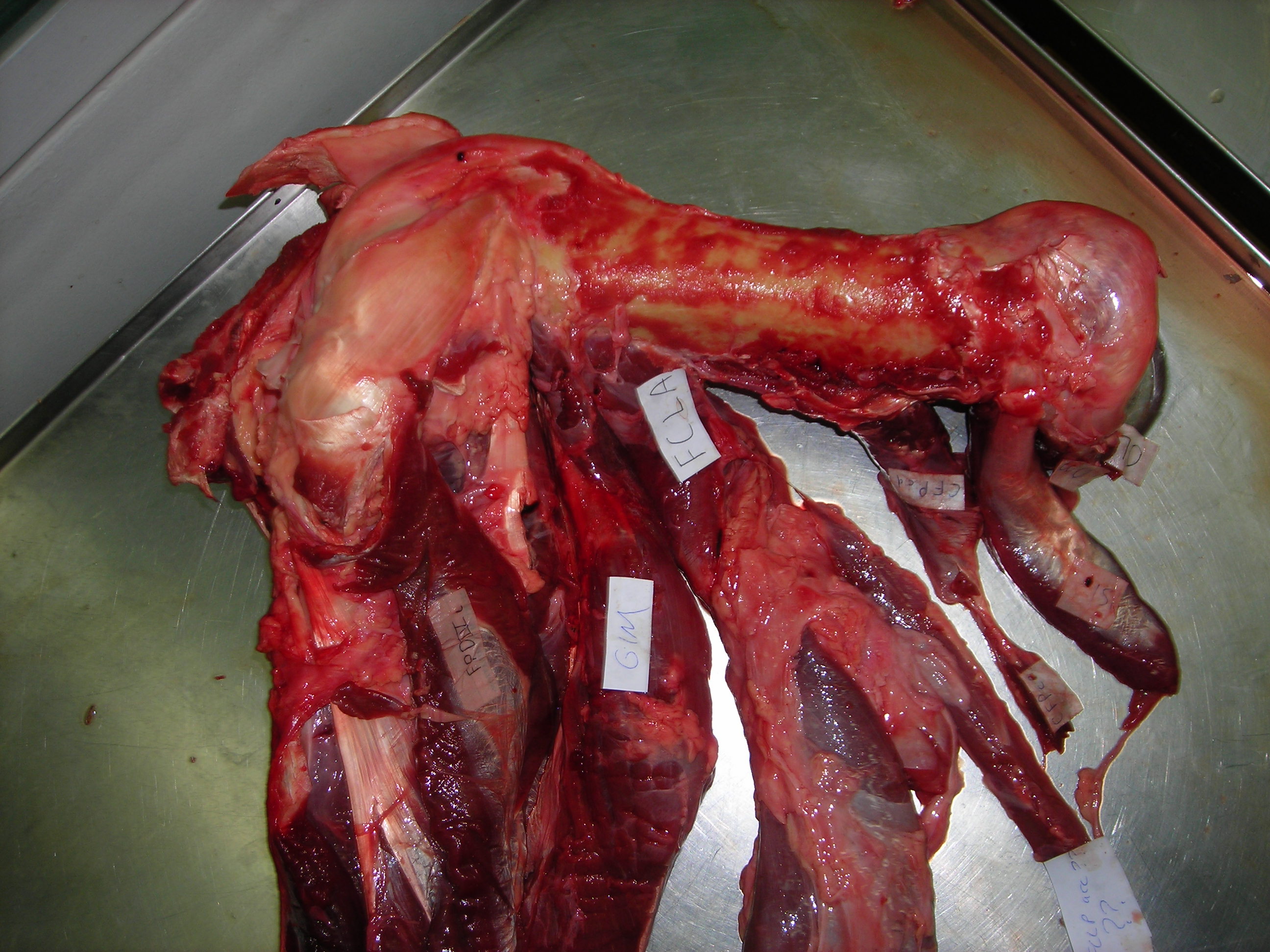
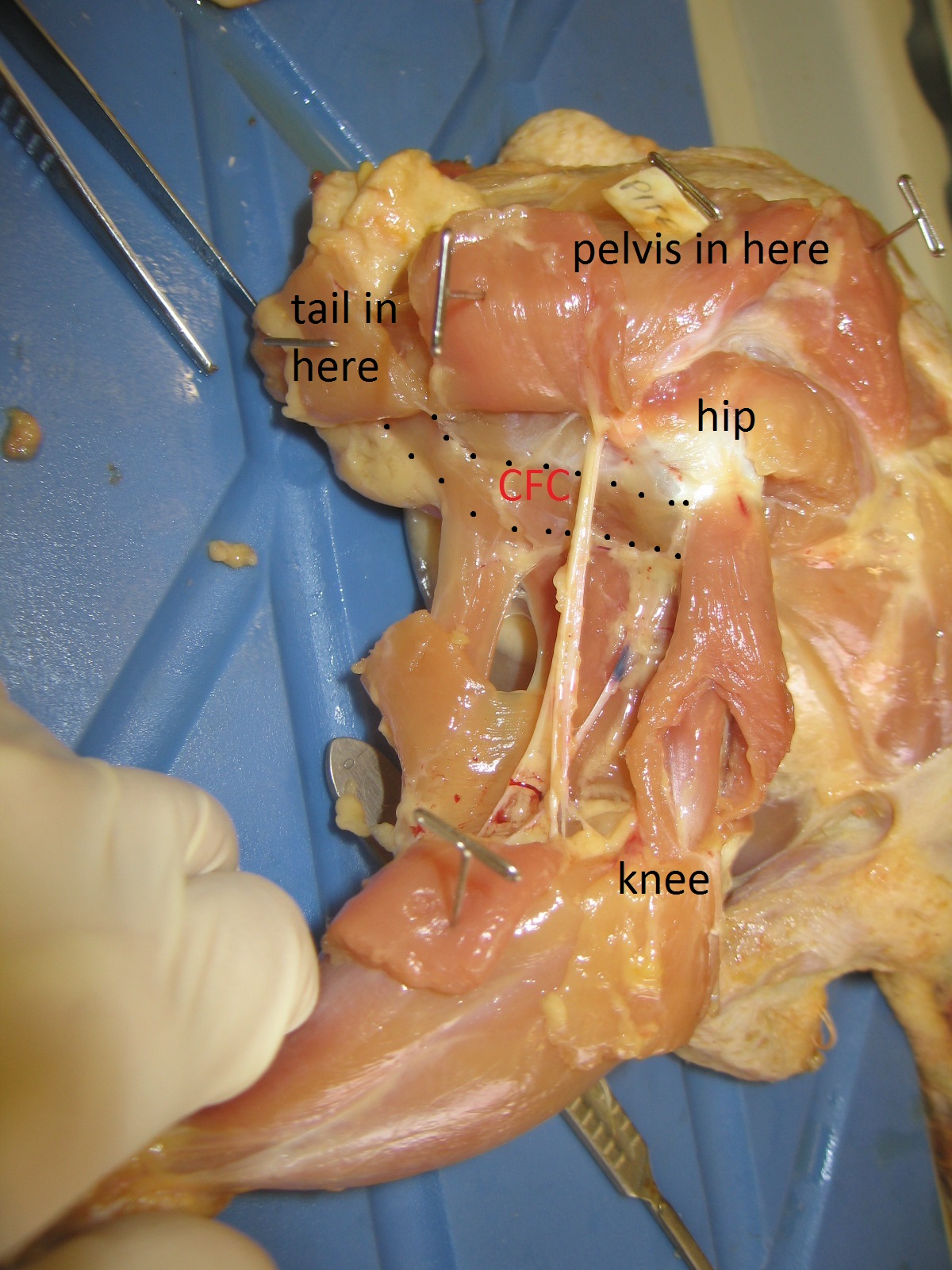
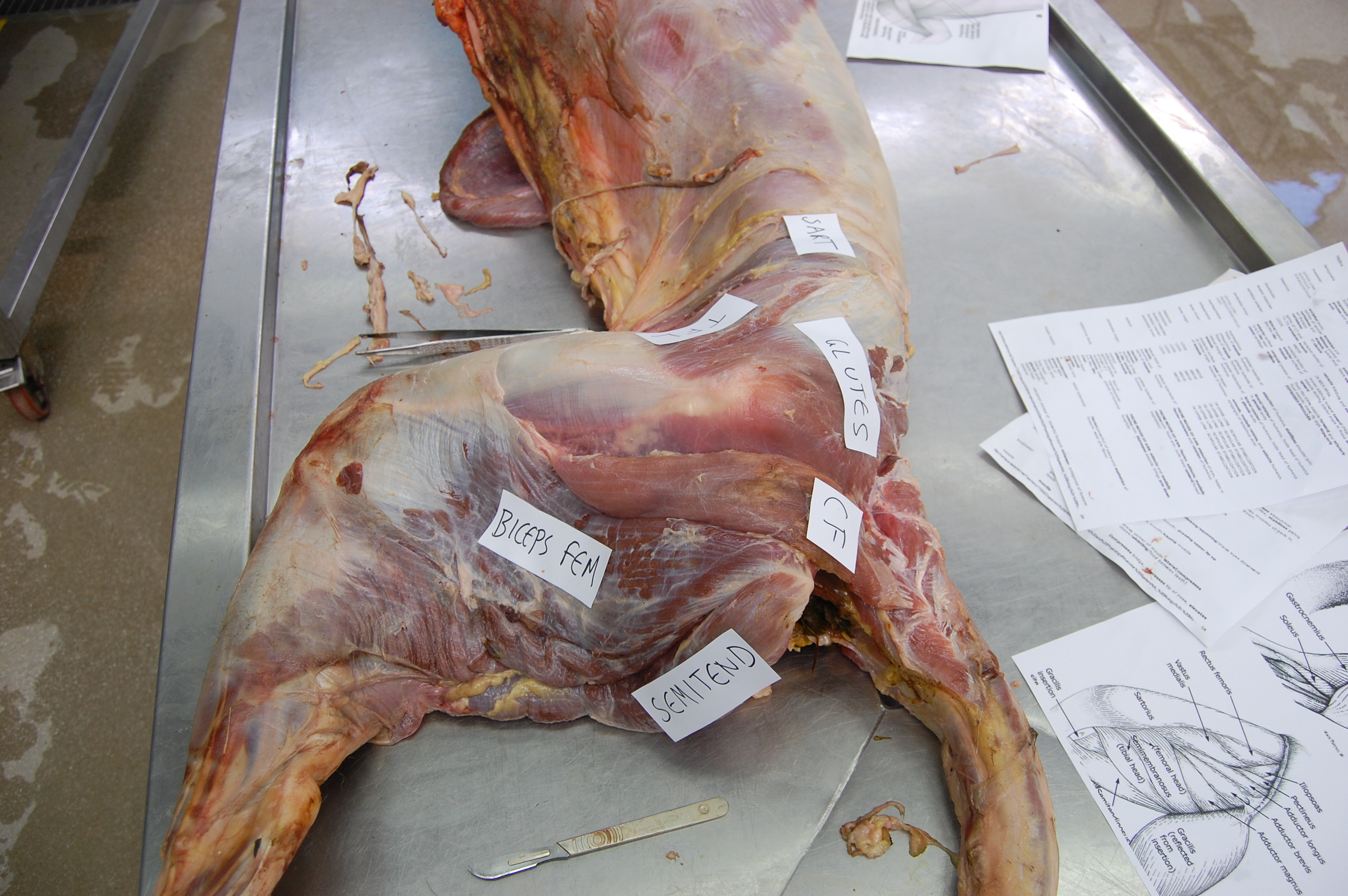
The blog is back! Briefly. With dinosaurs. Back in 2005, I published a paper in which I used a “SIMM” 3D musculoskeletal biomechanical model of Tyrannosaurus rex to analyse its muscle actions and infer a relatively upright hindlimb pose. This was an outcome from my NSF-funded postdoctoral research at Stanford University, in which engineers kindly taught me how to use SIMM (handing me a loaded gun?). Part of my plan all along was to build multiple such models along a rough evolutionary sequence to revisit old questions I had with past, qualitative functional morphology papers from 2000 onwards, and see if biomechanics could quantitatively reveal more about the functional evolution of dinosaur hindlimb muscles. So I got data for modelling some extinct dinosaurs (theropods Dilophosaurus, Allosaurus, Velociraptor) and living birds (Struthio, others) and published nuggets of that but held others back…
Stomach-Churning Rating: -1/10; dinosaurs!
