The early, hippo-like mammal Coryphodon. I didn’t know it had a patella but it does. From Yale Peabody Museum.
I’m not shy about my fondness for the patella (kneecap) of tetrapod vertebrates, and neither are the other members of RVC’s “Team Patella”. We’ve had a fun 3+ years studying these neglected bones, and today we’ve published a new study of them. Our attention has turned from our prior studies of bird and lepidosaur kneecaps to mammalian ones. Again, we’ve laid the groundwork for a lot of future work by focusing on (1) basic anatomy and (2) evolutionary history of these sesamoid bones, with a lot of synthesis of existing knowledge from the literature; including development and genetics. This particular paper is a sizeable monograph of the state of play in the perusal of patellae in placental and other synapsids. Here’s what we did and found, focusing mostly on bony (ossified) patellae because that allowed us to bring the fossil record better to bear on the problem.
Reference: Samuels, M., Regnault, S., Hutchinson, J.R. 2016. Evolution of the patellar sesamoid bone in mammals. PeerJ 5:e3103 https://doi.org/10.7717/peerj.3103
Stomach-Churning Rating: 1/10; bones and more bones.
The short version of the story is that mammals evolved bony kneecaps about five times, with marsupials gaining and losing them (maybe multiple times) whereas monotremes (platypus and echidna) and placentals (us and other mammals) didn’t do much once they gained them, and a couple of other fossil groups evolved patellae in apparent isolation.
Evolution of the patella in mammals: broad overview from our paper. Click to zoom in.
The marsupial case is the most fascinating one because they may have started with a fibrocartilaginous “patelloid” and then ossified that, then reduced it to a “patelloid” again and again or maybe even regained it. There needs to be a lot more study of this group to see if the standard tale that “just bandicoots and a few other oddballs have a bony patella” is true for the Metatheria (marsupials + extinct kin). And more study of the development of patellae in this group could help establish whether they truly do “regress” into fibrocartilage when they are “lost” in evolution, or if other, more flexible patterns exist, or even if some of the cases of apparent “loss” of a bony patella are actually instances of delayed ossification that only becomes evident in older adults. Our paper largely punts on these issues because of an absence of sufficient data, but we hope that it is inspiration for others to help carry the flag forward for this mystery.
The higgledy-piggledy evolution of a patella in Metatheria, including marsupials. Click to zoom in.
Some bats, too, do funky things with their kneecaps, analogous to the marsupial “patelloid” pattern, and that chiropteran pattern also is not well understood. Why do some bats such as Pteropus fruit bats “lose” their kneecaps whereas others don’t, and why do some bats and other species (e.g. various primates) seem to have an extra thing near their kneecaps often called a “suprapatella”? Kneecap geeks need to know.
The short-nosed bandicoot (marsupial) Isoodon, showing a nice bony patella as typifies this group. From Yale Peabody Museum.
Otherwise, once mammals evolved kneecaps they tended to keep them unless they lost their hindlimbs entirely (or nearly so). Witness the chunky patellae of early whales such as Pakicetus and join us in wondering why those chunks persisted. The evolutionary persistence of blocky bits of bone in the knees of various aquatic animals, especially foot-propelled diving birds, may help answer why, as the hindlimbs surely still played roles in swimming early in cetacean evolution. Ditto for sea cows (Sirenia) and other groups.
Early whale Ambulocetus, showing hefty kneecaps.
But I’m still left wondering why so many groups of land vertebrates (and aquatic ones, too) never turned parts of their knee extensor tendons into bone. We know a bit about the benefits of doing that, to add leverage to those joints that enables the knee muscles to act with dynamic gearing (becoming more forceful “low gear” or more speedy “high gear” in function). Non-avian (and most early avian/avialan) dinosaurs, crocodiles, turtles, amphibians, early mammal relatives, and almost all other known extinct lineages except for those noted above got by just fine without kneecaps, it seems, even in cases where a naïve biomechanist would expect them to be very handy, such as in giant dinosaurs.
A quoll, Dasyurus, with what is probably a fibrocartilaginous “patelloid”. From Yale Peabody Museum.
However, tendons don’t turn to bone unless the right stresses and strains are placed upon them, so maybe kneecaps are a “spandrel” or “exaptation” of sorts, to abuse Gould’s ghost, whose adaptive importance is overemphasized. Maybe that adaptive myopia overshadows a deeper ontogenetic story, of how tissues respond to their history of mechanical loading environment. It has been speculated that maybe (non-marsupial) mammals have broadly “genetically assimilated” their kneecaps, fixing them into semi-permanence in their genetic-developmental programmes, whereas in contrast the few studies of birds indicate more responsiveness and thus less assimilation/fixation. That “evo-devo-mechanics” story is what now fascinates me most and we’ve poked at this question a bit now, with some updates to come- watch this space! Regardless, whether an animal has a bony vs. more squishy soft tissue patella must have consequences for how the knee joint and muscles are loaded, so this kind of question is important.
Giant marsupial Diprotodon (at NHM London); to my knowledge, not known to have had kneecaps- why?
In the meantime, enjoy our latest contribution if it interests you. This paper came about when first author Dr. Mark Samuels emailed me in 2012, saying he’d read some of my old papers on the avian musculoskeletal system and was curious about the evolution of patellae in various lineages. Unlike many doctors and vets I’ve run into, he was deeply fascinated by the evolutionary and fossil components of patellae and how those relate to development, genetics and disorders of patellae. We got talking, found that we were kindred kneecap-spirits, and a collaboration serendipitously spun off from that, soon adding in Sophie. It was a blast!
It has been almost three months since my last post here, and things have fallen quiet on our sister blog Anatomy to You, too. I thought it was time for an update, which is mostly a summary of stuff we’ve been doing on my team, but also featuring some interesting images if you stick around. The relative silence here has partly been due to me giving myself some nice holiday time w/family in L.A., then having surgery to fix my right shoulder, then recovering from that and some complications (still underway, but the fact that I am doing this post is itself evidence of recovery).
Stomach-Churning Rating: 4/10; semi-gruesome x-rays of me and hippo bits at the end, but just bones really.
X-ray of my right shoulder from frontal view, unlabelled
Labelled x-ray
So my priorities shifted to those things and to what work priorities most badly needed my limited energy and time. I’ve also felt that, especially since my health has had its two-year rough patch, this blog has been quieter and less interactive than it used to be, but that is the nature of things and maybe part of a broader trend in blogs, too. My creative juices in terms of social media just haven’t been at their ~2011-2014 levels but much is out of my control, and I am hopeful that time will reverse that trend. Enough about all this. I want to talk about science for the rest of this post.
My team, and collaborators as well, have published six recent studies that are very relevant to this blog’s theme- how about we run through them quickly? OK then.
Panagiotopoulou, O., Pataky, T.C., Day, M., Hensman, M.C., Hensman, S., Hutchinson, J.R., Clemente, C.J. 2016. Foot pressure distributions during walking in African elephants (Loxodonta africana). Royal Society Open Science 3: 160203.
