I still have my original photocopy, from my grad school days circa 1996, of the 1983 Ted Garland classic paper “The relation between maximal running speed and body mass in terrestrial mammals”, festooned with my comments and highlighter pen marks and other scribblings. That paper remains the backbone of many research questions I am interested in today, and I often think about its underlying concepts. Here’s the key scatterplot from that paper, which I could almost replot by hand from memory, it is so full of implications (and can be clicked to embiggen it, perhaps even speedily depending on your internet connection):

Stomach-Churning Rating: 1/10; data and their ramifications; offal-free.
The major points (IMO there are less exciting ones about which theoretical scaling model the data best fit) of the paper are: (1) the fastest-running mammals are neither the smallest nor largest, but those around ~100 kg body mass; (2) if you fit a linear equation to the data (see above; hashed line), it seems like speed increases with body mass linearly (with no limit to that increase, within the body mass range of the data), but if you analyze individual groups of mammals they either don’t change speed significantly with size or they get slower– refer back to point #1 and the polynomial regression that is shown in the figure above (curved line). That’s the biological-question-driven science at the core of the paper (with some methods-y questions at their foundation; e.g. should we use a linear or polynomial regression to fit the data? The latter fits best, and gives a different answer from the former, so it matters.).
But what also fascinates me is the question of data. As the author, who taught me Evolution as an undergrad at U Wisconsin (this had a big impact on me), fully admits in the paper, the ~3-page table of data “necessarily sacrifices some accuracy for completeness”. This paper is about a big question, how mammal speed changes with size, and so its big question explicitly allows for some slop in the data (I will return to this issue of slop later). But given that very few of the data points have very accurate measurements for speed, or for body mass for that matter, how much can we trust an x-y plot of those data, no matter what method is used? Oh there is so much opportunity here for geeky pedantry and niggling scrutiny of data points, true, but hold on…
Plenty of follow-up papers have mused over that latter question, and spin-off ones. Here are some of their plots, re-analyzing the same or very similar data in different contexts. A look at how these papers examine these data and related questions/methods leads into some avenues of science that fascinate me:
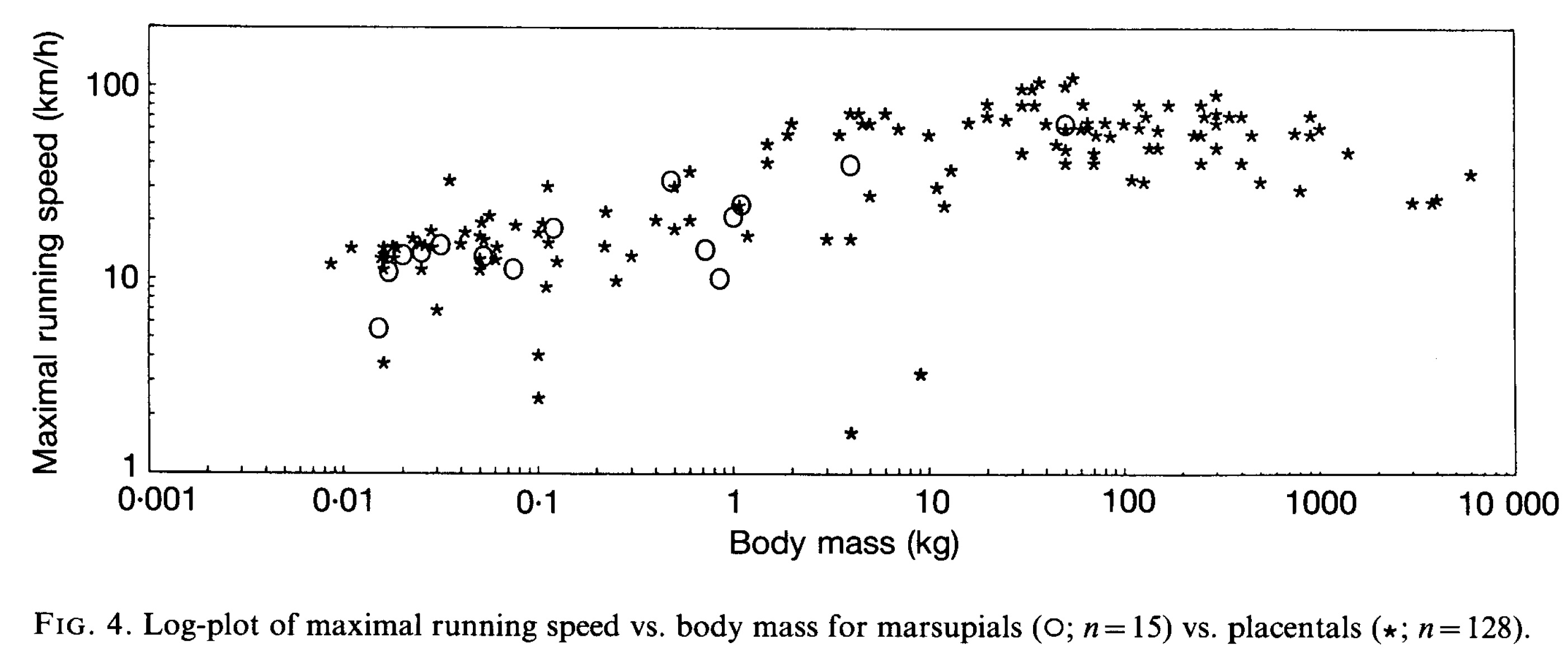
Garland and Baudinette (link to pdf here) checked whether placental (i.e. most; including us) mammals could run/hop faster than marsupial (pouched; e.g. kangaroos) mammals. Their results said “not really”, as the plot intimates. Scatter in the data, especially between 0.01-10 kg, confounds the issue- there’s a lot of specialization going on (notably, animals that are very slow for their size, e.g. sloths). But marsupials are not, as had been suggested before, inferior to placentals in some basic way such as running ability.
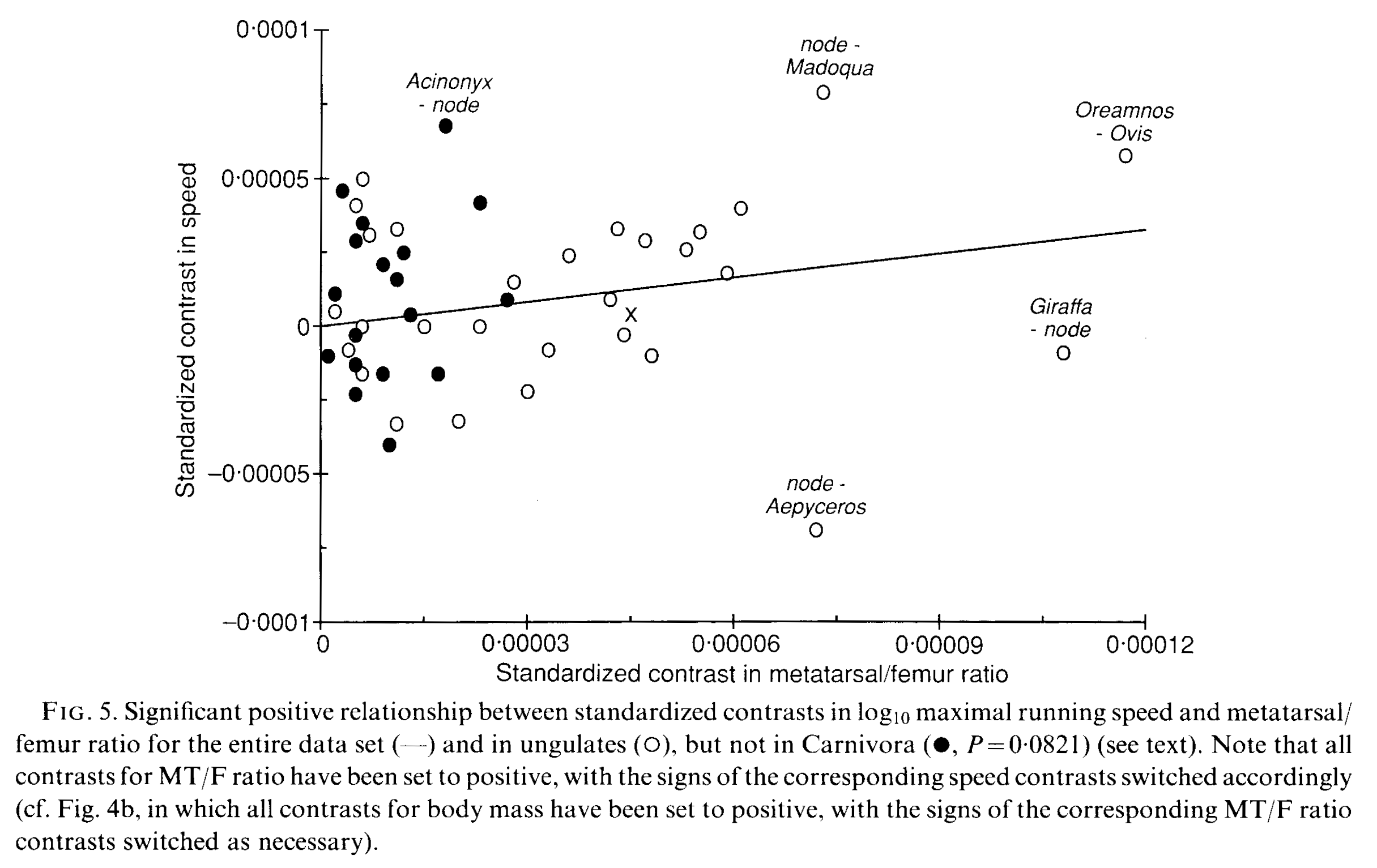
Above, Garland and Janis 1993 (link to pdf here) examined how the ratio of metatarsal (“sole bones” of the lower end of the leg/foot) vs. femur (thigh bone) length relate to speed, with evolutionary relationships taken into account. The methods (“independent contrasts” and its conceptual kin; I won’t delve