I handed these 3D model data off to my PhD student Vivian Allen in ~2007, charging him with the task of making more models to flesh out the phylogeny and finish what I’d started. And he sure did. He graduated, did a couple of postdocs with me, and we gradually massaged his thesis chapter on this topic into a draft paper. Easier said than done, though! That’s why 14 more years have passed.
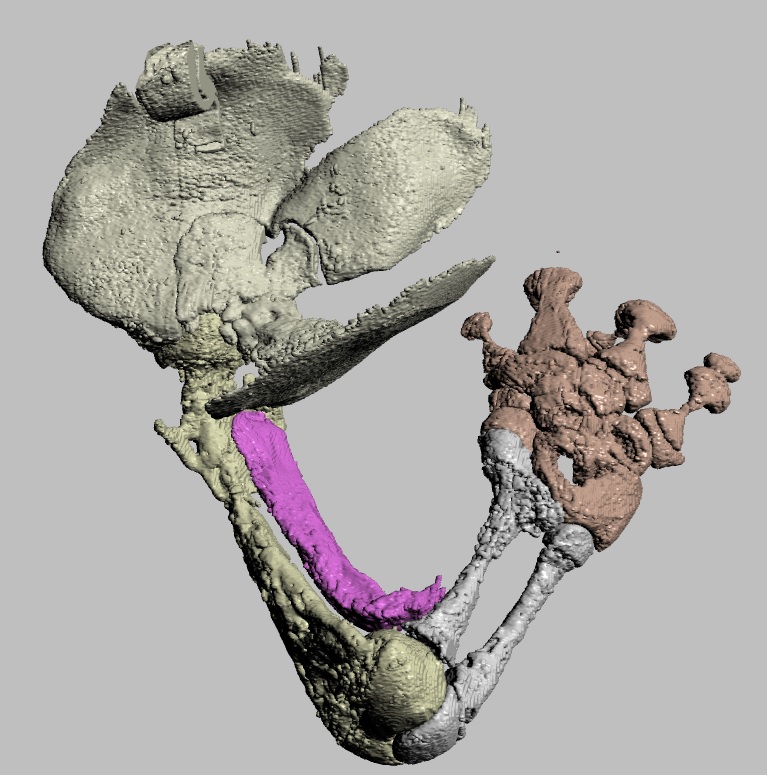
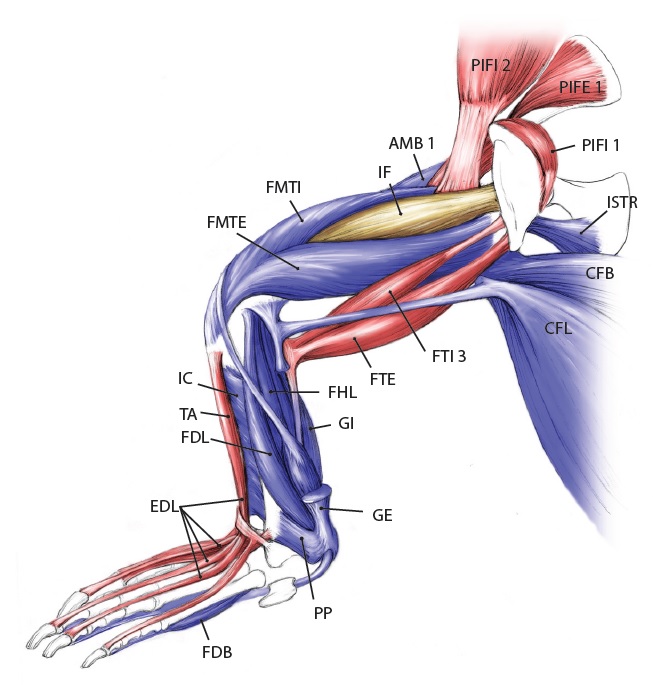
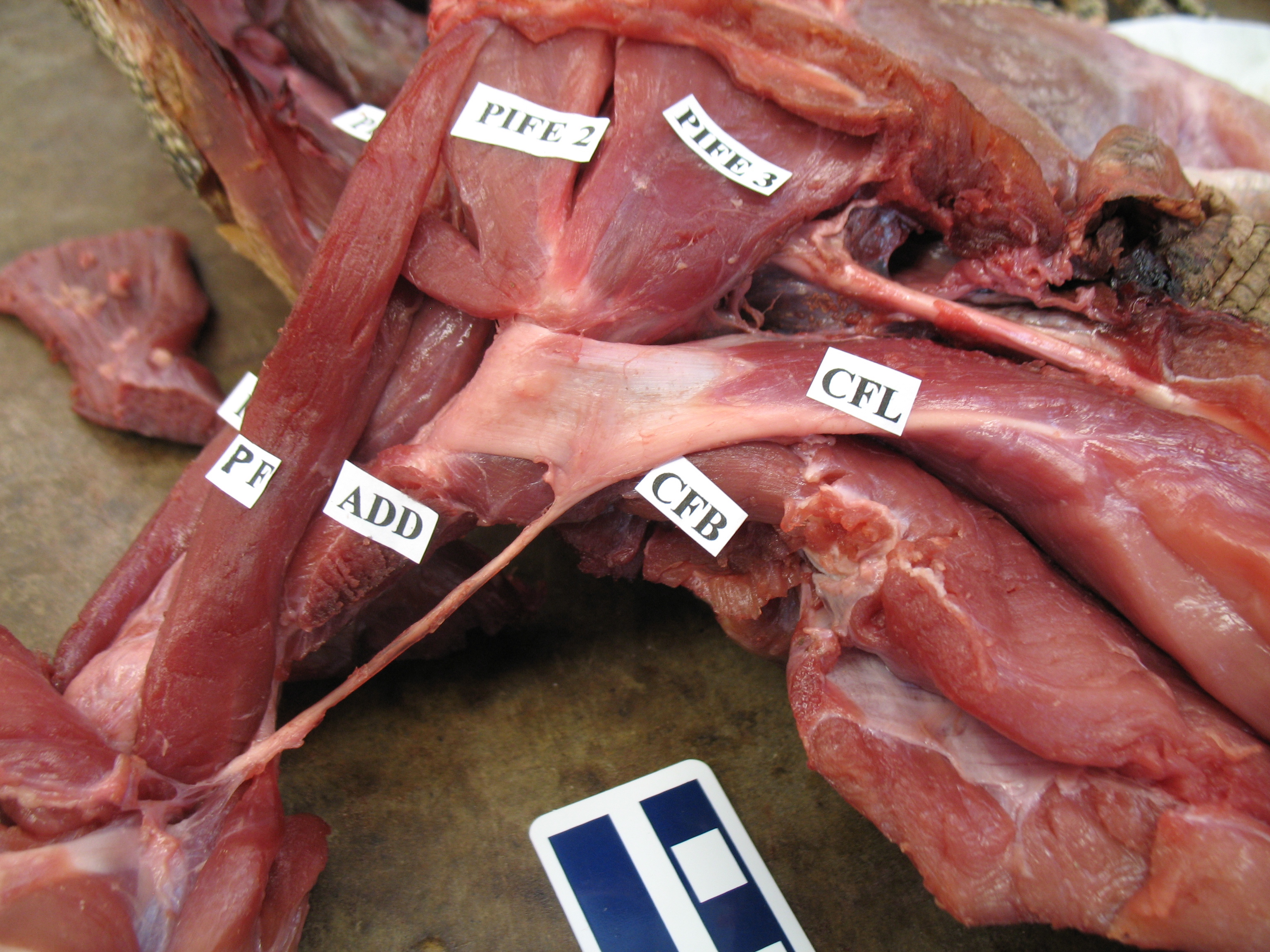
Viv came up with some clever tools in MATLAB software code (from which he became a very competent programmer and went on to a successful career in that!) to boil complex data on muscle leverages (moment arms) across a wide range of joint motion for the hindlimbs for each taxon.