Our Australian collaborators got five African elephants together in Limpopo, South Africa and walked them over pressure-measuring mats, mimicking our 2012 study of Asian elephants. While sample sizes were too limited to say much statistically, in qualitatively descriptive terms we didn’t find striking differences between the two species’ foot pressure patterns. I particularly like how the centre of pressure of each foot (i.e. abstracting all regional pressures down to one mean point over time) followed essentially the same pattern in our African and Asian elephants, with a variable heelstrike concentration that then moved forward throughout the step, and finally moved toward the outer (3rd-5th; especially 3rd) toes as the foot pushed off the ground, as below.
African elephant foot COP traces vs. time in red; Asian elephant in orange-yellow. Left and right forefeet above; hindfeet below.
Gradually, this work is moving the field toward better ability to use similar techniques to compare elephant foot mechanics among species, individuals, or over time– especially with the potential of using this method (popular in human clinical gait labs) to monitor foot (and broader musculoskeletal) health in elephants. I am hopeful that a difference can be made, and the basic science we’ve done to date will be a foundation for that.
Panagiotopoulou, O., Rankin, J.W., Gatesy, S.M., Hutchinson, J.R. 2016. A preliminary case study of the effect of shoe-wearing on the biomechanics of a horse’s foot. PeerJ 4: e2164.
Finally, about six years after we collected some very challenging experimental data in our lab, we’ve published our first study on them. It’s a methodological study of one horse, not something one can hang any hats on statistically, but we threw the “kitchen sink” of biomechanics at that horse (harmlessly!) by combining standard in vivo forceplate analysis with “XROMM” (scientific rotoscopy with biplanar fluoroscopy or “x-ray video”) to conduct dynamic analysis of forefoot joint motions and forces (with and without horseshoes on the horse), and then to use these data as input values for finite element analysis (FEA) of estimated skeletal stresses and strains. This method sets the stage for some even more ambitious comparative studies that we’re finishing up now. And it is not in short supply of cool biomechanical, anatomical images so here ya go:
Above: The toe bones (phalanges) of our horse’s forefoot in dorsal (cranial/front) view, from our FEA results, with hot colours showing higher relative stresses- in this case, hinting (but not demonstrating statistically) that wearing horseshoes might increase stresses in some regions on the feet. But more convincingly, showing that we have a scientific workflow set up to do these kinds of biomechanical calculations from experiments to computer models and simulations, which was not trivial.
And a cool XROMM video of our horse’s foot motions:
Bates, K.T., Mannion, P.D., Falkingham, P.L., Brusatte, S.L., Hutchinson, J.R., Otero, A., Sellers, W.I., Sullivan, C., Stevens, K.A., Allen, V. 2016. Temporal and phylogenetic evolution of the sauropod dinosaur body plan. Royal Society Open Science 3: 150636.
I had the good fortune of joining a big international team of sauropod experts to look at how the shapes and sizes of body segments in sauropods evolved and how those influenced the position of the body’s centre of mass, similar to what we did earlier with theropod dinosaurs. My role was minor but I enjoyed the study (despite a rough ride with some early reviews) and the final product is one cool paper in my opinion. Here’s an example:
The (embiggenable-by-clicking) plot shows that early dinosaurs shifted their centre of mass (COM) backwards (maybe related to becoming bipedal?) and then sauropods shifted the COM forwards again (i.e. toward their forelimbs and heads) throughout much of their evolution. This was related to quadrupedalism and giant size as well as to evolving a longer neck; which makes sense (and I’m glad the data broadly supported it). But it is also a reminder that not all sauropods moved in the same ways- the change of COM would have required changes in how they moved. There was also plenty of methodological nuance here to cover all the uncertainties but for that, see the 17 page paper and 86 pages of supplementary material…
Randau, M., Goswami, A., Hutchinson, J.R., Cuff, A.R., Pierce, S.E. 2016. Cryptic complexity in felid vertebral evolution: shape differentiation and allometry of the axial skeleton. Zoological Journal of the Linnean Society 178:183-202.
Back in 2011, Stephanie Pierce, Jenny Clack and I tried some simple linear morphometrics (shape analysis) to see how pinniped (seal, walrus, etc) mammals changed their vertebral morphology with size and regionally across their backbones. Now in this new study, with “Team Cat” assembled, PhD student Marcela Randau collected her own big dataset for felid (cat) backbones and applied some even fancier techniques to see how cat spines change their shape and size. We found that overall the vertebrae tended to get relatively more robust in larger cats, helping to resist gravity and other forces, and that cats with different ecologies across the arboreal-to-terrestrial spectrum also changed their (lumbar) vertebral shape differently. Now Marcela’s work is diving even deeper into these issues; stay tuned…
Example measurements taken on felid vertebrae, from the neck (A-F) to the lumbar region (G-J), using a cheetah skeleton.
Charles, J.P., Cappellari, O., Spence, A.J., Hutchinson, J.R., Wells, D.J. 2016. Musculoskeletal geometry, muscle architecture and functional specialisations of the mouse hindlimb. PLOS One 11(4): e0147669.
RVC PhD student James Charles measured the heck out of some normal mice, dissecting their hindlimb muscle anatomy, and using microCT scans produced some gorgeous images of that anatomy too. In the process, he also quantified how each muscle is differently specialized for the ability to produce large forces, rapid contractions or fine control. Those data were essential for the next study, where we got more computational!
Charles, J.P., Cappellari, O., Spence, A.J., Wells, D.J., Hutchinson, J.R. 2016. Muscle moment arms and sensitivity analysis of a mouse hindlimb musculoskeletal model. Journal of Anatomy 229:514–535.
James wrangled together a lovely musculoskeletal model of our representative mouse subject’s hindlimb in the SIMM software that my team uses for these kinds of biomechanical analyses. As we normally do as a first step, we used the model to estimate things that are hard to measure directly, such as the leverages (moment arms) of each individual muscle and how those change with limb posture (which can produce variable gearing of muscles around joints). James has his PhD viva (defense) next week so good luck James!
The horse and mouse papers are exemplars of what my team now does routinely. For about 15 years now, I’ve been building my team toward doing these kinds of fusion of data from anatomy, experimental biomechanics, musculoskeletal and other models, and simulation (i.e. estimating unmeasurable parameters by telling a model to execute a behaviour with a given set of criteria to try to perform well). Big thanks go to collaborator Jeff Rankin for helping us move that along lately. Our ostrich study from earlier this year shows the best example we’ve done yet with this, but there’s plenty more to come.
I am incredibly excited that, now that my team has the tools and expertise built up to do what I’ve long wanted to do, we can finally deliver the goods on the aspirations I had back when I was a postdoc, and which we have put enormous effort into pushing forward since then. In addition to new analyses of horses and mice and other animals, we’ll be trying to push the envelope more with how well we can apply similar methods to extinct animals, which brings new challenges– and evolutionary questions that get me very, very fired up.
Here we are, then; time has brought some changes to my life and work and it will continue to as we pass this juncture. I suspect I’ll look back on 2016 and see it as transformative, but it hasn’t been an easy year either, to say the least. “Draining” is the word that leaps to mind right now—but also “Focused” applies, because I had to try to be that, and sometimes succeeded. I’ve certainly benefited a lot at work from having some talented staff, students and other collaborators cranking out cool papers with me.
