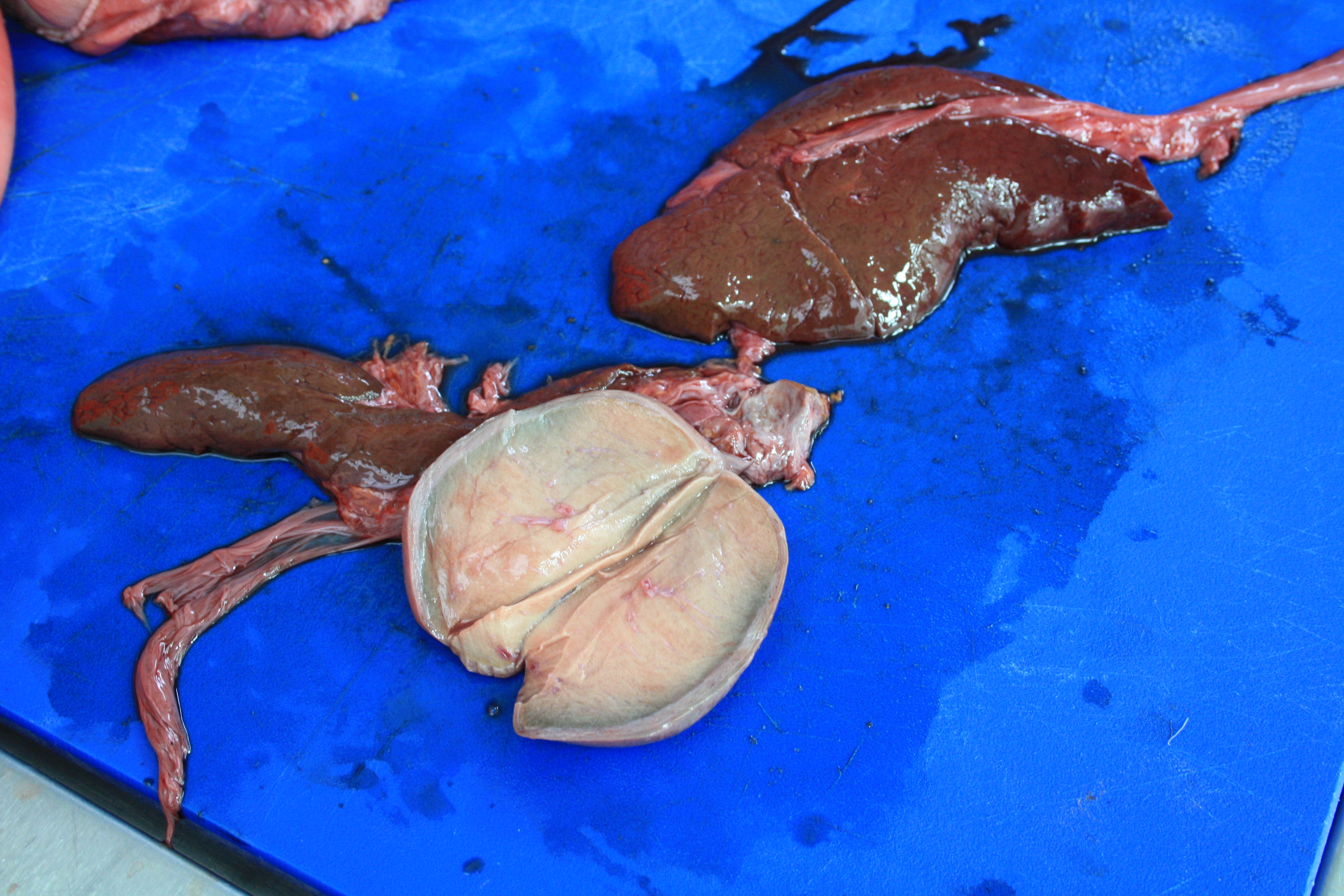
I’ve described our “Walking the Cat Back” Leverhulme Trust-funded project with Dr. Anjali Goswami and colleagues before, but today we really got stuck into it. We’re dissecting a 46kg male Snow Leopard (Panthera uncia) as the first “data point” (actually several hundred data points, but anyway, first individual) in our study of how limb and back muscles change with size in felids. No April Fools’ pranks here; real science-as-it-happens.
Stomach-Churning Rating: 7/10 for skinned leopard and globs of fat. Much worse in person, hence the downgrading from what could be a higher score. Don’t click the photos to emkitten them if you don’t want to see the details.
This leopard is the same one that Veterinary Forensics blogged about. It died in a UK cat conservation/recovery centre. Today is simply a short post, but it is the first in what will surely be a continued series of posts on felid postcranial anatomy and musculoskeletal biomechanics by our felid research team, with bits of natural history and evolution thrown in when we can manage. As befits one of my curt “Anatomy Vignette” posts, pictures will tell the story.
Skinned and mostly de-fatted snow leopard, with fat piled up on the lower left hand corner near the hind feet. Here we are identifying and then removing and measuring the individual muscles. Project postdoc Andrew Cuff is hard at work on the forelimb while I’m mucking around with the hindlimb. The fat here is about 3kg subcutaneous fat, so around 6.5% of body mass. And as the cat has been around for a while, that fat has gone a bit rancid and that is not nice. Not nice at all, no… Usually smells do not bother me, but this took some adjustment. Fortunately, the muscles are still OK, and work is coming along well.
UCL PhD student Marcela Randau, carving up our cat’s limb muscles. As usual in comparative biomechanics, we measure the “architecture”- parameters of the muscle that relate in a somewhat straightforward fashion to function. This muscular architecture includes things like muscle mass, the lengths of the fibers (fascicles) that make up the muscles, and the angle of the fascicles to the muscle’s line of action. These parameters correlate reasonably well with the force and power that the muscle can develop, and its working range of length change. Other posts here have discussed this more, but by measuring the architecture of many muscles in many felids of different sizes, we can determine how felids large and small adapt their anatomy to support their bodies and move their limbs. This will help to solve some lingering mysteries about the odd ways that cats move and how their movement changes with body size.