into that morass more here!) did not exist for looking at phylogeny’s effects on the results in Garland’s 1983 paper. Yet “cursoriality” (relative elongation of the lower limb) had been thought to relate to running speed for over 80 years at that time, so that was what they tested: how much does limb-elongation correlate in a positive way with maximal running speed? They found that the answer was “sort of”, but that other things like home range size, energetics, ecology, etc. might explain as much/more, so caveat emptor. And by looking at the plot above, it’s evident that there’s a lot of specialization (scatter, along the x and/or y axes– check out the giraffe/Giraffa and cheetah/Acinonyx outliers, for example). While ungulates seemed to have a better relationship of speed and limb dimensions, their predatory carnivoran relatives did not.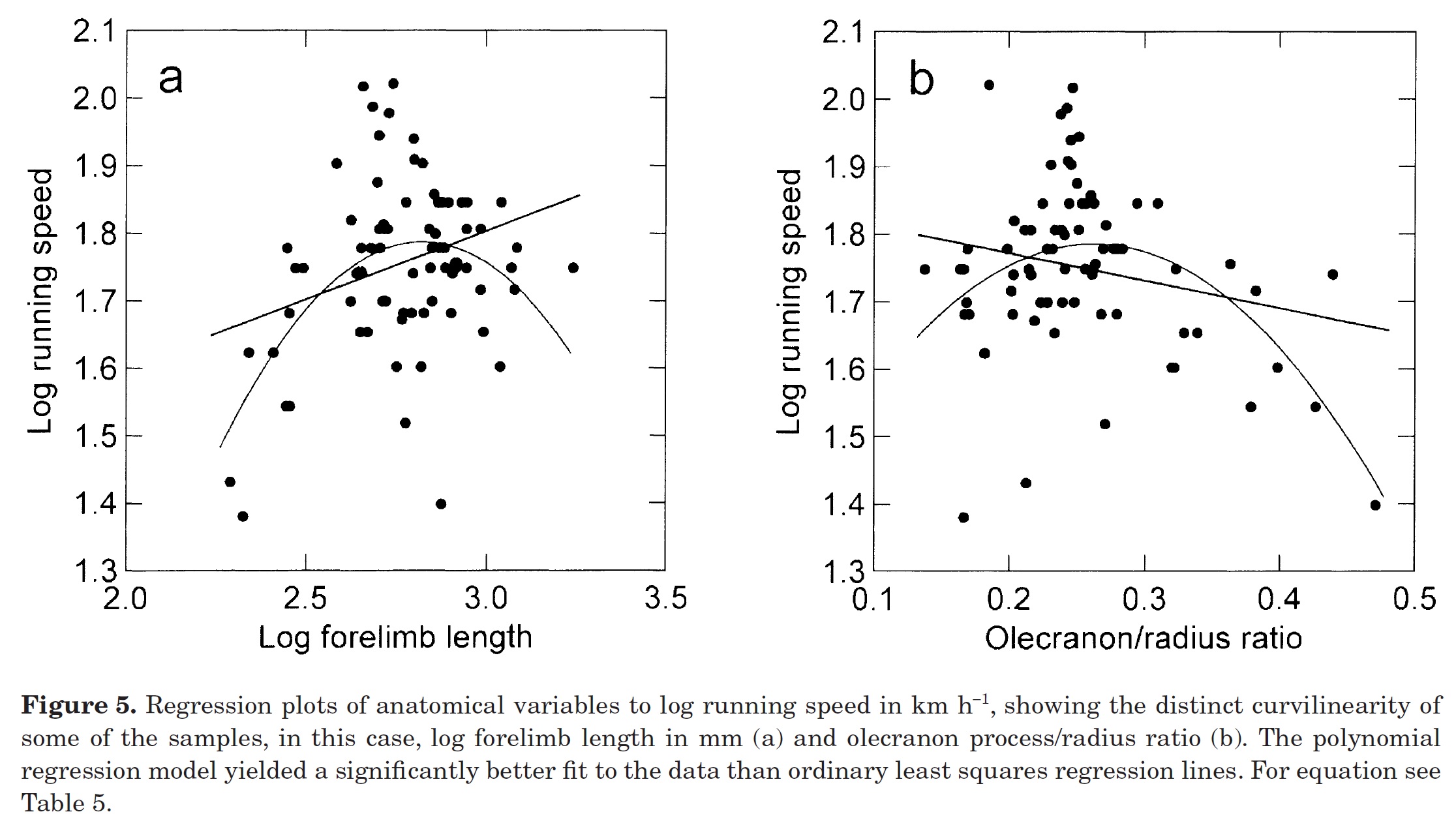
Christiansen was one of two studies in 2002 that looked back on those Garland 1983 data in a new way, and like the 1993 study with Janis considered these data in light of limb lengths too. The plot above delved into how running speed changes with lengths of forelimb bones, again finding appreciable curvilinearity (indirectly supporting the non-linear scaling idea– even at large sizes, relatively longer-legged mammals aren’t faster). The plot on the right side (b) measured the relative length of the olecranon process; the “funny bone” that acts as a lever for support of the elbow joint against gravity. Again, even mammals that have stouter elbow-supporting processes aren’t faster; there’s a “happy medium” of elbow-osity for optimizing running speed (and huge scatter in the data!). Ultimately, this analysis concluded that it wasn’t speed that animal anatomy seemed to be optimizing overall, especially as size increased, but rather energetic cost, although there was a lot of variation in the data and accounting for phylogeny only muddled things up more (as it tends to do).