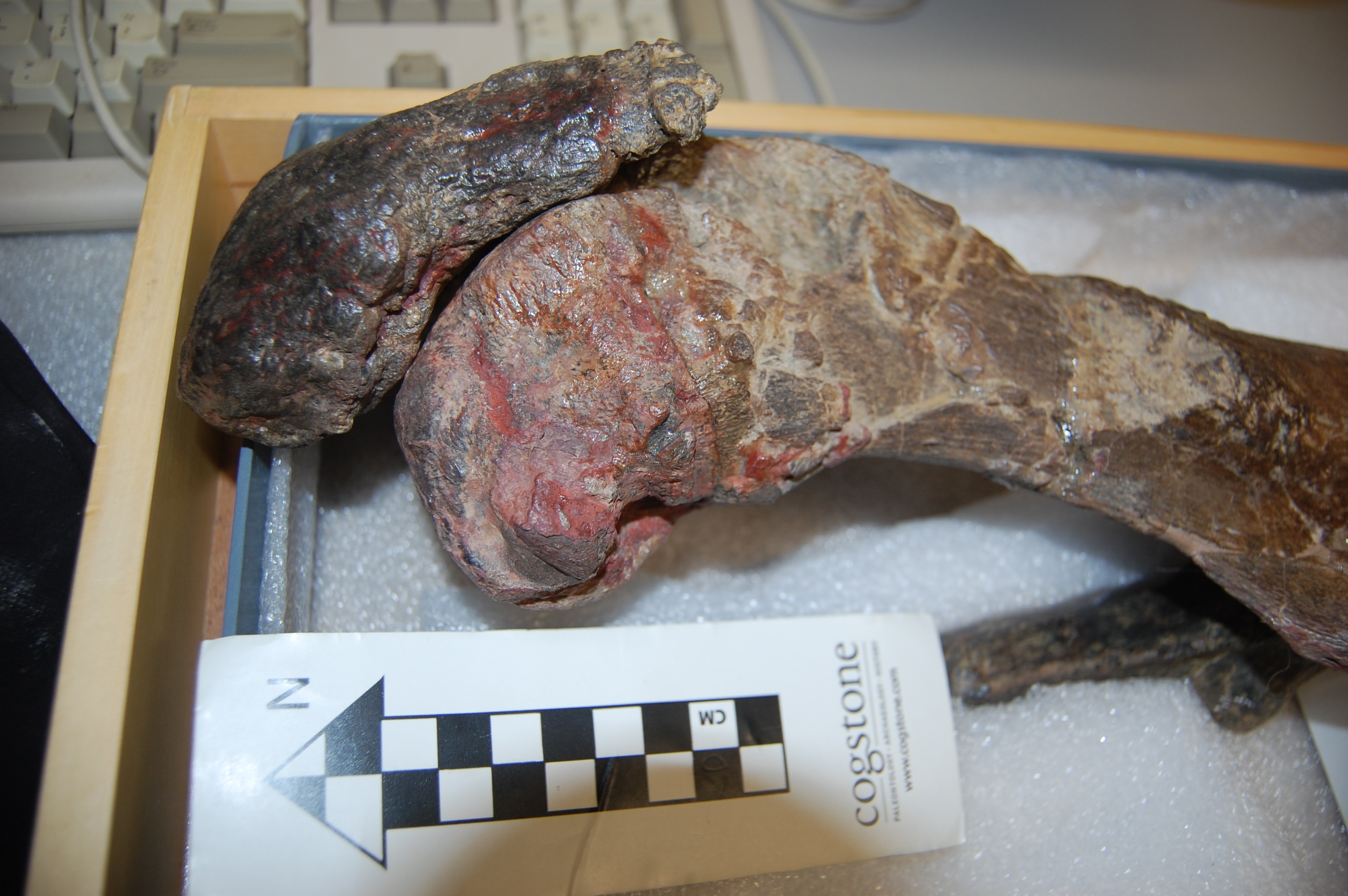
I still have time to do other things, too. Once in a while, a cool critter manifests in The Freezers. Check out a hippo foot from a CT scan! It’s not my best scan ever (noisy data) but it shows the anatomy fairly well, and some odd pathologies such as tiny floating lumps of mineralized soft tissue here and there. Lots to puzzle over.
Who needs “Ice Road Truckers” when you have the “John’s Freezer” team on the road with fossils, amphibians, felids and 3D phenotype fun? No one, that’s who. We’re rocking the Cheltenham Science Festival for our first time (as a group), and pulling out all the stops by presenting two events! Here’s the skinny on them, with updates as the week proceeds.
Stomach-Churning Rating: 2/10 for now (just bones), but it could change once the cheetah dissection is under way… 8/10 bloody cheetah bits but only at the end (updated)
Right now, Lauren Sumner-Rooney (of “Anatomy To You” and other fame) is on-site with a rotating team of others from our lab, in the “Free Activity Tents” area of the Imperial Gardens/Square, inside a marquee where we’ll be showing off our NERC-funded tetrapod research all week. This “First Steps” event features not only our past and present work with Jenny Clack, Stephanie Pierce, Julia Molnar and others on Ichthyostega & its “fishapod” mates, but also our “scampering salamanders” research in Spain, Germany and England. I’ve blogged a lot about all that, and won’t repeat it here, but you can see a 3D-printed Ichthyostega skeleton, view the skeleton in a virtual reality 3D environment, see related specimens and engage in kid-friendly activities, and talk to our team about this and other related research.
Ichthyostega 3D printed backbone is born!
The central themes of that event are how bone structure relates to function and how we can use such information, along with experimental measurements and computer models of real salamanders, to reconstruct how extinct animals might have behaved as well as how swimming animals became walking ones. How did fins transform into limbs and what did that mean for how vertebrates made the evolutionary transition onto land? If you know my team’s work, that encapsulates our general approach to many other problems in evolutionary biomechanics (e.g. how did avian bipedalism evolve?). Added benefits are that you too can explore this theme in a hands-on way, and you can talk with us about it in person. That continues all week (i.e. until Saturday evening); I’ll be around from Thursday afternoon onwards, too. Kids of all ages are welcome!
Ichthyostega 3D print taking shape!
Then, on Saturday for our second free event we join forces with Ben Garrod (master of primate evolution, the secrets of bones, and “Attenborough and the Giant Dinosaur”) and RVC’s forensic pathologist Alexander Stoll as well as Sophie Regnault (“sesamoid street” PhD student w/me). As the “Large Animal Dissection” title hints, it’s not the right kind of gig to bring small kids to. There will be blood and stuff— we’ll be dissecting a cheetah together from 10am-4pm. This will involve walking through all the major organ systems, giving evolutionary anecdotes, and plenty more, with an aim to understand how the magnificent adaptations of cheetahs evolved—but also to investigate what problem(s) this animal faced that led to its sad demise. By the day’s end, there will just be a skeleton left. Get a front row seat early for this event, which serendipitously ties into “Team Cat”’s Leverhulme Trust-funded research project (we wanted a big animal and it just happened to be a cheetah; I had hoped for a giant croc or a shark or something but can’t complain!).
Ichthyostega 3D print is ready!
If you miss these events, please do cry bitter tears of regret. But don’t despair, there will be another “big cat dissection” in the London area in ~November (watch here for details), and plenty more fossil tetrapod stuff to come, plus a LOT more dinosaurs on the horizon!
Guess the bones! (photo by Zoe Self)
And please come back to this blog post for more pics and stories as the week carries on… For hashtag afficionados, you can follow the fun on Twitter etc. at #firststepsCSF16. What a world we live in!