This research is being driven forward mainly by Andrew and Marcela, shown above, so I wanted to introduce them and our odoriferous fat cat. Upcoming dissections: 1-2 more snow leopards, tiger, various lions, ocelot, black-footed cat, leopard, and a bunch of moggies, and whatever else comes our way. All were EU zoo/park mortalities (there are a LOT of big cats out there!).
(John: here’s a guest post from my former PhD student, soon to be 100% legit PhD, Dr., and all that jazz, Julia Molnar!)
This is my first guest post, but I have been avidly following what’s in John’s freezer (and the blog too) for quite a while. I joined the lab in 2009 and left a month ago on the bittersweet occasion of surviving my PhD viva (oral exam/defense), so I’d like to take a moment here to thank John and the Structure & Motion Lab for a great 4 years!
Moving on to freezer-related matters; specifically, a bunch of frozen crocodile spines. It was late 2011, and the reason for the spines in John’s freezer was that John, Stephanie Pierce, and I were trying to find out more about crocodile locomotion. This was anticipated to become my first major, first-author research publication (but see my Palaeontologia Electronica paper on a related subject), and I was about to find out that these things seldom go as planned; for example, the article would not be published for more than three years (the research took a long time!). Before telling the story of how it lurched and stumbled toward eventual publication, I’ll give you some background on the project.
Stomach-Churning Rating: 3/10; x-ray of dead bits and nothing much worse.
A stumbly sort-of-bounding crocodile. They can do better.
First of all, why crocodiles? For one thing, they’re large, semi-terrestrial animals, but they use more sprawling postures than typical mammals. Along with alligators and gharials, they are the only living representatives of Crocodylomorpha, a 200+ million year-old lineage that includes wolf-like terrestrial carnivores, fish-like giants with flippers and a tail fin, even armored armadillo-like burrowers. Finally, crocodiles are interesting in their own right because they use a wide variety of gaits, including bounding and galloping, which are otherwise known only in mammals.
Nile crocodile skeletal anatomy
OK, so why spines? Understanding how the vertebral column works is crucial to understanding locomotion and body support on land, and inter-vertebral joint stiffness (how much the joints of the backbone resist forces that would move them in certain directions) in particular has been linked to trunk movements in other animals. For this reason, vertebral morphology is often used to infer functional information about extinct animals, including dinosaurs. However, vertebral form-function relationships have seldom been experimentally tested, and tests on non-mammals are particularly scarce. So we thought the crocodile spines might be able to tell us more about the relationship between vertebral morphology, mechanics, and locomotion in a broader sample of vertebrate animals. If crocodile spine morphology could be used to predict joint stiffness, then morphological measurements of extinct crocodile relatives would have some more empirical heft to them. Several skeletal features seem to play roles such as levers to mechanically stiffen crocodile spines (click to emcroc’en):
Anatomy of a crocodile vertebra
We decided to use a very simple technique that could be replicated in any lab to measure passive stiffness in crocodile cadavers. We dissected out individual joints were and loaded with known weights. From the movement of the vertebrae and the distance from the joint, we calculated how much force takes to move the joint a certain number of degrees (i.e. stiffness).
Me with crocodile vertebra and G-clamp
X-ray of two crocodile vertebrae loaded with a metric weight to calculate their joint’s stiffness
Afterwards, we boiled the joints to remove the soft tissues – the smell was indescribable! We took 14 measurements from each vertebra. All of these measurements had been associated with stiffness or range of motion in other studies, so we thought they might be correlated with stiffness in crocodiles also.
Some of the vertebral measurements that were related to stiffness
Despite my efforts to keep it simple, the process of data collection and analysis was anything but. I recall and exchange with Stephanie Pierce that went something like this:
Stephanie: “How’s it going?”
Me: “Well, the data are messy, I’m not seeing the trends I expected, and everything’s taking twice as long as it was supposed to.”
Stephanie: “Yes, that sounds like science.”
That was the biggest lesson for me: going into the project, I had been unprepared for the amount of bumbling around and re-thinking of methods when the results were coming up implausible or surprising. In this case there were a couple of cool surprises: for one thing, crocodiles turn out to have a very different pattern of inter-vertebral joint stiffness than typical mammals: while mammals have stiff thoracic joints and mobile lumbar joints, crocodiles have stiffer lumbar joints. Many mammals use large lumbar movements during bounding and galloping, so crocodiles must use different axial mechanics than mammals, even during similar gaits. While that’s not shocking (they did evolve their galloping and bounding gaits, and associated anatomy, totally independently), it is neat that this result came out so clearly. Another unexpected result was that, although several of our vertebral measurements were correlated with stiffness, some of the best predictors of stiffness in mammals from previous studies were not correlated with stiffness in crocodiles. The study tells a cautionary tale about making assumptions about extinct animals using data from only a subset of their living relatives or intuitive ideas about form and function.
Finally, the experience of doing the experiments and writing the paper got me interested in other aspects of crocodilian functional anatomy. For instance, how does joint stiffness interact with other factors, such as muscle activity and properties of the ribs, skin, and armor in living crocodiles? Previous studies by Frey and Salisbury had commented on this, but the influence of those factors is less tractable to experiment on or model than just naked backbones with passively stiff joints. In the future, I’d like to study vertebral movements during locomotion in crocodiles – especially during bounding and galloping – to find out how these patterns of stiffness relate to movement. In the meantime, our study shows that, to a degree, crocodile backbone dimensions do give some clues about joint stiffness and locomotor function.
To find out more, read the paper! It was just featured in Inside JEB.
