One of my favourite museums in the world, and certainly one of the best natural history museums in the UK, is Cambridge’s Museum of Zoology, AKA “University Museum of Zoology at Cambridge” (UMZC). It is now nearing a lengthy completion of renovations; the old museum exhibits and collections were excellent but needed some big changes along with the re-fabbed “David Attenborough Building” that houses them. As a longtime fan of the exhibits and user of the collection (and microCT scanner), I hurried to see the new museum once it officially opened.
And that makes a great excuse to present a photo-shoot from my visit. This focuses on the “mammal floor” below the entrance- the upper floor(s?) are still being completed and will have the birds, non-avian tetrapods, fish, etc. But the UMZC is strong in mammals and so it is natural for them to feature them in this chock-full-o-specimens display. Less talk, more images. Here we go!
All images can be clicked to mu-zoom in on them.
Stomach-Churning Rating: 3/10; bones and taxidermy and innocuous jars.

The building. The whale skeleton that hung outside for years is now cleaned up and housed right inside; you walk under it as you enter.

Entrance.

First view past the entryway: lots of cool specimens.

View from the walkway down into the ground/basement level from the entry. As specimens-per-unit-volume goes, the UMZC still scores highly and that is GOOD!

Explanation of frog dissection image below.
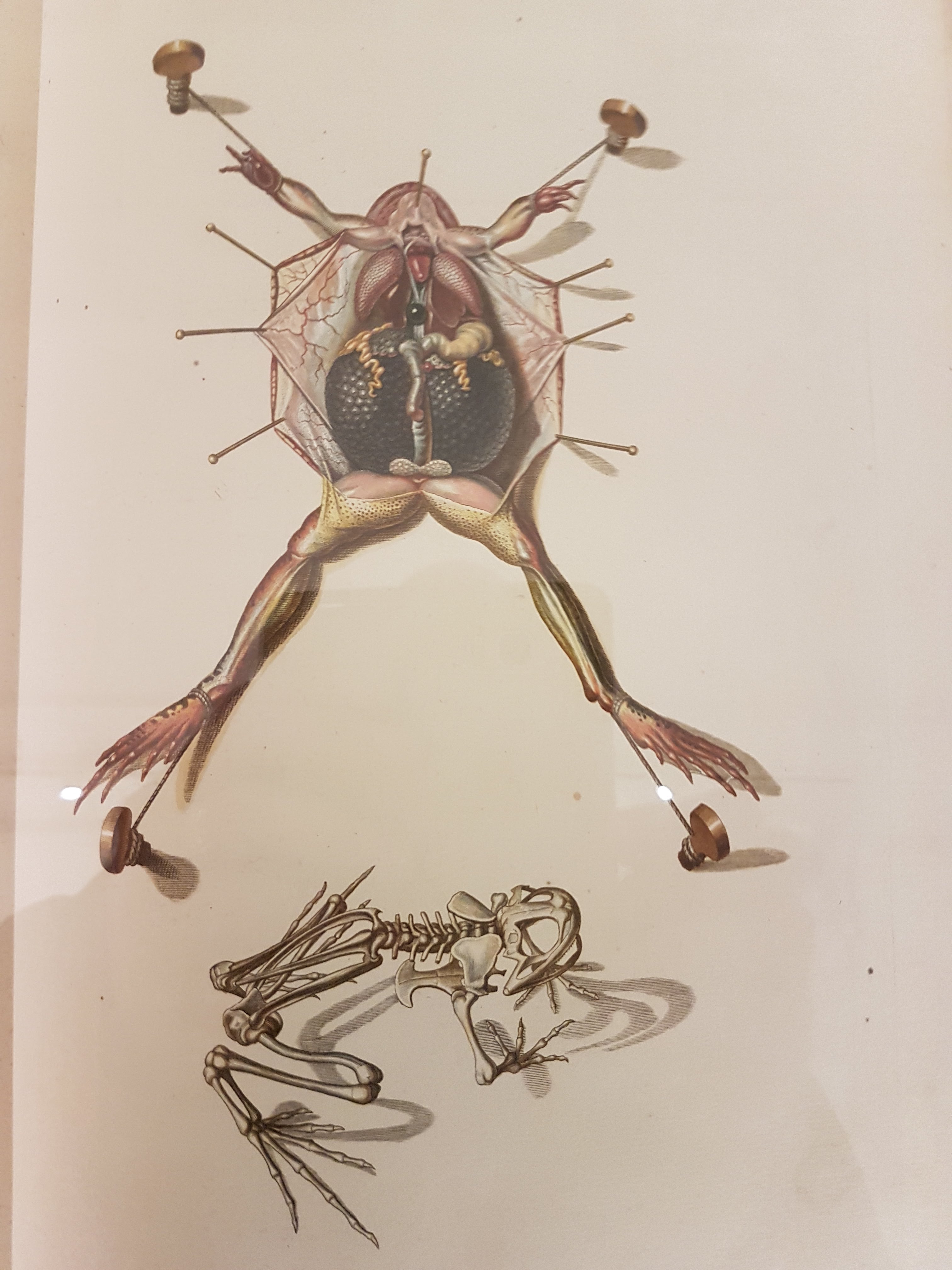
Gorgeous old frog dissection illustration; such care taken here.
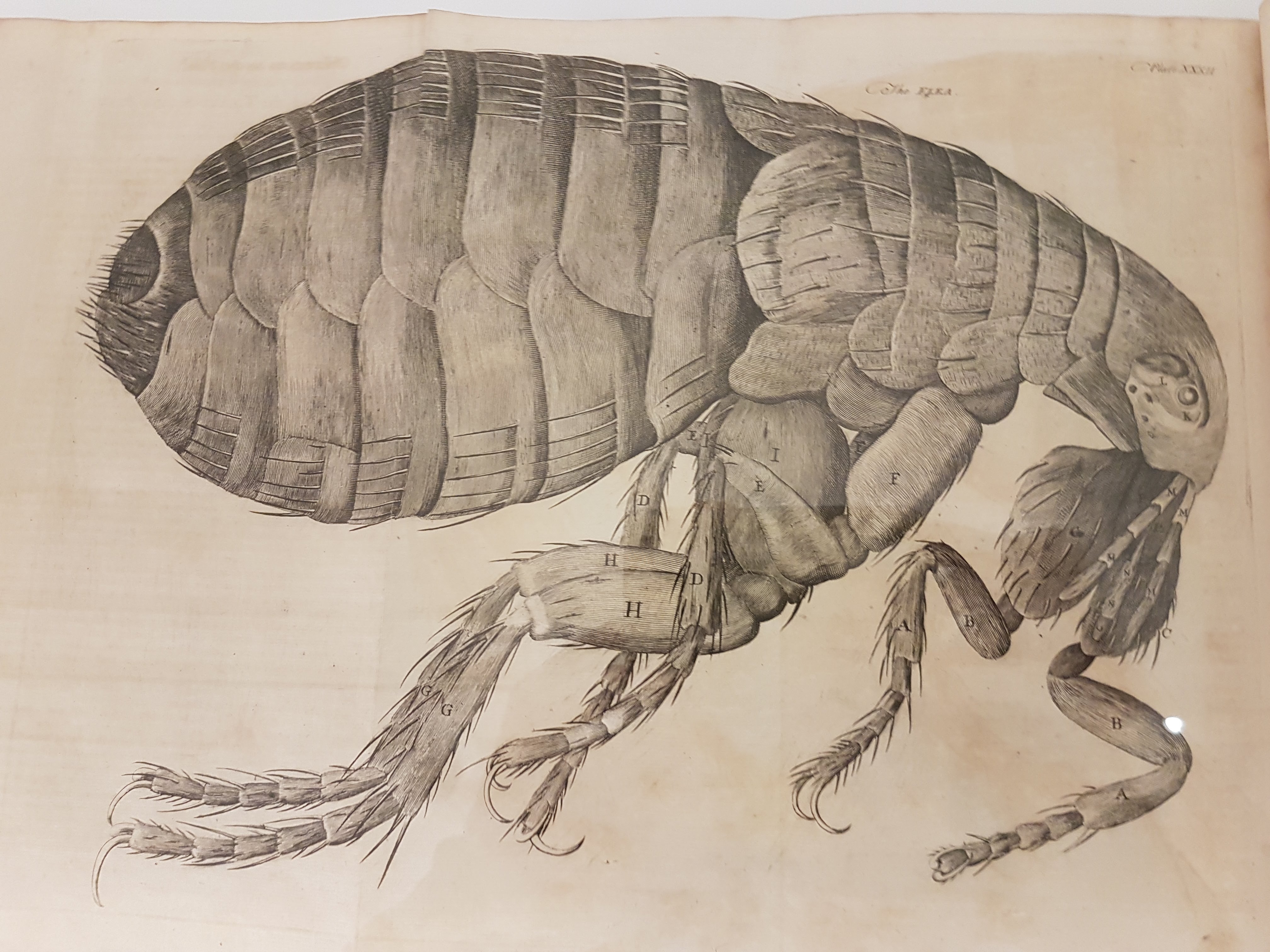
Leeuwenhoek’s flea woodcut; I think from Arcana Naturae Detecta (1695). There is an impressive display of classic natural history books near the entryway.