Update 1: While you’re here, check out our Youtube playlists of tetrapod-related videos:
Update 2: Photos of our main stand (about tetrapod evolution)
Our poster/banner display looks nice.
Our tent brings in some punters.
Our bones excite people young and old, sighted and blind.
Fun with stickers and lab t-shirts.
And…
Update 3: Cheetah meat & greet
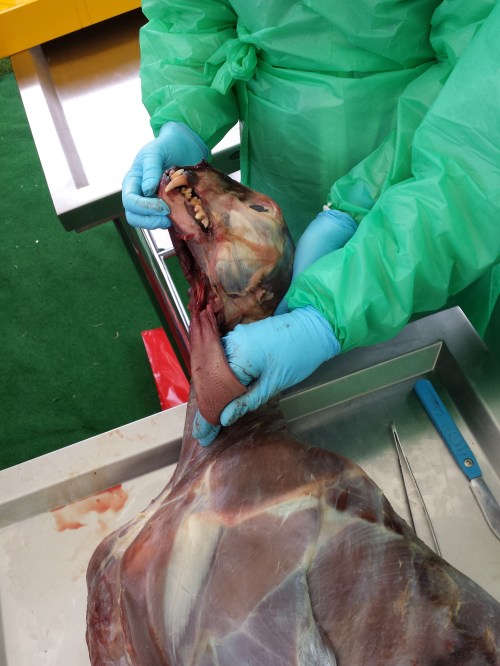
Ben, Alex, Sophie and I tackled the cheetah dissection today and it went GREAT! Much better than I’d optimistically expected. Rain didn’t scare the crowds off and neither did the gore, which there was some of (gelatinous spinal cords, lumpy tumors and at least one flying tiny bit of cheetah flesh that landed on a good-natured audience member!). Photos will tell the tale:
Peek-a-boo!
Sophie and Alex help us get set up in our tent.
Our initial rough schedule- although we ended up improvising more after lunch.
Dissectors assemble!
The beast revealed. It was skinned by the museum that loaned it to us.
Alex showing his talent: removing the viscera in one piece from end to end, starting with the tongue.
Impressive finding of a surgical fixture (plate and wires) on the tibia, which had been used to hold the shattered bone back together long enough for it to heal. Added to the kidney disease and liver-spleen-lung cancer, this cheetah was in the sorriest shape of any cadaver I’ve seen yet.
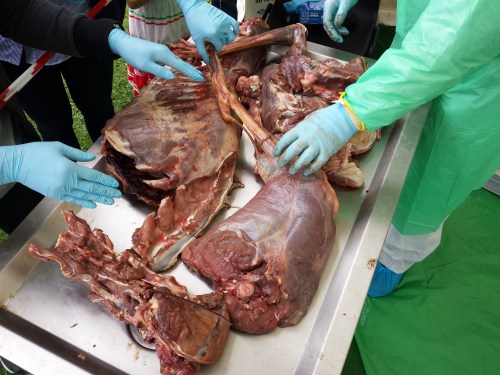
Cheetah coming to pieces: (from bottom) lumbar/pelvic region, hindlimb, thorax, forelimb and other bits.
Dr Adam Rutherford, an eye expert, did a nice dissection of the cheetah’s eye, here showing the tapetum lucidum (reflective membrane), which shows up as light blue colour. Its small size befits the not-very-nocturnal habits of cheetahs.
The lens of the cheetah’s eye. Now cloudy because of dehydration and crystalization, but still fascinating to see.
Seeking adaptations for running and swimming in the vertebral columns of ancient crocs
A guest post by Dr. Julia Molnar, Howard University, USA (this comes from Julia’s PhD research at RVC with John & colleagues)
Recently, John and I with colleagues Stephanie Pierce, Bhart-Anjan Bhullar, and Alan Turner described morphological and functional changes in the vertebral column with increasing aquatic adaptation in crocodylomorphs (Royal Society Open Science, doi 10.1098/rsos.150439). Our results shed light upon key aspects of the evolutionary history of these under-appreciated archosaurs.
Stomach-Churning Rating: 5/10; a juicy croc torso in one small photo but that’s all.
Phylogenetic relationships of the three crocodylomorph groups in the study and our functional hypotheses about their vertebrae. * Image credits: Hesperosuchus by Smokeybjb, Suchodus by Dmitry Bogdanov (vectorized by T. Michael Keesey) http://creativecommons.org/licenses/by-sa/3.0
As fascinating as modern crocodiles might be, in many ways they are overshadowed by their extinct, Mesozoic cousins and ancestors. The Triassic, Jurassic, and early Cretaceous periods saw the small, fast, hyper-carnivorous “sphenosuchians,” the giant, flippered marine thalattosuchians, and various oddballs like the duck-billed Anatosuchus and the aptly named Armadillosuchus. As palaeontologists/biomechanists, we looked at this wide variety of ecological specializations in those species, the Crocodylomorpha, and wanted to know, how did they do it?
Of course, we weren’t the first scientists to wonder about the locomotion of crocodylomorphs, but we did have some new tools in our toolbox; specifically, a couple of micro-CT scanners and some sophisticated imaging software. We took CT and micro-CT scans of five fossil crocodylomorphs: two presumably terrestrial early crocodylomorphs (Terrestrisuchus and Protosuchus), three aquatic thalattosuchians (Pelagosaurus, Steneosaurus, and Metriorhynchus) and a semi-aquatic modern crocodile (Crocodylus niloticus). Since we’re still stuck on vertebrae (see, e.g., here; and also here), we digitally separated out the vertebrae to make 3D models of individual joints and took measurements from each vertebra. Finally, we manipulated the virtual joint models to find out how far they could move before the bones bumped into each other or the joints came apart (osteological range of motion, or RoM).
Our methods: get fossil (NHMUK), scan fossil, make virtual fossil and play with it.
Above: Video of a single virtual inter-vertebral joint from the trunk of Pelagosaurus typus (NHMUK) showing maximum osteological range of motion in the lateral direction (video). Note the very un-modern-croc-like flat surfaces of the vertebral bodies! (modern crocs have a ball-and-socket spinal joint with the socket on the front end)
While this was a lot of fun, what we really wanted to find out was whether, as crocodylomorphs became specialized for different types of locomotion, the shapes of their vertebrae changed similarly to those of mammalian lineages. For example, many terrestrial mammals have a lumbar region that is very flexible dorsoventrally to allow up-and-down movements during bounding and galloping. Did fast-running crocodylomorphs have similar dorsoventral flexibility? And did fast-swimming aquatic crocodylomorphs evolve a stiffer vertebral column like that of whales and dolphins?
Above: Video of how we modelled and took measurements from the early crocodylomorph Terrestrisuchus gracilis (NHMUK).
Our first results were puzzling. The Nile croc had greater RoM in side-to-side motions, which makes sense because crocodiles mostly use more sprawling postures and are semi-aquatic, using quite a bit of side-to-side motions in life. The part that didn’t make sense was that we found pretty much the same thing in all of the fossil crocodylomorphs, including the presumably very terrestrial Terrestrisuchus and Protosuchus. With their long limbs and hinge-like joints, these two are unlikely to have been sprawlers or swimmers!