Iriarte-Diaz was the other 2002 study to tackle the speed-vs-size issue. It focused primarily on whether mammal speeds showed “differential” (i.e. non-linear) scaling with size, as per the polynomial regression in Garland’s 1983 study. It showed that smaller mammals seemed to either get slightly slower with increasing size or else not change maximal speed (depending on detailed methods/data stuff that don’t matter here), whereas bigger mammals exhibited very strong declines of speed with size past a threshold (optimal) body mass.
So, repeated analyses of Garland’s 1983 data (and modifications of those data) at least uphold the fundamental conclusion that big land mammals cannot move quickly, in an absolute sense (meters/sec or kph or mph) — and much more so in a relative sense (e.g. body lengths/second or other normalized metrics). We might then ask why, and my research scrutinizes this issue in terms of the fundamental mechanisms of movement biomechanics and anatomy that might help to explain why, but for brevity I won’t go there in this post. I want to wander elsewhere.
I want to wander back to those data used in the above (and other) studies. All of the studies discuss the quality of the data and bemoan the lack of quality. I’d agree with them that it’s hard to imagine most of the data being consistently off in a biased way that would fundamentally alter their conclusions. But I still worry. We should worry about the data points for the extreme animals- the fastest, slowest, largest and smallest. We should worry about subjectively removing “outliers” such as hippos or cheetahs, as they do change some of the results.