Dürer/other rhino art image and info.

Darwin was famed for collecting beetles when he should have been studying theology at Cambridge as a youth, and here is some of his collection. Dang.

Darwin’s finches!

Darwin kicked off some of his meticulous work with volumes on barnacles; specimens included here; which helped fuel insights into evolution (e.g. they are “retrograde” crustaceans, not mollusks).

Darwin’s voyage: fish & other preserved specimens.

I think this is a solitaire weka (flightless island bird; see Comment below). I’ve never seen them displayed w/skeleton + taxidermy; it’s effective here.

Eryops cast. More early tetrapods will surely be featured on the upper floor; this one was on the timeline-of-life-on-Earth display.

I LOVE dioramas and this seabird nesting ground display is very evocative, especially now that I’ve visited quite a few such islands.

Mammal introduction; phylogenetic context.

Monotreme glory.

UMZC is well endowed with thylacines and this one is lovely.

“TAZ FEEL NAKED!”

Narwhal above!

Rhinocerotoidea past, present, and fading glory. 😦

Ceratotherium white rhino. The horn is not real; sadly museums (and even zoos) across the world have to worry about theft of such things, given that some people think these horns are magic.

Ceratotherium staring match. You lose.

Ceratotherium stance.

Foot of a Sumatran rhino juxtaposed with a horse’s for Perissodactyla didaction.

A tapir. As a kid, I used to wander around the house pretending to be a tapir but I did not know what noise they’d make so I’d say “tape tape tape!”.

Big Southern Elephant Seal.

Squat little fur seal.

Hippopotamus for the lot of us. (baby included)

Hippo facedown.

Skull of a dwarf Madagascar hippo.

Cave bear and sabretooth cat make an impressive Ice Age demo.

It’s a wombat.

Ain’t no don like a Diprotodon! (also note its modern miniature cousin the wombat, below)

Diprotodon facial.

Diprotodon shoulder: big clavicles bracing that joint region.

Diprotodon knee: even in big marsupials, the “parafibula”/lateral sesamoid of the knee is still generally present. And why it is there/what it does deserves much more study.