So we started looking for other parts of the croc that might affect RoM. The obvious candidate was osteoderms, the bony scales that cover the back. We went back to John’s Freezer and got out a nice frozen crocodile to measure the stiffness of its trunk and found that, sure enough, it was a lot stiffer and less mobile without the osteoderms. If the fairly flexible arrangement of osteoderms in crocodiles had this effect on stiffness, it seemed likely that (as previous authors have suggested; Eberhard Frey and Steve Salisbury being foremost amongst them) the rigid, interlocking osteoderms running from head to tail in early crocodylomorphs would really have put the brakes on their ability to move their trunk in certain ways.
Testing the stiffness of (Nile) crocodile trunks to learn the effects of osteoderms, skin, muscles, and ribs. We hung metric weights from the middle of the trunk and measured how much it flexed (Ɵ), then removed bits and repeated. Click to em-croccen.
Another cool thing we found was new evidence of convergent evolution to aquatic lifestyles in the spines of thalattosuchians. The more basal thalattosuchians, thought to have been near-shore predators, had stiffness and RoM patterns similar to Crocodylus. But Metriorhynchus, which probably was very good at chasing down fast fish in the open ocean, seems to have had greater stiffness. (The stiffness estimates come from morphometrics and are based on modern crocodiles; see here again, or just read the paper already!) A stiff vertebral column can be useful for a swimmer because it increases the body’s natural frequency of oscillation, and faster oscillation means faster swimming (think tuna, not eel). The same thing seems to have happened in other secondarily aquatic vertebrate lineages such as whales, ichthyosaurs, and mosasaurs.
So, our results were a mixed bag of adaptations particular to crocs and ones that seem like general vertebrate swimming specializations. Crocodylomorphs are important because they are the only group of large vertebrates other than mammals that has secondarily aquatic members and has living members with a reasonably similar body plan, allowing us to test hypotheses in ways that would arguably be impossible for, say, non-avian dinosaurs and birds. The take-home message: crocodylomorphs A) are awesome, and B) can teach us a lot about how vertebrates adapt to different modes of life.
Well, that was an eventful week for me, although today’s post will focus on one event: the debut of the film Jurassic World. Briefly though, the awesome “T. rex Autopsy” documentary debuted (I was going to post more about it but all I’d have left to say is that I was very pleased with the result), I also showed up briefly in “Top 10 Biggest Beasts Ever” talking about the giant rhinocerotoid Paraceratherium and the stresses on its feet, our paper on ostrich musculoskeletal modelling was published (more in a future post) after ~12 years of me diddling around with it, and much more happened. Then to cap it all off, very shortly after I hit the “publish” button on my last post, I had four tonic clonic seizures in a row and spent a hazy night in the hospital, then the past week recovering from the damage. Nothing like another near-death (no exaggeration there, I’m afraid) experience to cap off an exciting week. But strangely, what I feel more interested in talking about is, like I said, Jurassic World, but this is not a review, as you will see here.
Stomach-Churning Rating: 0/10; just SPOILERS if you haven’t seen the film yet!
I guess I have to give a brief review of the film and say that I was entertained, to a degree, but it was not a great piece of film-making. It was a far cry from the original but then so were the sequels, and maybe it was better than them. The mosa-star was the most novel, memorable bit. I didn’t care for the Indominus villain, but then when you bring genetic engineering into a film like this, you’ve basically thrown out the rulebook and can make your dinosaurs as magical as you want; we’re already in “X-Men” territory here and almost in “Pacific Rim”-land.
Chris Pratt has signed on for at least another sequel to Jurassic World and the ending of the film already started that ball rolling. So I find it fun to speculate wildly, and certainly incorrectly, on what the sequel might do. What does the Jurassic future have in store?
First of all, who survived to re-appear in the next film? We’re left with the Bryce Dallas Howard character, who probably will return with Pratt to further develop their rather uninteresting social/romantic dynamic, rather than start afresh with someone else. The kids of course survived, as always, and as always they won’t return, as that’s not interesting and they didn’t have much to do except scream and (highly implausibly) hotwire an old car. Much as I’d like the parents from my hometown of Madison, WI to return, they won’t either for the same reasons. But we really only need Pratt and his high-heeled sweetie for the next film. Everyone else memorable(?) seems to have died, although it would be wonderful to bring Goldblum back for some smarmy wit (please!).
Second of all, the next film can’t be set in Jurassic World. There’s not much left to do there (JW already spent much of its time hearkening back to JP), and there’s no way the park would re-open. We need something new. I think by now we’re (very) tired of characters running around islands full of dinosaurs and the Blackfish parallel was milked dry in the latest movie. We need to spend a film with the dinosaurs amongst humanity (as Lost World briefly did), and much as I’d love to see the crazy drug lord/kidnapping plot happen, it won’t. But JW did set one thing up that has to happen now in its sequel: the paramilitary role of engineered, trained dinosaurs. We now know they can sort of train their dinosaurs and they can forge them to be anything they want to in terms of geno/phenotypes. They’ll learn from some mistakes of JW and engineer (or already did by the end of JW, at some remote site) some more compliant, deadlier animals, having largely given up on the public exhibition angle. The naked raptors and T. rex probably have to re-appear (sigh), but enough already of the giant uber-theropods like Spinosaurus and Indominus. The latter was already enough of a reprise of the former (plus psychic talents and chameleon powers etc.). Something truly novel is needed.
Unless they engineer a hyper-aggressive, intelligent sauropod or ceratopsian, which would admittedly be neat, I have this prediction (which is probably wrong but hey!): they have shown they can hybridize anything. There must be fewer and fewer “normal” (1990s…) dinosaurs now in the JW universe. So the next big step, which someone in the JW universe surely would do, is to hybridize dinosaurs and humans. Maybe some raptor-human hybrids, maybe also saving a tyranno-human hybrid for a surprise late appearance. But this is the sensible next step because it allows them to play with the (tired) Frankenstein monster trope but also touch on the hot topic of human cloning and human GM.
And by unleashing dino-human hybrids, or at least some freaky clicker-trained and engineered super-dinos, they could also explore the military theme, which the JW universe still hasn’t delved into much. What if those hyper-smart, deadly hybrid dinos, led by Pratt and Howard’s expert training, were used to combat an ISIS-analogue terrorist threat? Dino-Avengers in the badlands of Afghanistan or Iraq? Too predictable perhaps, but that’s a film that the public will want to see. Yeah there’s plenty of stupid there, but there’s no turning back– each film ups the ante, as JW ironically reminds us several times. We’re already in firmly in stupid-land, and the science has largely advanced to the point of magic. My idea is too uncomfortably close to the abandoned John Sayles plot, true. Darn. And (groan) kids have to be involved in some way to make it a family film so it rakes in the $$$ again, so either they get caught in the middle of the paramilitary mess or they are the ones that have to be saved… or the hybrid dinos are cute-ish kids themselves that Pratt and Howard must manage… (shades of Teenage Mutant Ninja Turtles/Kick-Ass?) I’ve found that more fun to think about than dwelling on the flaws of the movie, which is frankly too easy.
(Another good theme that Vivian Allen suggested to me would be climate change and invasive species—i.e. planet warms, dinos are already loose and go feral in waterlogged Central America, ecological disaster is looming and something must be done to round up the dinos… could work in some other bits like ecotourists or drug runners?)