Julia Molnar, Stephanie Pierce, John Hutchinson (2014). An experimental and morphometric test of the relationship between vertebral morphology and joint stiffness in Nile crocodiles (Crocodylus niloticus). The Journal of Experimental Biology 217, 757-768 link here and journal’s “Inside JEB” story
I Can’t Remember Freezermas… Can’t Tell Dissection from a CT. Deep down Inside I Feel to Freeze. These Wonderful Scenes of Anatomy! Now That the Week Is Through with Me, I’m Waking up; Ratites I see And There’s Not Much Left of These: Nothing remains but bones now
(digested from Metallica’s “One“, in …And Justice For All, the pummeling, slickly produced, huge-sounding, Jason Newsted-bass-playing leviathan of a thematic album(1988). It was all downhill for Metallica after this one, but it was a good year for rock! The song is about a soldier who had traumatic injuries and was left paralyzed, “locked-in” to his own mind. Themes/footage from “Johnny Got His Gun” (1939 book/1971 movie) are interspersed. Did you see this track coming? If so, you’re just as demented as I am; congrats!)
And so another year ends; we’re at the final post of Freezermas 2014: The Concept Album. We had 7 tracks involving leitmotifs of ostriches and cats and 2 vs. 4 legs, and CTs and x-rays, and epic dissections, and disturbing pathologies, and some twisted lyrics that mangled classic albums. There are so many more concept albums I could have touched on- great ones by Rush, Yes, Savatage, Helstar, Mastodon… many more. But I’ll give you a chance to sit in the DJ’s seat in this post!
Stomach-Churning Rating: 6/10. Some internal organs.
Today’s one mystery dissection photo is of two things, and the Mystery Anatomy challenge is to identify both (the 2-part brown thing and the 1-part whitish thing). They are from our friend the ostrich.
Your task is to weave your answer into the lyrics of a song from any concept album (2 lines or more)– you must identify the song, artist and album with your answer so we can figure out the tune. Any genre is OK as long as it is clearly a concept album (music, that is). You have freedom. Use it wisely! As always, bonus points for extra cleverness.
We’ll let Maytagtallica sing us out:
♫Hold my breath as I wait for points Oh Please John, blog more?♫
Freezermas continues! Today we have a treat for you. Lots of detailed anatomy! This post comes from my team’s dissections of an ostrich last week (~3-7 February 2014), which I’ve been tweeting about as part of a larger project called the Open Ostrich.
However, before I go further, it’s as important as ever to note this:
Stomach-Churning Rating: 9/10: bloody pictures of a dissection of a large ostrich follow. Head to toes, it gets messy. Just be glad it wasn’t rotten; I was glad. Not Safe For Lunch!
If the introductory picture below gets the butterflies a-fluttering in your tummy, turn back now! It gets messier. There are tamer pics in my earlier Naked Ostriches post (still, a rating of 6/10 or so for stomach-churning-ness there).
Research Fellow Jeff Rankin, myself and technician/MRes student Kyle Chadwick get to work, removing a wing.
This is a male ostrich, 71.3 kg in body mass, that had gone lame in one foot last summer and, for welfare reasons, we had to put down for a local farmer, then we got the body to study. We took advantage of a bad situation; the animal was better off being humanely put down.
The number for today is 6; six posts left in Freezermas. But I had no idea I’d have a hard time finding a song involving 6, from a concept album. Yet 6 three times over is Slayer’s numerus operandi, and so… The concept album for today is Slayer’s 1986 thematic opus “Reign in Blood” (a pivotal album for speed/death metal). The most appropriate track here is the plodding, pounding, brooding, then savagely furious “Postmortem“, which leads (literally and figuratively, in thunderous fashion) to the madness of the title track, after Tom Araya barks the final verse:
“The waves of blood are rushing near, pounding at the walls of lies
Turning off my sanity, reaching back into my mind
Non-rising body from the grave showing new reality
What I am, what I want, I’m only after death”
I’m not going to try to reword those morbid lyrics into something humorous and fitting the ostrich theme of this post. I’ll stick with a serious tone for now. I like to take these opportunities to provoke thought about the duality of a situation like this. It’s grim stuff; dark and bloody and saturated with our own inner fears of mortality and our disgust at what normally is politely concealed behind the integumentary system’s viscoelastic walls of keratin and collagen.
But it’s also profoundly beautiful stuff– anatomy, even in a gory state like this, has a mesmerizing impact: how intricately the varied parts fit together with each other and with their roles in their environment, or even the richness of hues and multifarous patterns that pervade the dissected form, or the surprising variations within an individual that tell you stories about its life, health or growth. Every dissection is a new journey for an anatomist.
OK I’ve given you enough time to gird yourself; into the Open Ostrich we go! The remainder is a photo-blog exploration of ostrich gross anatomy, from our detailed postmortem.
Hey, a short post here to say go check this new blog out! I love it. The first main post-introductory post is a dissection of a snow leopard, documenting a real vet case attempting to figure out why it died. The “Veterinary Forensics blog” is going cool places, and it is a kindred spirit to this blog. You might, as I do sometimes when walking into a veterinary pathology/postmortem facility, see surprising and rare stuff– like in this photo of urban foxes:
Jason Anderson, vertebrate paleontologist and anatomist at the University of Calgary (Canada), shared these two intriguing photos with me, and agreed for me to share them with you. Yay, thanks Jason! Good timing for a badly needed Mystery Dissection post:
What are they (species ID) *AND* from what region of the body?
(they are the same region, same specimen, same animal)
RULE: Your answer must be in the form of a winter holiday song (at least four lines)!
