…a daily picture of anatomy! And today it is three pictures, scoob-a-dee!
Welcome back again to Freezermas!
For the previous days of Freezermas we first had 1 picture, then 2, now guess how many we have today? Right, we’ve settled into a groove and have three (plus one silly one). Today is fresh beefy anatomy day! No focus on bones, but on soft tissues– however, once again, I’m representin’ bird legs! And this time, no mystery things to identify; sorry. But if you want to muscle in on some myology, today is the day for you. I will unwrap the thigh of an ostrich and consider the major muscles that power rapid running in this biped, and how they illuminate the evolution of bipedal motion along the line of descent to birds. For more ostrich escapades, see this old post. And we’re off!
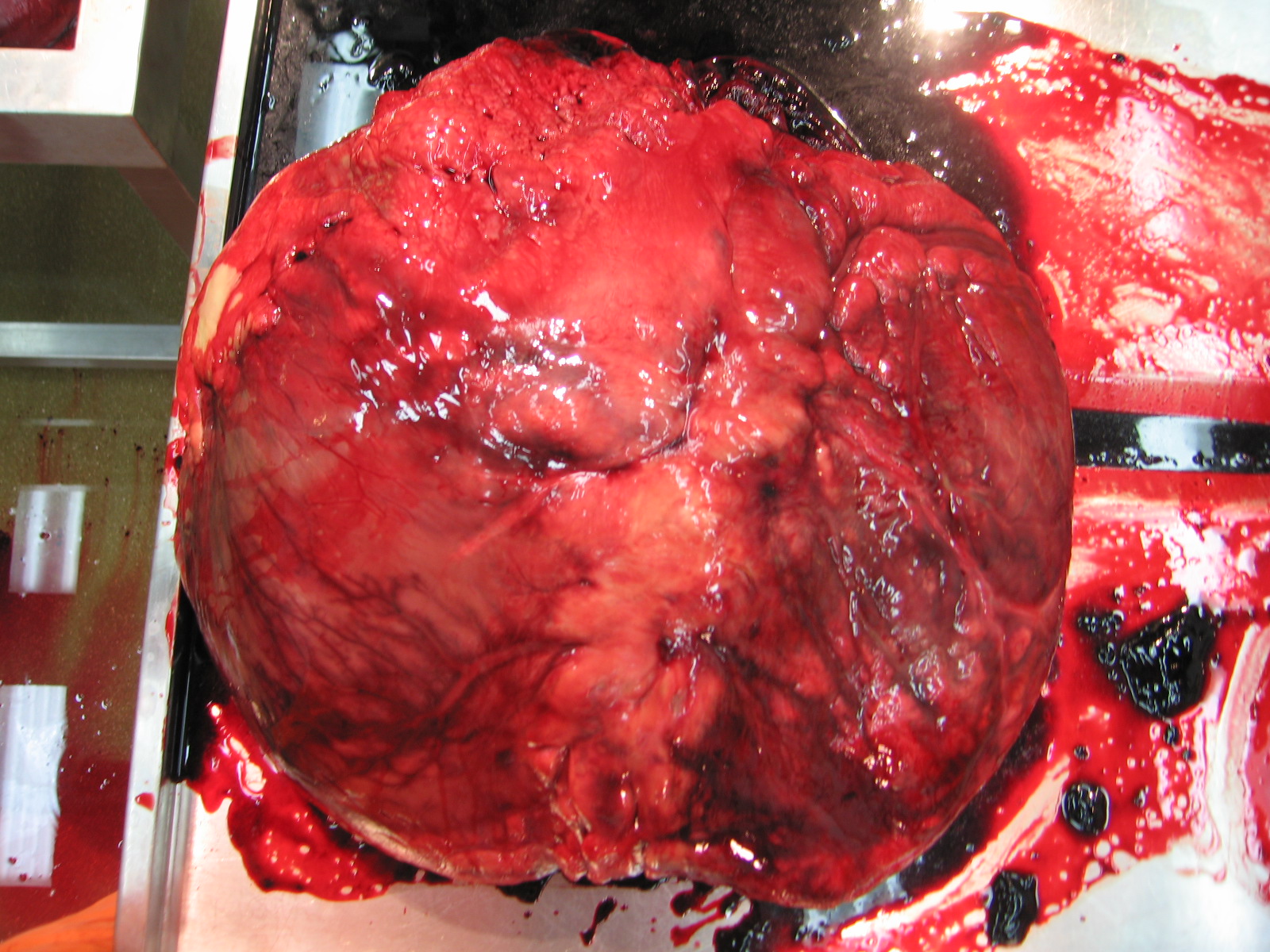
Stomach-Churning Rating: 7/10; plenty of fresh, red, meaty meat from ostrich leg muscles.