That’s as far as my wildly speculative ruminating has taken me, but I wanted to turn it over to you, Freezerinos. If you were to make the next film (will it be “Jurassic World 2”? “Jurassic Army”? or as I’ve proffered in the post’s title, “Jurassic Future”?), what would it be (A) in your ideal world where you call all the shots (yes, lots of colourful feathery dinos, I know), vs. (B) in a more likely (less daring, more Hollywood) reality, along the lines of what I’ve tried to do here? (but I surely will be wrong, although we’ll see in 2-5 years!)
Ambitious experimental and morphological studies of a modern fish show how a flexible phenotype may have helped early “fishapods” to make the long transition from finned aquatic animals into tetrapods able to walk on land.
Stomach-Churning Rating: 1/10. Cute fish. Good science. Happy stomachs!
Photo by Antoine Morin, showing Polypterus on land.
Napoleon Bonaparte’s military excursions into Egypt in 1798-1799 led a young French naturalist, Ètienne Geoffroy Saint-Hilaire, to cross paths with a strange fish that had paired lungs and could “walk” across land on its stubby, lobelike fins. In 1802, he dubbed this fish “Polyptère bichir”1, today known as the Nile bichir, Polypterus bichir La Cepède 1803. The bichir’s mélange of primitive and advanced traits helped to catapult Geoffroy into scholarly conflict with the reigning naturalist Georges Cuvier back in France and to establish Ètienne as a leading anatomist, embryologist and early evolutionary researcher of repute even today2. Now, on their own excursion under the very “evo-devo” flag that the discoverer of Polypterus helped raise, Canadian scientists Standen et al.3 suggest how the remarkable plasticity of the skeleton of Polypterus (the smaller west African relative of P. bichir, P. senegalus or “Cuvier’s bichir”) reveals a key part of the mechanism that might have facilitated the gradual transition from water to land and thus from “fishapods” to tetrapods (four-limbed vertebrates).
In a bold experiment, the authors raised 149 young bichirs on land and in water for eight months, then studied how they moved on land vs. in water, and also how the ultimate shape of the skeletal elements of the paired front fin bases differed between the land- and water-raised bichirs. Standen et al.3 discovered that both the form and function of the fins’ foundations transformed to better satisfy the constraints of moving on land. Land-acclimated bichirs took faster steps on land, their fins slipped across the substrate less, they held their fins closer to their body, their noses stayed more aloft and their tails undulated less, with less variable motions overall—behaviours that the authors had predicted should appear to enhance walking abilities on land. In turn, the bones of the neck and shoulder region altered their shape to produce a more mobile fin base with greater independence of fin from neck motion, along with improved bracing of the ventral “collarbone” region. These environmentally-induced traits should have fostered the locomotor changes observed in “terrestrialized” fish and aided the animals in resisting gravity, and they represent a common biological phenomenon termed developmental plasticity4,5. Interestingly, the land-reared fish could still swim about as well as the wholly aquatic cohort, so there was not a clear trade-off between being a good swimmer and a good walker, which is surprising.
Considered alone, the developmental plasticity of bichir form and function shows how impressive these amphibious fish are. But Standen et al.’s study3 ventured further, to apply the lessons learned from bichir ontogeny to a phylogenetic context and macroevolutionary question. The phenotypic plasticity during bichir development, they infer, could have been harnessed during the evolutionary transformation of fins for swimming into limbs for walking, in the “fishapod” ancestors of tetrapods. Indeed, bichirs are close to the base of the family tree of fishes6, and other living relatives of tetrapods have reduced or lost their fins (lungfishes) or adapted to strange deep-sea swimming lifestyles, never walking on land (coelacanths). Thus perhaps bichirs and the “fishapod” lineage share what Geoffroy would have called “unity of type”, today termed homology, of their developmental plasticity in response to a land environment. Surveying the fossil record of early “fishapods” and tetrapods, Standen et al.3 found that the macroevolutionary changes of neck and shoulder anatomy in these gradually more land-adapted animals parallel those they observed in terrestrialized Polypterus, providing ancillary support for their hypothesis.
A further test of the application of Polypterus’s plasticity to fossil tetrapods is naturally difficult. However, the “fishapod” lineage has some exceptional examples of fossil preservation. With sufficient sample sizes (e.g. fossil beds that reveal growth series, such as the Late Devonian Miguasha site in Canada7) and palaeoenvironmental gradients in fish or tetrapods, one could imagine performing a rigorous indirect test. Even small samples could be helpful– for example, the early tetrapod Ichthyostega exhibits some developmental changes in its forelimb suggesting that it became more terrestrial as it grew, whereas the related Acanthostega does not evidence such changes8— this hints at some developmental plasticity in the former animal.
During the Devonian period (~360-420 million years ago), were the “fishapod” ancestors of tetrapods floundering about on land now and then, gradually shifting from anatomy and behaviours that were more developmentally plastic (as in bichirs) to ones that were more canalized into the terrestrialized forms and functions that more land-adapted tetrapods retained? An attractive possibility is that the developmental plasticity could have led to fixation (reduction of plasticity), an evolutionary phenomenon called genetic assimilation, which another intellectual descendant of Geoffroy, Conrad Hal Waddington, promoted from the 1950s onwards9, a concept that now enjoys numerous cases of empirical support10 that this one may eventually join.
The nature of the genetic and developmental mechanism that bichirs use to achieve the observed developmental plasticity is still unclear. If it has a high enough degree of heritability, then it could be selected for in cross-generational experiments with bichirs. With sufficient time and luck raising these unusual fish, the hypothesis that their plastic response to a terrestrial environment can become genetically assimilated could be directly tested. This study could thus become an epic exemplar of how genetic assimilation can contribute not only to microevolutionary change but also to major macroevolutionary events, as was presciently suggested in a seminal review of developmental plasticity4.
This genetic assimilation is the Polypterus study’s reasonable speculation, and one that Geoffroy likely would have applauded, all the more for involving his beloved bichirs. Much as Napoleon’s landfall in Egypt was not a lasting success, bichirs never left wholly terrestrial descendants despite their malleable locomotor system. But the same type of plastic developmental mechanism that bichirs use today to make tentative, floppy incursions of the terrestrial realm might have been harnessed by our own “fishapod” forebears, leaving a far more revolutionary dynasty upon the Earth.
References
Geoffroy, E. (1802). Histoire naturelle et description anatomique d’un nouveau genre de poisson du Nil, nommé polyptère. Annales du Muséum d’Histoire Naturelle 1:57-68.
Le Guyader, H., & Grene, M. (2004) Geoffroy Saint-Hilaire: A Visionary Naturalist. Univ. Chicago Press.
Standen, E. M., Du, T. Y., & Larsson, H. C. E. (2014). Developmental plasticity and the origin of tetrapods. Nature, published online.
West-Eberhard, M. J. (1989). Phenotypic plasticity and the origins of diversity. Annual Review of Ecology and Systematics 20:249-278.
Pigliucci, M., Murren, C. J., & Schlichting, C. D. (2006). Phenotypic plasticity and evolution by genetic assimilation. Journal of Experimental Biology 209(12):2362-2367.