If you’ve been following some of my recent tweets, I’ve been tweeting about the “joys” of increased academic paperwork around holiday-time; e.g. this one:
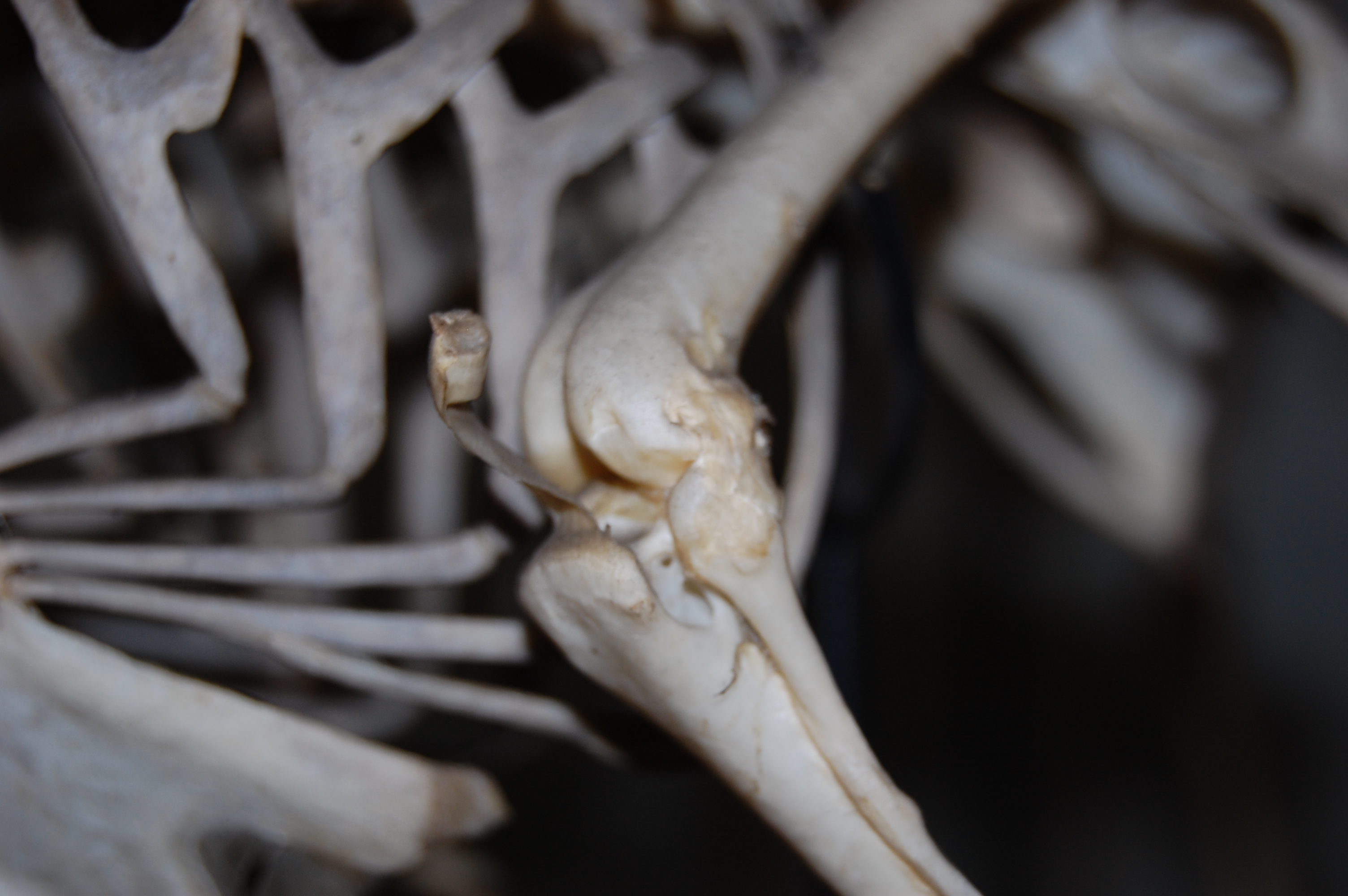
Here, I give you a long-planned post on the patella (“kneecap bone”) of birds, which was my Royal Society Senior Research Fellowship sabbatical project for 2012-13. This is only a brief introduction to the anatomical issues at hand, err, I mean at knee…
Stomach-Churning Rating: 6/10; mostly skeletons/fossils, but there are a few images of the dissection of a guineafowl, which is fresh and meaty.
Archaeopteryx, the Berlin specimen. Helluva fossil, but nary a patella!
The question I am exploring, first of all, is simply how the patella evolved, because it seems to be present in almost all living birds. However, it is absent in all non-avian dinosaurs, and indeed most Mesozoic birds, too. There is barely a hint of any precursor structure (a “patelloid”) in other reptiles, but lizards evolved their own patella that is quite different (a flattened lozenge, not a rectangular structure lying tightly confined in a “patellar groove” on the femur as it is in birds). Mammals evolved the knobbly, hemispherical kind of kneecap that we’re familiar with, possibly on several occasions (a different story!). So the patella evolved at least three times in the lizard, mammal and bird lineages– and possibly more than once in each of these groups. And that’s about it for almost 400 million years of tetrapod evolution, except for a few very rare instances in fossils and sort-of-patella-like things in some frogs or other weirdos.
Fossil birds exhibit no clear presence of a patella until we come very close to modern birds on the avian stem of the tree of life (see below). And then, suddenly in modern birds, there is a lot of variation and not much good documentation of what kind of patella exists. This makes it challenging to figure out if the patella is ancient for modern birds or if it evolved multiple times, or how it changed after it first evolved– let alone bigger questions of what the patella was “for” (performance benefits, functional consequences, etc.; and developmental constraints) in the birds that first evolved it.
Considering that the patella is such an obvious bone in some birds, and certainly affects the mechanics of the knee joint (forming a lever for the muscles that cross it; homologous to our quadriceps muscles) and hence locomotion, it is a compelling research topic for me.
What follows is a pictorial guide to the patella of some birds, in sort of an evolutionary/temporal sequence (see my earlier post for a recap of some major groups), with a focus on animals I’ve studied more intensively so far (with >10,000 species, there is a lot that could be done):
The early Cretaceous bird Gansus (from the IVPP in Beijing), represented by many beautifully preserved specimens, all of which lack a patella. This absence is characteristic of other stunningly preserved fossil Chinese birds, indicating that this is almost certainly an ancestral absence of a patella, until…
The famed Cretaceous diving (flightless) bird Hesperornis, from Wikipedia/Smithsonian. Note the massive, conical/crested patella in front of the knee (jutting up and overlapping the ribs/vertebrae close to the pelvis; see also below). That elongate patella is characteristic of many diving birds that use foot-propelled swimming; it has evolved many times in this fashion. Other hesperornithiform birds show some transformational states in their anatomy toward this extreme one.
