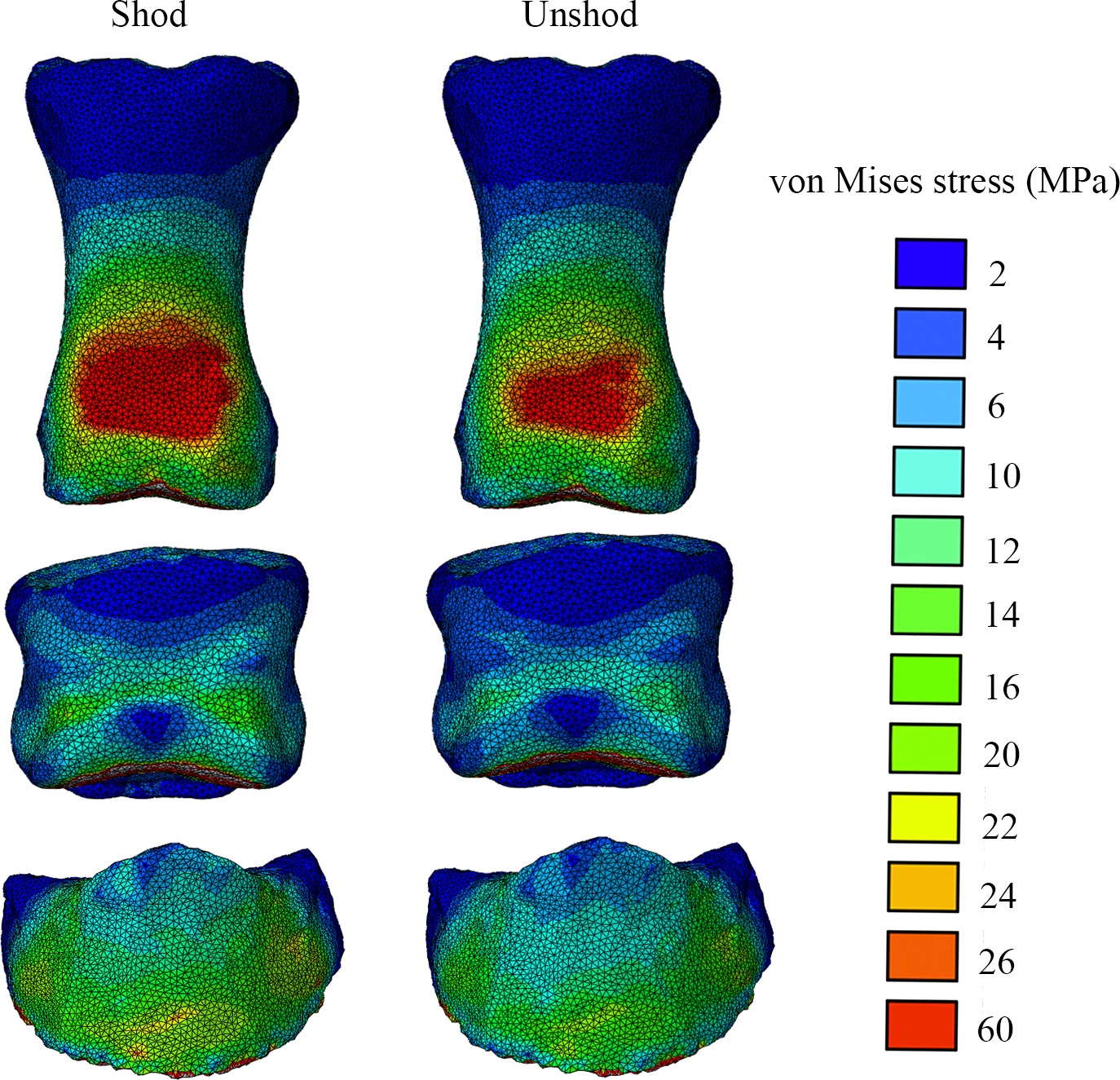
Finally, about six years after we collected some very challenging experimental data in our lab, we’ve published our first study on them. It’s a methodological study of one horse, not something one can hang any hats on statistically, but we threw the “kitchen sink” of biomechanics at that horse (harmlessly!) by combining standard in vivo forceplate analysis with “XROMM” (scientific rotoscopy with biplanar fluoroscopy or “x-ray video”) to conduct dynamic analysis of forefoot joint motions and forces (with and without horseshoes on the horse), and then to use these data as input values for finite element analysis (FEA) of estimated skeletal stresses and strains. This method sets the stage for some even more ambitious comparative studies that we’re finishing up now. And it is not in short supply of cool biomechanical, anatomical images so here ya go:
Above: The toe bones (phalanges) of our horse’s forefoot in dorsal (cranial/front) view, from our FEA results, with hot colours showing higher relative stresses- in this case, hinting (but not demonstrating statistically) that wearing horseshoes might increase stresses in some regions on the feet. But more convincingly, showing that we have a scientific workflow set up to do these kinds of biomechanical calculations from experiments to computer models and simulations, which was not trivial.