Near, T. J., Dornburg, A., Tokita, M., Suzuki, D., Brandley, M. C., & Friedman, M. (2014). Boom and bust: ancient and recent diversification in bichirs (Polypteridae: Actinopterygii), a relictual lineage of ray‐finned fishes. Evolution68:1014-1026.
Cloutier, R. (2013). Great Canadian Lagerstätten 4. The Devonian Miguasha Biota (Québec): UNESCO World Heritage Site and a Time Capsule in the Early History of Vertebrates.Geoscience Canada40:149-163.
Callier, V., Clack, J. A., & Ahlberg, P. E. (2009). Contrasting developmental trajectories in the earliest known tetrapod forelimbs.Science324:364-367.
Waddington, C. H. (1953). Genetic assimilation of an acquired character. Evolution 7:118-126.
Crispo, E. (2007). The Baldwin effect and genetic assimilation: revisiting two mechanisms of evolutionary change mediated by phenotypic plasticity. Evolution 61:2469-2479.
This is the mammoth image I remember, from a 1971 book, with no artist credited. It’s actually not as good as I remember, by modern standards at least.
Mammoths and I go way back, not quite to the Ice Age but at least to the late 1970s with my family’s visits to the University of Wisconsin Geology Museum, and Milwaukee Public Museum, to name two prominent places that inspired me. And one of my favourite science books had a colourful mammoth painting on the cover (above), an image that has stayed with me as awesomely evocative.
Stomach-Churning Rating: 3/10. But there’s a butt below, but that’s too late for you now. And there’s poo and other scatological (attempts at) humour. Otherwise, bones and a baby mammothsicle.
Fast forward to the 2000’s and I’m studying mammoths, along with their other kin amongst the Proboscidea (elephants and relatives). I even bumped into a frozen mammoth in Sapporo, Japan, nine years ago–
Yep. That’s what it looks like. Nope, not the front end. That dark orifice is not the mouth. This is a mammoth that was found on Bolshoi Lyakhovsky island, in the east Siberian arctic (New Siberian Islands archipelago), in 2003. Just think of finding this and being all excited then realizing, “Jackpot! Wait… Oh man, I just found the ass. I’ve discovered a mammoth bunghole, dammit.” Still, it’s pretty damn amazing, as frozen Ice Age buttocks go. I’d love to find one. I would not be bummed.
found on Bolshoi Lyakhovskiy island in 2003
What I know now that I didn’t realize as a kid, is that a mammoth is an elephant in all but name. Mammoths are more closely related to Asian elephants than either is to African elephants, and all of these elephants are members of the group Elephantidae. If we saw a smallish Columbian mammoth, we’d probably mostly look upon it as similar to a slightly hairy Asian elephant (but a scientist would be able to spot the distinctive traits that each has). Only woolly mammoths adopted the uber-hirsute state that we tend to think of as a “mammoth” trait. Think about it: a big animal would benefit most from a thick hairy insulation in an extremely cold habitat, and Columbian mammoths ranged further south than Woolly ones. No mammoths were radically different from living elephants, unless you count the dwarf ones. But as a kid, like most people do, I saw them as something else: an exotic monster of the past, eerily unlike anything today, and bigger too. And mammoths have the added mystique of the extinct.
Now I see mammoths as neither exotic nor that far in the past. Giant ground sloths, now those are still alien and exotic to me. I don’t get them. I know elephants pretty well, and I can understand mammoths in their light and in light of mammoth fossils. Various mammoth species persisted as late as maybe 10,000 (for the Woolly and Columbian species; the latter seeming to vanish earlier) to <4000 (for isolated Siberian forms) years ago, into quasi-historic times. And only some mammoths got larger than African elephants (Loxodonta) do, such as Columbian mammoths (~10,000 kg or more maximal body mass; Loxodonta is closer to 7-10 tonnes at best).
Lately, coincidence has brought me new knowledge of – and even greater interest in – mammoths.
First, a fortunate last-minute visit to Waco, Texas’s “Mammoth Site” (see my Flickr photo tour here) two weeks ago during a short visit to give a talk in that fine central Texan city.
Second, the subject of today’s post: the Natural History Museum’s new special exhibit “Mammoths: Ice Age Giants“, which is open until 7 September. The exhibit was created by the Field Museum in Chicago, but the NHM has given it a special upgrade under the expert guidance of mammoth guru Prof. Adrian Lister of the NHM, who was very kind to give me a tour of the exhibit.
What follows is primarily a photo-blog post and review of the exhibit, but with some thoughts and facts and anecdotes woven through it. Dark setting, glass cases, caffeination, crowds, and mobile phone camera rather than nice SLR in hand means that the quality isn’t great in my images– but all the more reason to go see the exhibit yourself! All images can be clicked to em-mammoth them.
On entry, one views a mammoth skeleton with a timelapse video backdrop that shows how the landscape (somewhere in USA) has changed since ~10,000 BCE.
The first part of the exhibit does a nice job of introducing key species of Proboscidea (elephants and their closest extinct relatives), with a phylogeny and timescale to put them into context, starting with the earliest forms:
from species like the tapir-sized Moeritherium…
Skull of Moeritherium, reconstructed. Not that different from an early sirenian (seacow) in some ways, and general shape, whereas still quite a long way from a modern elephant in form– but the hints of tusks and trunk are already there.
…To the early elephantiform Phiomia, here shown as a smallish animal but I’m told it actually got quite large. And continuing with giant terrestrial taxa…
I was awed by this reconstruction of the huge early elephantiform-relative Deinotherium, with the short, swollen trunk and downturned tusks– so bizarre!
Looking down onto the roof of the mouth of an NHM specimen of Deinotherium. Big, sharper-edged, almost rhino-like teeth; far from the single mega-molars of modern elephants.
The lower jaw (top) and fairly straight tusk (bottom) of the widespread, early elephantiform Gomphotherium.
The big “shovel-tusked” elephantiform Amebelodon. This was one of the earliest stem elephants I learned of as a kid; the odd tusks still stir wonder in me.
Amebelodon lower jaw, sans shovel tusks. Extended chin looks like some sort of childrens’ fun-slide. To me, anyway.
Next, there are some fun interactive displays of elephant biomechanics!
How would a mammoth hold up its head? This lever demonstration shows how a nuchal ligament helps. Tension on the nuchal ligament is a force that acts with a large lever (represented by the big neural spines on the vertebrae around the shoulders, forming the mammoths’ “hump” there), creating a large moment (i.e. torque; rotational force) that holds the head aloft.
I love this robotic elephant trunk demonstration. It captures some of the weirdness of having a muscular hydrostat attached to your lip and nostrils. Not so easy for a human to control!
But forget the myths about elephants having 40,000 to 150,000 muscles in their trunk. They have three muscle layers: a circumferential one, an oblique one and a longitudinal one. Like any muscles, especially ones this large, the layers each consist of many muscle fibres. That’s where the 40-150k myth comes from, but muscle fibres (cells) are at a more microscopic level than whole muscles (organs). Elephants do have excellent control of their trunks, but it’s not magical. It’s just different.