Here you are looking at a right hindlimb of an ostrich, in side/lateral view. To help orient yourself, the hip lies deep in the middle of the image and the knee is the rounded bump near the bottom right corner, with the shank angling sharply back toward the bottom left.
I’ve labelled six muscles in yellow. As usual for sauropsid (bird/reptile) pelvic limb muscles, they have sensible names that reflect their attachments. They don’t have so many silly old mammalian names like pectineus or latissimus, which tell you rather little about the muscles themselves. We can thank 19th century anatomists like two of my anatomist heroes, Hans Gadow and Alfred Romer (who refined Gadow’s earlier work and made it more popular among English-speakers and palaeontologists), for that enlightened nomenclature.
The six muscles seen above are the IC (iliotibialis cranialis), IL (iliotibialis lateralis), “AMB2?” (one of the ambiens muscles– correctly identified; ignore the ?), ITC (iliotrochantericus caudalis), CFP (caudofemoralis pars pelvica) and FCLP (a mouthful to say: flexor cruris lateralis pars pelvica). The ambiens is the one oddly, non-anatomically named muscle, and has nothing to do with helping you sleep (pssst– wake up! Muscles are exciting!), but everything to do with the state of total awesomeness, which is what “ambiens” means. Maybe. Or I am making shit up.

The IC, IL and AMB2 are parts of the triceps femoris group (discussed in my 1st Freeezermas post), or for mammal fans the quadriceps. The IC and AMB are in front of the hip so they flex it (move the thigh forward; protract it); the IL is right around the hip so it can flex or extend the hip (protract/retract the femur); all three of these can extend (straighten) the knee joint to varying degrees. The IC is fairly typical for a bird except for its size, and helps to quickly swing the leg through the air between steps. Some birds have multiple parts of the IL, but ostriches and many others have simplified it to one major mass; regardless, it is a major muscle used to support the weight of the body.
The AMB2 is a remarkable muscle unique to ostriches; it can also be called the dorsal ambiens muscle. Typical birds just have a single head of the AMB sitting on the preacetabular (pubic) tubercle, so in front of and below the hip. It has a crazy tendon that snakes past the knee (in some birds, perforating/grooving the patella) into the lower leg muscles and may be able to even pull on the toes. But ostriches, for some reason, added a second head of this muscle that was shifted way up onto the front of the pelvis (the ilium; dorsal bladelike bone). Crocodilians also have a 2nd ambiens muscle but in a different position, and almost certainly as an example of convergent evolution. The function of the ambiens is mysterious, but this muscle has featured prominently in avian systematics/taxonomy, evolution (invoked as a key muscle used in perching) and more.
These muscles of the triceps femoris group are easily identifiable in crocodiles and other reptiles because they are remarkably similar in their attachments. The main changes these muscles experienced during the evolution of bipedalism, dinosaurs and later birds are simply proportional– they got bigger, with stronger, larger attachments on the pelvis and the front of the knee (the CC/LC, if you remember from Freezermas day 1).
The ITC is a muscle that is very dear to me. I’ve written a lot about it, and I love saying the name “Iliotrochantericus caudalis”- it is musical to me. For mammal fans, think gluteal muscles (medial gluteal in particular). It is a huge, pennate muscle (short and strongly angled muscle fibres in a “sandwich” with a tendinous sheet between the two layers of fibres). It has a short, broad tendon that wraps around the trochanteric crest (a structure on the upper front end of the femur with a history that goes wayyyy back into dinosaurs; long story!) to insert in a scarred depression. The ITC seems to mainly rotate the femur around its long axis to help support the body. I could go on and on about this muscle, which is part of the enigmatic “deep dorsal” thigh muscle group — the homologies of this group among land vertebrates are still controversial and confusing. But I will spare you the on-and-on. Incidentally, the ITC is the “oyster” in birds that is the best cut of meat. And in ostriches it makes a massive steak.
The CFP also has a cool evolutionary history. It runs from the back of the pelvis to the middle of the femur, closely adjoined to the caudal head of the muscle (CFC), which is more vestigial. In birds the CFP is usually not a large muscle, but in other sauropsids/reptiles it can be fairly hefty, although almost never as hefty as its more famous counterpart the caudofemoralis longus (= CFC in birds). Probably any dinosaur specialist is familiar with its origin and its insertion: respectively, the “brevis fossa” on the back of the ilium; a big shelf of bone; and the fourth trochanter of the femur; a crest of bone that is reduced to a scar/tubercle in birds. Much like its tail-based counterpart, the CFP became progressively reduced closer and closer to birds. This is related to a reduction in the amount of movement of the femur/thigh during locomotion, as birds shortened their tails and shifted their balance forward, as Steve Gatesy showed in a classic 1990 paper. Hopefully there will be more about this subject in a future paper from my team…
The FCLP is another muscle that didn’t change much, except by getting larger as we trace its evolution from early reptiles to birds. It is a “hamstring” muscle that is an important power source during locomotion in birds like the ostrich, because it retracts the lower limb (flexes the knee; hence flexor cruris in its name) as well as the femur/thigh (extends the hip). Your semitendinosus muscle is a good comparison to it. Indeed, these two differently named muscles are homologous– our very distant tetrapod ancestor had the same single muscle, and its descendants didn’t change it that much on our lineage or on the avian/reptile one.