Check this out! More Hesperornis (cast), with the femur on the left and the patella on the right. The bloody patella is almost as long as the femur! That’s nuts. With kind permission from the Natural History Museum, London.
Exhibited ostrich (Struthio camelus) skeleton in left side view showing the patella (white arrow), on exhibit atThe Natural History Museum at Tring, Hertfordshire, UK. Ostriches are remarkable in that they have this elongate patella (actually a double patella; there is a smaller, often-overlooked second piece of bone) and yet are rather basal (closer to the root of the modern avian family tree)– however, they obviously are specialized in ways other than this double patella, most notably their very large size, flightlessness, and elongate legs. So the unusual patella is more likely linked to their odd lifestyle than a truly primitive trait, at least to some degree (but stay tuned: what happened with the patella in other members of their lineage, the ratites/palaeognaths, is much less well understood!).
Note that ostriches and Hesperornis together hint that the presence of a patella might have been an ancestral trait for living birds, but their patellae are so different that the ancestral state from which they evolved must have been different, too; perhaps simpler and smaller. Hence we need to look at other birds…
Skinned right leg of a Helmeted Guineafowl, Numida meleagris, above. That whitish band of tissue in the middle of the screen, on the front of the knee, is part of what is concealing the patella. That is an aponeurosis (connective tissue sheet, like a thin tendon) of the muscles corresponding to our “quads” or our tensor fascia latae, detailed more below. Guineafowl are fairly basal and well-studied in terms of their bipedal locomotion, so they are an important reference point for avian form and function.
Right guineafowl leg, with patella exposed. Here I’ve peeled away that white band of tissue and associated muscles, which have been reflected toward the bottom of the screen (AIL and PIL labels corresponding to the anterior and posterior parts of the Iliotibialis lateralis muscle). The tip of the scalpel is contacting the patella. It’s not much to see, but lies atop the bright yellow fat pad that cushions it against the femur. You should be able to see a groove in the end of the femur just above that fat pad, which is where the patella sits and slides up and down as the knee moves/muscles contract. This is called the patellar groove, or sulcus patellaris.
Left leg of another guineafowl (with right tibiotarsus behind it, on the left) showing the patellae in articulation; in medial (inside) and cranial (front) views, respectively. The patella is the little rectangular bit of bone in the top middle of the screen, interposed between femur (thigh) and tibiotarsus (shank).With kind permission from the Natural History Museum, London.
Right leg of a Cape Penguin (Spheniscus demersus) from the University Museum of Zoology in Cambridge, showing the big lumpy patella in this wing-propelled diver. They still walk long distances on land, so presumably a patella plays some role in their gait, helping to explain its large size, which like the ostrich and Hesperornis seems to be a novel trait. Notice the groove across the patella, made by the tendon of the ambiens (like our sartorius/”tailor’s muscle”), which crosses from the inside to the outside of the leg via this route. This groove is often considered a useful phylogenetic character in modern birds, as its contact with the patella (sometimes via a hole, or foramen) varies a lot among species.
A hornbill, Buceros sp., from the UMZ Cambridge museum as well. This displays the possibly-more-typical, little rounded patellar nubbin that many birds have. See below for more.
Closeup of the knee/patella of the hornbill, Buceros sp., from above. Not much to squawk about, patella-wise, but it’s there.
And so we complete our quick tour of the avian patella, in its grand variation and humble beginnings.
Why does an ostrich have a patella and a Tyrannosaurus, Edmontosaurus or Triceratops did not? Why were birds the only bipedal lineage to evolve a patella (mammals and lizards gained a patella as small quadrupeds), and why did some bipeds like kangaroos “lose” (reduce to fibrous tissue, apparently) their patella?
These are the kinds of mysteries my group will now be tackling, thanks to a generous Leverhulme Trust grant on sesamoid bone ontogeny, mechanics and evolution. My group is now Dr. Vivian Allen as the postdoc, Sophie Regnault as the PhD student, and Kyle Chadwick as the technician and MRes student, along with numerous collaborators and spin-off projects. We’re looking forward to sharing more! But for now, I hope that I’ve engendered some appreciation for the avian patella, as the silly title indicates (“fella” used in the general sense of anyone!). This work is all unpublished, but some of this should be out in not too long, in much more lavish detail! Much as the patella is the “forgotten lever “of the avian hindlimb, it is the fulcrum about which a substantial part of my research group’s activity now pivots.
Hola from Barcelona, where 500ish of us are telling each other about the latest research in the field of morphology (like anatomy, but broader, deeper, more explanatory; but if you prefer to think of it as anatomy that’s OK by me)!
#ICVM and #ICVM2013 (favoured) are the hashtags, and http://icvm2013.com/ is the website, and there’s Facebook and all that too! You can read the full programme and abstracts here. It’s the best damn conference in the universe and I am not remotely biased. It happens every 3 years somewhere in the world and is always chock full of 5 days of glorious new information on animal form and function and much more, with just too many interesting talks to ever be able to take it all in.
I am speaking a few times and want to share a talk that is about sharing the glory of morphology in public.
Morphology research, that is; please put your clothing back on!
It’s a text-heavier talk than my rules-of-conference-talks normally would allow, but I’m going for it, as that makes it better for sharing because my dulcet tones will not accompany the version I am sharing online. Someday in the future, at a conference venue that is better set up for reliably live-broadcasting a talk (this is NO FAULT of the excellent organizing committee of ICVM/ISVM!), I would just do it live, but not today, not here.
The point of the talk should be obvious from the first slide (as in my last post). But I’ll presage it by saying that another subtext, which might not come through so strongly in the slides as opposed to my spoken words, is that we need to tell people that we’re doing morphology/anatomy research! We should not be shy of that label because deans or geneticists or conventional wisdom or what/whomever might say (very, very wrongly!) that it is a dead or obsolete science.