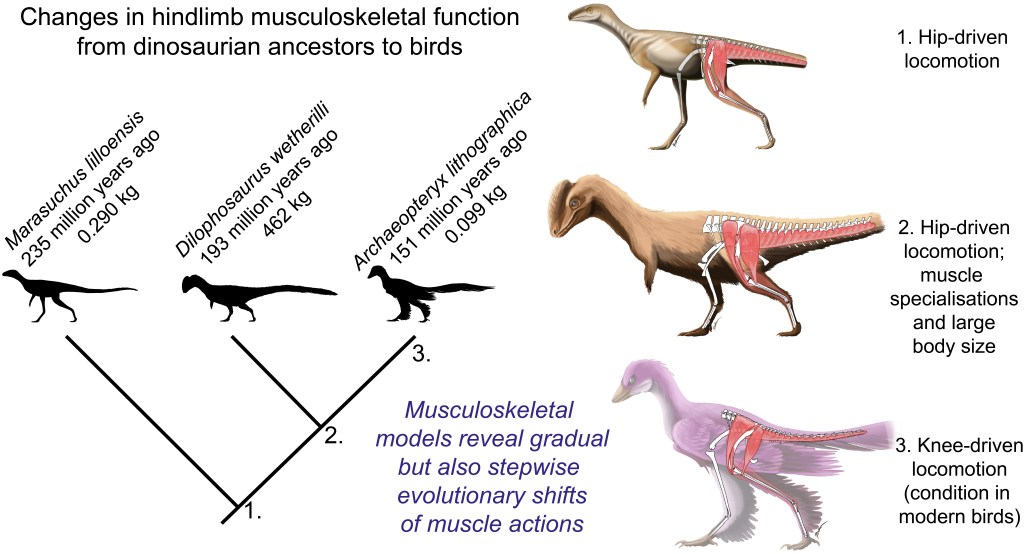
These data then were fed into further code that took the results from all models, ultimately 13 of them from an Australian freshwater crocodile to two living birds and 10 extinct dinosaurs plus close cousin Mara/Lagosuchus (Figure 1). The code expressed these leverages as changes in ancestral values along the main branch of the evolutionary tree from early (Triassic) “ruling reptile” Archosauria (represented here just by the croc as a proxy) to modern birds, and 9 main ancestral “nodes” in between. Our code tracked both how each of 35 hindlimb muscles we modelled evolved in its leverage, as well as overall “average” leverage of functional groups around the hip, knee and ankle joints.
So, back and forth we went for some 10 years playing with the models (see Video below), data and code, and the paper describing the whole thing, slowly closing in on a final version but also sometimes distracted by our other projects and Real Life Stuff like health and children, and concerns about how we conducted this study (i.e. a lot of fiddling).

Then I got a new grant “DAWNDINOS” that changed the scene for me, refocusing my team’s energies onto the Triassic (and early Jurassic) and the evolutionary biomechanics of diverse archosaurs’ locomotion, assessed with both LOTS of experimental studies of living crocs and birds, and LOTS of predictive simulations of locomotion. Stay tuned for much more on that from our team, but we’ve already published some key steps here. Most notably, we developed an improved protocol for modelling and simulating our animals, as shown by Bishop et al.’s 2021 study of the early theropod Coelophysis bauri (also appearing in the current paper). Awkwardly for me, that new method rendered our old models and methods a bit obsolete (although still fine), so I pushed to publish this current paper with Viv, and brought collaborator Dr. Brandon Kilbourne on board to aid in some final stats, figures and more. That finally did it, and now we’ve published the paper in Science Advances. Deep breath.
Well what’s the paper about, then? We used our 13 models and processed evolutionary functional patterns to test three main questions (hypotheses) about muscle leverage, making educated guesses at what might prevail from early Archosauria to Aves:
- Hip extensor / flexor (i.e. femur retractor/protractor) moment arm ratios remained constant. We weren’t sure what to expect, as these antagonists both seem to change a lot on the whole lineage, so we went with this prediction.