I’ve reflected the IL muscle out of the way so we can see the second layer of muscles underneath it. Now we see two more muscles of the thigh, and large ones at that– the FMTL (femorotibialis lateralis) and ILFB (iliofibularis).
The FMTL simply is a part of the triceps femoris group that only comes from the femur and hence only, but due to its large size powerfully, straightens the knee. Unlike the other muscles in this group, it has no action about the hip joint. It is very similar to your vastus lateralis muscle: its fleshy origin dominates the surface of the femur (thigh bone). There are two other parts of that muscle, hidden in this figure, much like our vastus group has multiple parts. Again, this is a muscle that enlarged on the lineage leading to modern birds.
And that evolutionary enlargement applies, too, to the ILFB, whose prominent insertion I discussed on day 1 of Freezermas. This huge “biceps” muscle (it is single-headed unlike in humans, so the name “biceps” does not apply well) is the most powerful of the “hamstring”-type muscles that extend the hip and flex the knee. Therefore it is important for the “knee-driven” locomotion of birds. And hence the ILFB enlarged during avian evolution– which is very evident from changes of both its bony origin on the back of the pelvis/ilium and its insertion on the fibula.

Here, for the terminus of today’s trio of struthious tributes and tribulations, I’ve moved the ILFB out of the way so you can see the various inner/medial layer of thigh muscles. Some of the former muscles are more exposed now, and we can see three new ones: the FCM (flexor cruris medialis), PIFM+(PIF)L (the tongue-twisting puboischiofemoralis medialis et lateralis), and tiny ISF (ischiofemoralis).
The FCM (~mammalian semitendinosus) is merely another, smaller part of the FCLP’s “hamstring” group, and its thin tendon blends with that of the FCLP, so it very much works with that muscle in locomotion, and has a similar evolutionary history.
The PIFM+L are “adductors”, but in birds they don’t really do any adduction (drawing the legs inwards) because they are right behind, rather than below or inside, the hip. They act as hip extensors/retractors of the femur, and probably aid more in holding the femur steady (“postural muscles”) than playing a major role in producing power for locomotion like the ILFB/hamstring group does. In earlier reptiles, they were much more important, for preventing the legs from splaying too far away from the body.
The ISF is usually quite a large muscle in birds, but ostriches and some other ratites have reduced it to a thin slip of muscle– often mistaken for other muscles (indeed, like a few other muscles I’ve described here, modern anatomists still get confused by this muscle– an otherwise superb recent description by Gangl et al., among others, mis-identifies this and some other muscles— an error an upcoming paper from my group will rectify). Normally the ISF sits atop a bone-free window on the outer surface of the pelvis, the ilio-ischiadic fenestra (literally a window in Latin) in birds; in ostriches it has moved more onto the ischium. In contrast, in other sauropsids it lies inside the pelvis, so during its evolution it became more lateral, but the insertion on the upper femur was maintained. It is a weak rotator and extensor of the hip, especially in ostriches in which its role is probably proportionately puny.
And there you have read a healthy chunk of my 2001 PhD thesis, condensed into less jargonious language. You might now know almost half of the key muscles of the avian hind limb. If you made it this far, you are one awesome anatomical enthusiast. If you eat meat, apply this lesson to the next chicken thigh you consume, to consumate this enthusiasm.
A broader point I’d like to make here is that anatomy is best conveyed not only along with the functional narrative (How does anatomy work?) but also the evolutionary tale (Where did anatomy come from and What were the consequences of its changes? Why did it change?). This takes it away from dry memorization of terms and locations, and carries it into the realm of explaining why nature is the way it is, and how every organism’s biology has a richly detailed and complex background. This style portrays nature as much more like that tangled bank that Darwin so enchantingly envisioned. I’ve tried to do that justice here, using this one ostrich whom we affectionately called Twinkletoes, or Twinkie, when we dissected it back in 2002.
Happy Freezermas! Sing it: “On the third day of Freezermas, this blo-og gave to me: one tibiotarsus, two silly pictures, a-and three muscle layers from Twinkie!”
Read Full Post »