While natural history, evolution, palaeontology and other fields allied to morphology do pretty well in the public eye, I don’t see people often reminded that what they are being told about in science communication is a NEW DISCOVERY IN ORGANISMAL MORPHOLOGY and that we are still discovering such new things about morphology all the freaking time! (e.g. my team’s research on elephant false sixth toes, or Nick Pyenson‘s team’s research on whale chin sense organs to name just 2 such studies, both published on the same day in Science!)
Indeed, many of those discoveries such as new fossils/exotic living things with cool features, cool developmental mechanisms that produced complex structures, or insights into how organisms are able to do amazing things are implicitly morphological discoveries, but the fanfare too often goes to natural history, palaeontology, evo-devo or some other area rather than explicitly to morphology.
In contrast, I too often hear people poo-pooing anatomical research as yesterday’s science.
Vesalius’s classic skeleton (from Wikipedia), which is great but to me also conjures misleading connotations of anatomy as a defunct discipline that old dead dudes did.
We need to sell ourselves better not only in that regard, of a renaissance of discoveries and insights in our field, but also in the sense of being in a renaissance that is driven by TOTALLY AWESOME TECHNOLOGICAL AND METHODOLOGICAL ADVANCES, especially computerized tools. We’re just as fancy in terms of techy stuff as any other biologists, but we don’t shout it from the rooftops as much as other disciplines do.
We’re not just primitive scientists armed only with scalpels and maybe a ruler now and then, although that simple approach still has its sublime merits. We’re building finite element models, running dynamic computer simulations, taking high-resolution CT or synchotron scans, manipulating embryos, digging up fossils, sequencing genes– you name it, morphologists may be doing it! (For similar views see Marvalee Wake’s recent review of herpetology & morphology; I’m by far not the first person to make the arguments I’m making in my talk, but I am putting a personal spin on them)
And of course, as the talk is being delivered by me, you might rightly expect that I’ll say that we need to do more of this kind of cheerleading where we have maximal visibility and interaction, which includes online via social media, etc. I’ll discuss one other venue which has featured prominently here on this blog, too: documentaries. Oh I’m not done with that hobby horse, no sirree, not by a long shot!!
Anyway I should get back to preparing my talk but here is the link to the slideshow (props to Anne Osterrieder for the inspiration to put my slides up here):
Please discuss anything related to this topic in the Comments– I’d love to hear what you think!
I am happy to clarify what my shorthand notes in the slide text mean if needed. There are links in the talk to other sites, which you can click and explore.
The Unfeathered Bird book by Katrina van Grouw proclaims immediately in its Introduction that it “is not an anatomy of birds.” True– it is far more than that, and it would be a shame if it had just been a dry, technical avian osteology reference book. It is a unique blend of art and science- particularly avian anatomy, evolution, taxonomy, natural history and more. The Unfeathered Bird is written for a general audience; birders/twitchers or just natural history buffs would be ideal targets of its unfettered passion for all things avian. A 12-year-old who is very keen on animals could enjoy it, and it may ignite the flames of ornithological excitement in many young or older readers. I am glad it was not called “The Naked Bird” as that would have caused some serious misconceptions (badum-tish!). The book is dripping with illustrations (at least one every two pages, often more). Almost all of the illustrations (except some paintings in the style of the cover) are in the same brownish sketch style that, like much of the book, evokes a bygone era of dark wooden cabinets and shadowed halls packed with skeletons, with nary an interactive graphics display, animatronic dinosaur or hyperdetailed cladogram in sight. It feels like an homage to the Victorian naturalists’ joy for anatomical detail conveyed through painstakingly detailed woodcuts. And while many still think of feathers as “the defining feature of birds,” enough about feathers already. Seriously. This is a book is about what lies beneath, and how all that non-fluffy stuff is important for birds’ lives, too.
(image-intense post; all can be clicked to embiggritate)
Katrina with peacock feather headdress? (back cover pic and rear view of same skeleton)
Katrina with front cover framed pic and the peacock skeleton that went with it.
The Introduction continues to explain that the book is truly about how the external anatomy of birds is linked to the bony anatomy, which might remind astute readers of modern approaches like the extant phylogenetic bracket. The rest of the book uses both skeletal and unfeathered, quasi-myological illustrations to get this point across vividly. The explanatory text is written at a basic enough level for the average reader and is just the right length, with interesting anecdotes and natural history facts that even the expert reader will find interesting or even inspirational (e.g. possibly a goldmine for research ideas). First there is a 26 page “Basic” section with an introduction to avian osteology, with bountiful sketches to illustrate key organs and text explaining how it all fits together in the fully accoutered bird. The decision to use classical Linnean taxonomy (defunct or re-arranged taxa from the Systema Naturae like Accipitres, Picae, Anseres, Grallae, Gallinae and Passeres; which are the six “Specific” chapters in the second section of the book) was a good one- it enhances the classical feel of the tome and gives the author a great opportunity to discuss convergent evolution and how that misled past ornithologists.
But for me, the book is most pleasurable for the visualizations and the passion for all things birdy that weaves through them and the accompanying text. The removal of feathers, or even all soft tissues, from bird bodies (posed in naturalistic behaviours) that van Grouw renders in her illustrations shows birds in a new light, emphasizing the strangeness and diversity that lie beneath. The author begins the book with a touching Acknowledgments section in which her husband Hein van Grouw, curator of birds at the Natural History Museum’s Tring collection, features very prominently, making it clear that the book was a team operation and comes from the heart after a 25-year journey. This gives the book a special warmth that is preserved throughout the remainder- although the illustrations are of flayed bodies or boiled / beetle-macerated skeletons, the tone is nothing less than an earnest love for birds of all kinds, and a zest for portraying those feelings to the reader in sketches and prose. It is a joyous celebration, not a somber litany, of the wonder of birds that can be gleaned from dead bodies. There is so much powerful, awesome imagery stuffed into those pages that it is hard to summarize. I’ll let five of my favourite images from the book (more are in her gallery and her book’s Facebook page; but even these are just the tip of the icebird) help get this across (used with permission of the author):
Naked kiwi in action.
The unscaled bird: guineafowl feet.