Diprotodon hip. I just find this animal’s anatomy fascinating head-to-tail.
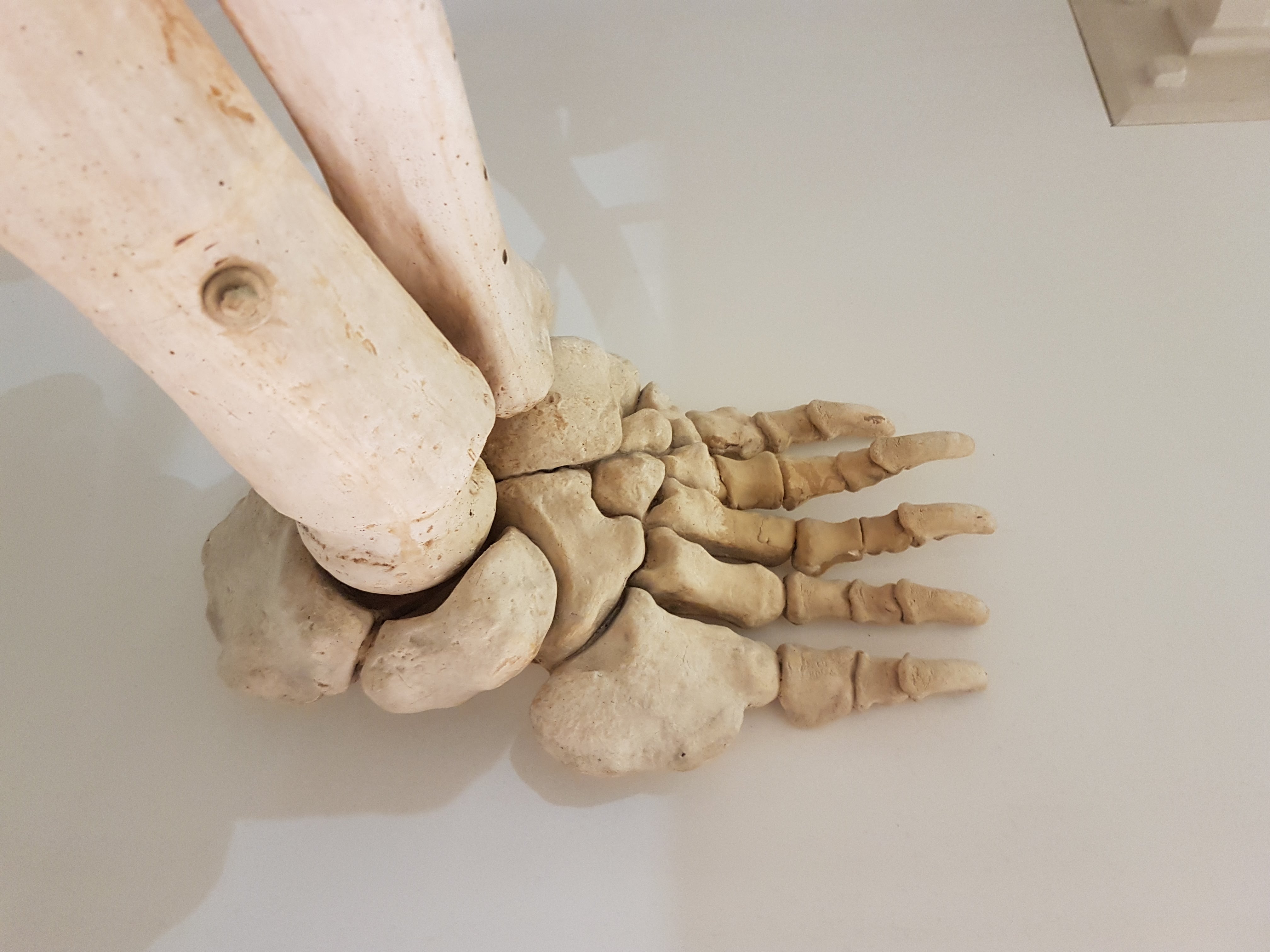
Diprotodon front foot. Absolutely freakish.

Diprotodon hind foot. Even weirder.

Your view after having been trampled in a supine position by a Diprotodon. Not a good way to go.

Diprotodon got back.

Elephant seal’s butt continues my series of photos of big animals’ bottoms.

Asian elephant’s butt view.

African elephant butt.

Sectioned elephant skull to show pneumatic resonating chambers.

Paenungulates: hyraxes, Sirenia, elephants & kin (evolutionary demo).

AND MY HYRAX!
Sorry. Had to.

Megatherium side view.

Megatherium. Yeah!

Megatherium hindlegs fascinate me. Well-heeled.

Tamandua duo.

Silky anteater; wonderful.

Armadillos.

Anteaters round out a fab display on Xenarthra.

The UMZC has everything from aardvarks to zebus. Here, conceptualized with other Afrotheria.

Golden moles: the more I read about them, the more they fascinate me.

We can all use some more solenodons in our lives!

Example of the phylogenetic context used throughout exhibits.

If you’ve got a good Okapi taxidermy, you’d better use it.

It’s a giraffe. Did you guess right?

Gerenuk showing off its bipedal capacity.

Warthogs have an inner beauty.

Pangolin. Glad to see it back on exhibit.

Nice little brown bear.

Double-barrelled shot of hyenas.

Colugo!

Nice to see some Scandentia featured.

My brain says this is a springhare (Pedetes) so I am going with what my brain says and anyway I really like this display.

When I saw this I thought, “That’s a nice… rodent thingy.” And so “rodent thing” it shall be labelled here. Enjoy the rodent thingy. Some serious taxidermy-fu in action.

Moonrats– now there’s something you seldom see a full display of. Well done!
That’s part I of this sneak peek at the evolving exhibits- I will put up a part II once the upper floor exhibits open. I highly encourage a visit!
For Mike: 





































































")