I worry about elephants, for example: my work has shown that they can “run” about 7 meters/second or ~25 kph; not the 35 kph used as data for African elephants (from speedometer-y anecdotal estimates)– ~1.4 times the speed we’ve been able to measure for both species. See this old “blog post” (sort of) for more information on the tortuous history of characterizing elephant speeds and gaits. And are a white rhino and hippo able to run at this same 25 kph speed as the original data in the 1983 study state, or faster/slower? No one has really nicely measured this so we can’t be sure, but I can imagine it being off by a similar 40% or so. On the other hand, if the bigger animals in the dataset are slower than the original data, that actually strengthens the conclusion that bigger animals are slower, so who cares that much, in the grand scheme of things?
We could worry about plenty of other maximal speed data points, and the “average” adult body masses assumed (although I doubt those would change the results as much as the speed errors). Maybe another question is, in doing such broad-scale analyses should we only include data points that have maximal precision (e.g. elephants, horses, cheetahs, greyhounds, humans and a few others)? We’d maybe be able to do a study of 20 or so species. I doubt it would show much that is different if we did, although I expect that sample size and noise would begin to dampen out the signal. See below.
However, a double standard begins to become evident here. In modern biomechanics (and probably the rest of biology/science), there’s a strong emphasis on data quality and technologically precise measurement. Garland’s 1983 study might be hard to get past peer review today (or maybe not). We agonize over single-species studies trying hard to measure animals’ maximal speeds (a very hard thing to be sure of in terms of motivation, but not intractable unless one takes an almost antiscientific/overly cynical view that animals could always be holding back some critical reserve unless they run for their lives– is that reserve 1%, 10% or 100%? Probably closer to the middle, in good studies). We measure multiple animals and many trials, in field and/or lab conditions, with documented video footage at high resolution and frame rate, with GPS tracking or other tools to maximize precision. We take pride in these high standards today. That’s what makes scientists wriggle uncomfortably when we look back at the data in those older maximal speed papers and ponder how few data points are verified, documented, precise and essentially trustworthy.
So should broad studies be working by the same standards as narrow studies? (I’m far, far, far from the first scientist to think about this but it’s interesting for me at least to think about it in this case and others) There is potential tension here between empiricists who want precise data and theoreticians who want to tackle those Big Questions, and that’s a pattern one can see throughout much of science. I sit on the fence myself, doing both approaches. I can think of plenty of similar examples, in “big data” palaeobiology, morphometrics, genomics, physics and so on. Some of those fields have nice databases with quality control over the data; they’ve maybe solved this problem to a large degree. This tiny area of mammalian maximal speeds hasn’t solved it, but how urgent is the need to?
On the flip side, even if the data points have some error of 10-20% or even 40% that error will probably be largely random, not biased toward assuming that bigger or smaller animals are slower than they truly are, or medium-sized animals faster. We still have the reliable cheetah data point (and racehorses, and greyhounds) showing >100 kph (and 70 kph) speeds for ~100 (and ~40, 400ish) kg animals, so there is evidence for a peak of maximal speed (the cheetah outlier, and one might also throw in pronghorn antelope or others that are pretty damn fast but not yet well measured) at medium body size. I expect there would be incremental overall progress if we did improve the data quality, and that would still be nice (comforting!) but it would be a tough, tough slog. Indeed, my team is doing its share of that, already tackling the data point for giraffes this year (stay tuned!). The potential gains are still there, especially for understanding the unique biology of individual species– that noise in the data (or specialization, if you prefer) is interesting!!! We need that kind of work, partly because the big questions, sexy as they are, still depend on having data quality as a foundation, and old questions still need revisiting from time to time as data quality is improved by those in the trenches of gathering it.
My team’s journal club has gone over the Garland paper lately and we’re hitting the others later this summer, but I wanted to throw these thoughts out there on this blog now to see if they generated any fun discussion, or they might introduce others to the science of maximal speeds and what we do/don’t know. One thing we don’t know much about is what kinds of patterns non-mammalian groups exhibit today. Chris Clemente did some great work on this with lizards, finding a pattern similar to the mammal one. I’ve struggled in my work to move toward trying to address similar questions for extinct groups, but there the data quality presents a challenge I find exciting rather than depressing, although I still have to shrug when I see limb lengths or proportions being used as a proxy for speed. We can do better.
So I’d love to hear your thoughts on any of the points here. Maybe some of the old-timers have stories from ye olden days when Garland’s work was originally published; I’d love to hear those, or other points/questions/favourite papers.