And a cool XROMM video of our horse’s foot motions:
Bates, K.T., Mannion, P.D., Falkingham, P.L., Brusatte, S.L., Hutchinson, J.R., Otero, A., Sellers, W.I., Sullivan, C., Stevens, K.A., Allen, V. 2016. Temporal and phylogenetic evolution of the sauropod dinosaur body plan. Royal Society Open Science 3: 150636.
I had the good fortune of joining a big international team of sauropod experts to look at how the shapes and sizes of body segments in sauropods evolved and how those influenced the position of the body’s centre of mass, similar to what we did earlier with theropod dinosaurs. My role was minor but I enjoyed the study (despite a rough ride with some early reviews) and the final product is one cool paper in my opinion. Here’s an example:
The (embiggenable-by-clicking) plot shows that early dinosaurs shifted their centre of mass (COM) backwards (maybe related to becoming bipedal?) and then sauropods shifted the COM forwards again (i.e. toward their forelimbs and heads) throughout much of their evolution. This was related to quadrupedalism and giant size as well as to evolving a longer neck; which makes sense (and I’m glad the data broadly supported it). But it is also a reminder that not all sauropods moved in the same ways- the change of COM would have required changes in how they moved. There was also plenty of methodological nuance here to cover all the uncertainties but for that, see the 17 page paper and 86 pages of supplementary material…
Randau, M., Goswami, A., Hutchinson, J.R., Cuff, A.R., Pierce, S.E. 2016. Cryptic complexity in felid vertebral evolution: shape differentiation and allometry of the axial skeleton. Zoological Journal of the Linnean Society 178:183-202.
Back in 2011, Stephanie Pierce, Jenny Clack and I tried some simple linear morphometrics (shape analysis) to see how pinniped (seal, walrus, etc) mammals changed their vertebral morphology with size and regionally across their backbones. Now in this new study, with “Team Cat” assembled, PhD student Marcela Randau collected her own big dataset for felid (cat) backbones and applied some even fancier techniques to see how cat spines change their shape and size. We found that overall the vertebrae tended to get relatively more robust in larger cats, helping to resist gravity and other forces, and that cats with different ecologies across the arboreal-to-terrestrial spectrum also changed their (lumbar) vertebral shape differently. Now Marcela’s work is diving even deeper into these issues; stay tuned…
Example measurements taken on felid vertebrae, from the neck (A-F) to the lumbar region (G-J), using a cheetah skeleton.
Charles, J.P., Cappellari, O., Spence, A.J., Hutchinson, J.R., Wells, D.J. 2016. Musculoskeletal geometry, muscle architecture and functional specialisations of the mouse hindlimb. PLOS One 11(4): e0147669.
RVC PhD student James Charles measured the heck out of some normal mice, dissecting their hindlimb muscle anatomy, and using microCT scans produced some gorgeous images of that anatomy too. In the process, he also quantified how each muscle is differently specialized for the ability to produce large forces, rapid contractions or fine control. Those data were essential for the next study, where we got more computational!
Charles, J.P., Cappellari, O., Spence, A.J., Wells, D.J., Hutchinson, J.R. 2016. Muscle moment arms and sensitivity analysis of a mouse hindlimb musculoskeletal model. Journal of Anatomy 229:514–535.
James wrangled together a lovely musculoskeletal model of our representative mouse subject’s hindlimb in the SIMM software that my team uses for these kinds of biomechanical analyses. As we normally do as a first step, we used the model to estimate things that are hard to measure directly, such as the leverages (moment arms) of each individual muscle and how those change with limb posture (which can produce variable gearing of muscles around joints). James has his PhD viva (defense) next week so good luck James!
The horse and mouse papers are exemplars of what my team now does routinely. For about 15 years now, I’ve been building my team toward doing these kinds of fusion of data from anatomy, experimental biomechanics, musculoskeletal and other models, and simulation (i.e. estimating unmeasurable parameters by telling a model to execute a behaviour with a given set of criteria to try to perform well). Big thanks go to collaborator Jeff Rankin for helping us move that along lately. Our ostrich study from earlier this year shows the best example we’ve done yet with this, but there’s plenty more to come.
I am incredibly excited that, now that my team has the tools and expertise built up to do what I’ve long wanted to do, we can finally deliver the goods on the aspirations I had back when I was a postdoc, and which we have put enormous effort into pushing forward since then. In addition to new analyses of horses and mice and other animals, we’ll be trying to push the envelope more with how well we can apply similar methods to extinct animals, which brings new challenges– and evolutionary questions that get me very, very fired up.
Here we are, then; time has brought some changes to my life and work and it will continue to as we pass this juncture. I suspect I’ll look back on 2016 and see it as transformative, but it hasn’t been an easy year either, to say the least. “Draining” is the word that leaps to mind right now—but also “Focused” applies, because I had to try to be that, and sometimes succeeded. I’ve certainly benefited a lot at work from having some talented staff, students and other collaborators cranking out cool papers with me.
I still have time to do other things, too. Once in a while, a cool critter manifests in The Freezers. Check out a hippo foot from a CT scan! It’s not my best scan ever (noisy data) but it shows the anatomy fairly well, and some odd pathologies such as tiny floating lumps of mineralized soft tissue here and there. Lots to puzzle over.