- Knee flexor / extensor ratios decreased; i.e. the flexors (“hamstrings” etc.) weakened and/or extensors (equivalent of our quadriceps) strengthened their leverage. Anatomy of the knee joint and muscles around it suggests this, plus since Gatesy’s 1990-onwards studies we’ve expected archosaurs to shift from more ‘hip-based’ to more ‘knee-based’ locomotion as we get closer to avian ancestry.
- Hip medial (internal) long-axis rotator / abductor (i.e. pronators of the limb vs. those that draw the leg away from the body) ratios increased. This idea comes right from my paper w/Gatesy in 2000, where we surmised that archosaurs shifted from relying on hip adductors (in crocs/other quadrupeds) to abductors (in bipedal dinosaurs; like humans) to medial rotators (‘torsional control’ as in birds today) during weight support.
Moreover, we reconstructed the evolution of 35 muscles’ actions across ~250 million years, which was a new step.
Here’s a summary of what we found (Figure 2):
So, overall hypothesis 1 about hip extensors/flexors ended up complicated; rejected because hip flexor leverage actually increased. Furthermore, we found that around the ancestral nodes for early theropod dinosaurs (Neotheropda through Avetheropoda; around 200 Mya), there were peaks in muscle leverage (size-normalized) that surprised us, and persisted despite many different analyses we threw at them over the years. As far as we could tell, these peaks that kept appearing for various muscles’ actions were “real” (estimates). Which meant these ancestors may have had specialised high leverage relative to both their own ancestors and descendants; the peaks got reversed in evolution. These ancestors had some other weird anatomical and functional traits, such as tightly articulated hip joints early on (which they lost later), increased body size in the later forms, more ‘macropredatory’ ecology (e.g. eating sauropods?), and a centre of mass of the body that was shifted forwards (due to big arms and heads/necks). This weirdness is a cool unexpected finding that showed up for the other hypotheses too, and it needs some more investigating. A ‘failed’ hypothesis test led to neat insights.

Hypotheses 2 and 3 found good support, on the contrary, overall (Figure 3). We seem to have been able to quantify the shifts from hip-based to knee-based, and abductor-based to medial-rotator-based, muscle actions. I find that very satisfying. Ankle weight support (extension) capacity also increased, which fits morphological changes fairly well. If you’re into archosaur limb muscle form and function, there’s a lot more food for thought in the paper.
Funnily enough, ~20 years has been sufficient time that we could have had plenty more models in this study if we’d delayed it even longer and re-re-re-analysed our data. But we had to draw the line somewhere and not infinitely revise with every new model we’ve been creating. With the current state of musculoskeletal modelling in my group, we could have more than doubled our sample size and fleshed out the most important gaps such as in the crocodile-lineage (extinct Pseudosuchia) and other Triassic forms plus elsewhere. A big challenge remains having some nice 3D-preserved early fossil birds beyond Archaeopteryx; e.g. so many nice Chinese ones are too flat (e.g. joints we need) to reliably model here. It’s something that can still be done and is worth doing, but I suspect the general trends we’ve found along the dinosaur lineage are “correct”.
What’s personally important to me about this paper is (1) how it not only bridges a huge morphofunctional gap across archosaur evolution in scientific terms, and (2) how we’ve completed a long-delayed project with stubbornness (and during a pandemic!), but also (3) how it bridges my past career from my PhD and postdoc to the present work with DAWNDINOS. We’re now forging well beyond what this new paper has done in terms of truly testing, as best we can (estimate) so far, how limb muscles of archosaurs functioned and evolved, and how these contributed to particular behaviours and performance (maybe even palaeoecology and evolutionary success/extinction?). The current paper is just simple modelling of muscle leverage, but leverage is only one (very important!) piece of muscle function and performance. With fully dynamic, anatomically integrative, physiologically and physically representative biomechanical computer simulations that predict what living and extinct archosaurs could or could not do, we can do even better. So watch for that! Hopefully it won’t all take 20 years, or 250 million.