Deplumed sparrowhawk with dove trophy, exalting in its triumph.
Budgerigar has made a friend? Or came to grips with its own mortality?
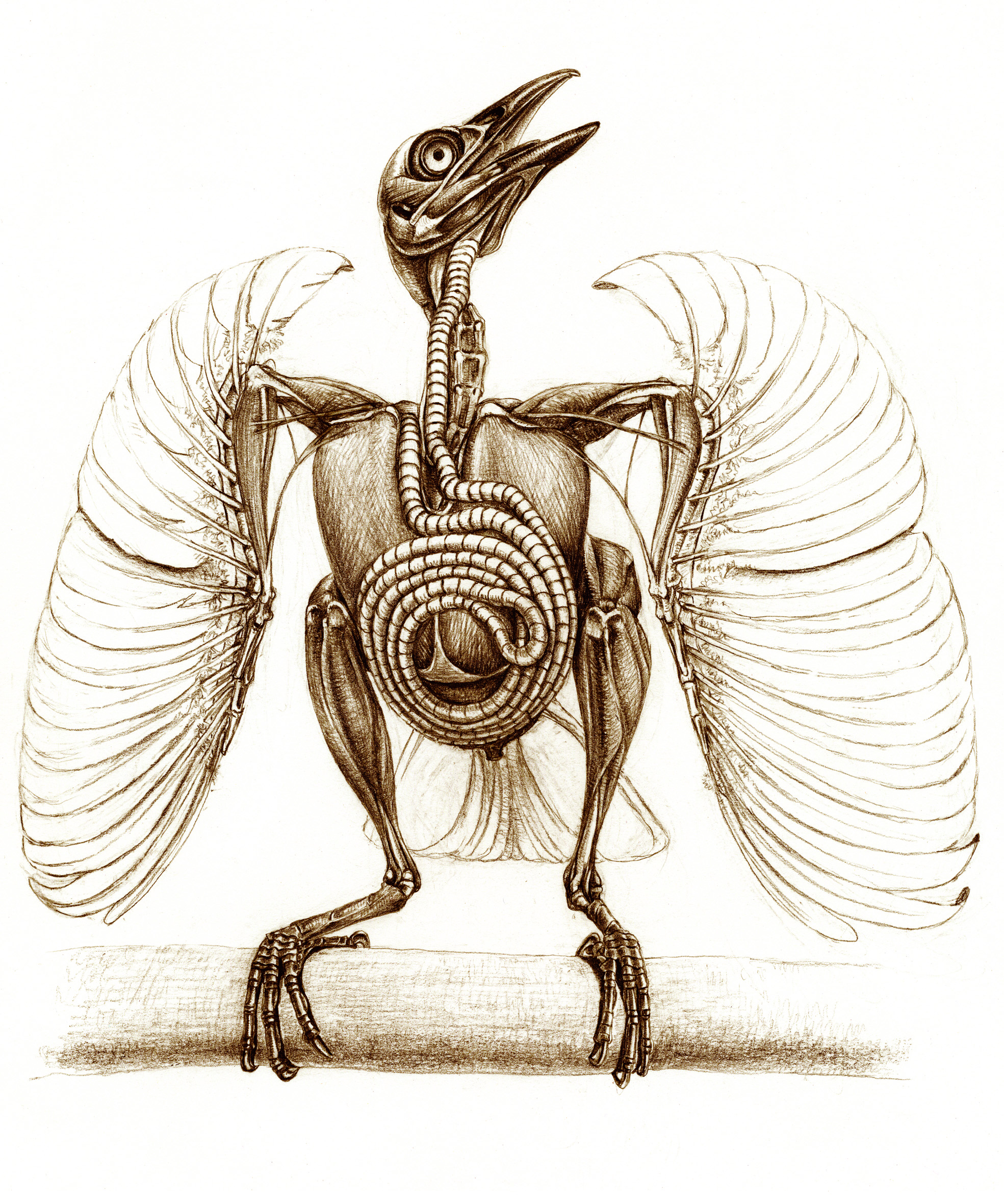
Trumpet Manucode’s WTF anatomy! Spiraling tracheal coil made me gasp in awe when I saw this image in the book.
Now I’ll depart from this post just being a book review. I went to the Tring collection to do some research, and arranged my trip so I’d also get to see the debut of a Tring special exhibit featuring The Unfeathered Bird, and also to meet Katrina as well as Hein van Grouw. The placement of the exhibit at Tring is apropos, because Katrina was a curator at the museum until a few years ago and Hein still is. But the inspiration for the work and the specimens used (with a few exceptions, including from other museums) are Katrina’s. She (with Hein’s help) procured bodies of birds to dissect, macerate and sketch for the book over its 25 year fledging period, noting in the Acknowledgments that “no birds were harmed” to do this– do read those acknowledgments, as there are some amusing tales there of how she obtained some specimens.
I was fortunate to be able to take some photos of the exhibit while they set it up, and grabbed some candid images of Katrina and colleagues during that process. The following images show off the exhibit, which is all in one clean, bright, simply adorned room in the Tring that lets Katrina’s framed sketches be the focus. Here are some examples:
Poster advert for the book in the Tring collections.
Tring exhibit setup, with Katrina, husband Hein, and helper finishing it up.
Tring exhibit now ready.
Tring exhibit case.
Framed sketches at Tring exhibit.
More framed sketches at Tring exhibit.
The exhibit is fun for people who are already Unfeathered Bird fans, and a good way of drawing in new ones. The book is a precious thing that any fan of birds, especially scientists, really needs to have a hard copy of. While it claims not to be an anatomy text, its illustrations provide ample opportunities to use it for that purpose. But really the point of owning all 287-plus pages is to bask in the warmth of true, pure appreciation for classic ornithology, which I found infectious. It is a book by and for bird lovers, but also for those that find the interface of art and science to be fascinating.
I confess I used to hate birds. I found them annoying and boring; all that flitting and twitting and pretentious feathers. “Get over yourselves, already, and calm down too!” was my reaction to them. When I started grad school, I had an open disdain for birds, even moreso than for mammals (OK, except cats). I was a “herp” fan through and through, for most of my life (childhood spent catching anoles in Florida, or stalking frogs in Ohio; during visits to my grandparents). What won me over was studying birds (and eventually mammals, too) as a young scientist, and learning how incredible they are– not just as endpoints in the story of theropod dinosaur evolution, as my thesis focused on, but as amazing animals with spectacular form-function relationships. The Unfeathered Bird is saturated with that amazement, so we’re birds of an unfeather.
Framed sketch of dodo head at Tring exhibit.
Entirely unfeathered Indian peafowl in matching views.
Painted Stork and Great Hornbill sketches.
Red junglefowl, wild ancestor of domestic chickens (and the book ends with several such breeds illustrated).
Katrina told me that she is already deep into writing the next book, whose subject I won’t spoil for you here but maybe we will be lucky enough to have her appear in the Comments and plug it? 🙂 (Her website does say “It was Hein’s stroke of genius to include domestic birds and they’ve provided the inspiration for my next project.” so the cat is out of the bag and amongst the pigeons!) It is great to hear that the book has done quite well sales-wise and critically, such as ~#67 on the Amazon sales list at one point– I hope this paves the way for more such books not only from Katrina, but from others engaged in lateral thinking (and still others) on the boundaries of science-art.
I’m letting the dogs out today. Science gone barking mad! Hopefully my puns will not screw the pooch.
Stomach-Churning Rating: 4/10; a dog cadaver’s leg (not messy), then just tame digital images of anatomy.
I am working with Rich Ellis, a former MSc student at Univ. Colorado (see his cool new paper here!), for a fun new collaboration this year. He was awarded a prestigious Whitaker Foundation scholarship to do this research, which focuses on how different animals stand up from a squatting position, with the legs about as bent as they can be.
We want to know how animals do this standing up movement, because it is in some ways a very demanding activity. Very flexed/bent limb joints mean that the muscles (and some tendons) are stretched about as far as they ever will be. So this places them at disadvantageous lengths (and leverage, or mechanical advantage) for producing force. We know almost nothing about how any animal, even humans, does this-– how close to their limits of length are their muscles? Which muscles are closest? Does this change in animals with different numbers of legs, postures, anatomy, size, etc? Such fundamental questions are totally unaddressed. It’s an exciting area to blaze a new trail in, as Rich is doing. So far, we’ve worked with quail, humans, and now greyhounds; in the past I did some simple studies with horses and elephants, too. Jeff Rankin from my team and other collaborators have also worked on six species of birds, of varying sizes, to see how their squat-stand mechanics change. Thus we’ve covered a wide diversity of animals, and now we’re learning from that diversity. “Diversity enables discovery,” one of my former PhD mentors Prof. Bob Full always says. Too true.
Greyhounds are interesting because they are medium-sized, long-legged, quadrupedal, quite erect in posture, and very specialized for fast running. Fast runners tend to have big muscles with fairly short fibres. Short fibres are bad for moving the joints through very large ranges of motion. So how does a greyhound stand up? Obviously they can do it, but they might have some interesting strategies for doing so- the demands for large joint motion may require a compromise with the demands for fast running. Or maybe the two demands actually can both be optimized without conflict. We don’t know. But we’re going to find out, and then we’ll see how greyhounds compare with other animals.
To find out, we first have to measure some dogs standing up. We’ve done that for about 8 greyhounds. Here is an example of a cooperative pooch:
Those harmless experiments, if you follow me on Twitter, were live-tweeted under the hashtag #StandSpotStand… I dropped the ball there and didn’t continue the tweeting long after data collection, but we got the point across– it’s fun science addressing useful questions. Anyway, the experiments went well, thanks to cooperative pooches like the one above, and Rich has analyzed most of the data.
Now the next step involves the cadaver of a dog. We could anaesthetize our subjects and do this next procedure to obtain subject-specific anatomy. But it really wouldn’t be ethically justified (and if I were an owner I wouldn’t allow it either!) and so we don’t. A greyhound is a greyhound as far as we’re concerned; they’ll be more like each other than either is like a quail or a human. Individual variation is a whole other subject, and there are published data on this that we can compare with.
We get a dead dog’s leg — we don’t kill them; we get cadavers and re-use them:
We study the hindlimb because birds and humans don’t use their forelimbs much to stand up normally, so this makes comparisons simpler. We’re collecting forelimb data, though, as we work with quadrupeds, for a rainy day.
We then CT scan the leg, getting a stack of slices like this– see what you can identify here:
It’s not so clear in these images, but I was impressed to see that the muscles showed up very clearly with this leg. That was doggone cool! Perhaps some combination of formalin preservation, fresh condition, and freezing made the CT images clearer than I am used to. Anyway, this turned out to be a treat for our research, as follows.
We then use commercial software (we like Mimics; others use Amira or other packages) to “segment” (make digital representations in 3D) the CT scan data into 3D anatomy, partitioning the greyscale CT images into coloured individual objects– two views of one part of the thigh are shown below.
What can you identify as different colours here? There are lots of clues in the images (click to embiggen):
And here is what the whole thigh looks like when you switch to the 3D imaging view:
Quite fetching image, eh?!
The next steps after we finish the limb segmentation are to apply the experimental data we observed for greyhounds of comparable size by importing the model and those data into biomechanics software (SIMM/OpenSim). We’ve done about 40 models like this for various species. I detailed this procedure for an elephant here.
Then, at long last, science will know how a greyhound stands up! Wahoo! Waise the woof! Stay tuned as we hound you with more progress on this research-as-it-happens. Rich just finished the above thigh model this week, and the rest of the leg will be done soon.
Many thanks to Rich Ellis for providing images used here. And thank you for persevering my puns; they will now be cur tailed.
Happy Freezermas! Sing it: “On the fifth day of Freezermas, this blo-og gave to me: one tibiotarsus, two silly Darwins, three muscle layers, four gory hearts, a-and five stages modelling a doggie!” ♪♫