Then we come to the centrepiece of the exhibit, the ~42,000 year old Woolly mammoth (Mammuthus primigenius) baby “Lyuba“, which the NHM added to the original exhibit in this new version, as a star attraction — and a big win. Adrian Lister related to me how he’d never seen Lyuba in person before (access to it was tightly guarded for years). So when the NHM received the crate and held a press event to open it and reveal Lyuba, a journalist asked Adrian to act excited, to which he responded something like, “I don’t need to act! I’m very excited!” I would be, too! Full story on Lyuba’s arrival, by NHM site here. A key paper on Lyuba by Fisher et al. is here.
Studies of tooth growth in Lyuba reveal her gestation period (like living elephants, ~22 months), season of birth (early spring), and age at death (~1 month), among other information.
Here we can see the right ear, which was gnawed off along with the tail by dogs of the reindeer herders that found and retrieved Lyuba in 2006. Regardless, there’s loads of anatomy preserved!
A hump of juvenile “brown fat” sits atop the head and neck of Lyuba. This probably was metabolized during growth to warm the baby; brown fat is packed with mitochondria and thereby conducts what is called “non-shivering thermogenesis”. Furthermore, Lyuba has very strange flanges on the trunk (also visible in 1 other frozen mammoth specimen, but here preserved very clearly! What were they used for?). More details are visible postcranially…
The body was naturally “freeze-dried”, with the addition of later rounds of soaking in formalin and ethanol, leaving the body dessicated and stiff, permanently stuck in a lifelike pose as seen below:
Whole view from an exhibit panel (you cannot photograph the specimen but these are fair game!). Here we see hair on the right forearm and remnant of the ear, and the labia and nipples showing it is a female mammoth are also preserved. The head-hump is lost during growth, and the shoulder changes to change the Asian elephant-like convex curvature of the back into the characteristic humped-shoulder form of a mammoth. But ontogeny still reveals the evolutionary connection of Elephas and Mammuthus.
Lyuba and scientists studying her, which also shows how rigid the carcass is; one can almost stand it up. Inside the digestive tract, researchers found chewed up plant material that was probably dung eaten by the baby to gain vital bacterial digestive flora, and Lyuba had plenty of body fat and ingested milk, indicating that she did not starve to death. Rather, vivianite in the respiratory tract indicates drowning as the cause of her demise. Perfusion of the body by these vivianites may have helped to preserve the body.
Answering a question the public may be wondering about: is the hype about cloning a mammoth very soon true? Nope. Well addressed, including what to me is the urgent question: would cloning a mammoth be ethical?
The fourth part of the exhibit takes on a largely North American focus to first illustrate what mammoths were like biologically, and second to wow the visitor with some huge beasts in full body, full scale glory, as we shall see!
Mammoth hair! These samples and recent molecular studies show that mammoths were not ginger-coloured as we long thought, but rather the ginger color comes as the dark grey-brown-black colour fades postmortem, as a preservational artefact (story here). I didn’t know that; cool.
Mammoth chow! I liked this addition to the exhibit. This brought mammoth ecology closer to home for me.
Mammoth poop!
After the biology explanations, let there be megafauna!
Mammoth skull! A nice one, too.
Top predators of Ice Age North America: Arctodus (short-faced bear– does the short face mean they were happy, unlike a long face? Sorry but they never are shown as very happy, unless it is the joy of whupass) and Homotherium (the other sabre-toothed cat; not the longer-toothed Smilodon).
Skulls of North American (mega)fauna: left to right, top to bottom: horse, short-faced bear, giant ground sloth, then camel, sabretooth cat, rabbit, direwolf (viva Ned Stark!), and pronghorn antelope.
Mastodon (Mammut americanum) skeleton!
Mammoths (and perhaps mastodons, etc.) seem to have been wiped out by a combination of climate change and habitat fragmentation, combined with what this item symbolizes: human hunting. This beautiful piece is the main part of an atlatl, or javelin-hurling lever. It would have given Ice Age hunters the extra power they’d need to penetrate mammoth hide and cause mortal injuries. It is also a great tie-in to my recent post on the British Museum’s odd-animals-in-art.
Finally, the exhibit surveys the kinds of mammoths that existed- there is a huge reconstruction of a Columbian mammoth near the mastodon (above), then smaller kinds and discussions of dwarfism, which is another strength of NHM mammoth research:
Woolly mammoth lower jaw (right) and its likely descendant, the pygmy mammoth of the Californian coastline, Mammuthus exilis.
The world’s smallest mammoth (left), molar tooth compared with that of its much larger ancestor Palaeoloxodon. The status of Mammuthus creticus as a dwarf mammoth from Crete was cemented by Victoria Herridge and colleagues, including Adrian Lister at the NHM.
Pygmy mammoth reconstruction. Shorter than me. I want one!
In the end, from all that glorious proboscidean diversity we were left with just 2 or 3 species of elephantids today (depending on your species concepts; it’s probably worth calling the African forest elephant its own species, Loxodonta cyclotis). The exhibit closes with a consideration of their conservation and fate. Ironically, this elephant skull could not be mounted with its tusks on display, because that would be commercializing ivory usage– even though the whole point of the exhibit’s denouement is to explain why elephants need protection!
Reactions to the exhibit: the photos tell the tale. It’s undeniably great, in terms of showing off the coolness of mammoths, other proboscideans and Ice Age beasties, to the general public. I felt like the factual content and learning potential was good. It didn’t feel at all like pandering to the lowest common denominator like some other exhibits I’ve seen (cough, Dino Jaws, cough). I loved the reconstructions, which were top quality in my opinion. I could have done with some more real skeletons, yet more realistically the exhibit hall was already large and full of cool stuff. But give me a break: Lyuba. This trumps everything. Going to see a real friggin’ frozen mammoth baby buries the needle of the awesomeness meter on the far right. That’s pretty much all I need to say. The spectacle was a spectacle.
This exhibit shows a lot of work, a lot of thought, and a personalized NHM touch that reflects the actual research (even very recent work!) that NHM staff like Prof. Lister are doing with collaborators around the globe. What more could we want, a herd of cloned mammoth babies frolicking around and tickling guests with their flanged trunks? Don’t hold your breath.
You’ve got just over 2 months to see the exhibit. Don’t come complaining on September 8 “BBBBBbbbut I didn’t know, I didn’t think it would be that cool! I just thought there’d be a guy in a Snuffleupagus suit signing autographs!” You have a duty as a Freezerino to go bask in the frozen glory of these Ice Age critters. There may be an exam at the end. 🙂
Is the exhibit kid-friendly? More or less. The text is more targeted at teenager-level or so, but the visual impact is powerful without it. I’d warn a sensitive child about the withered baby mammoth body before showing it to them, so they aren’t caught off guard and scarred by the experience. I saw plenty of kids in the exhibit and they all seemed happy. Parents may want to linger longer and absorb all the interesting information, whereas kids may blitz through or goof around, so plan accordingly if you’re inbound with sprogs.
You know what I was eyeing up in the gift shop…
Aside: The frozen mammoths get me wondering- what else does the Siberian (or extreme northern Canadian/Scandinavian) permafrost conceal? There are a lot of awesome Ice Age megafauna I’d cut my left XXXXX off to study quasi-intact… think about how amazing it would be to find a giant ground sloth (not bloody likely), sabretooth cat, or other species. There’s a lot of north up north. A lot of space and ice. A lot could happen. And climate change will make discoveries like this more likely, while the melting (and humanity) lasts…
Wool we ever find the Lyuba of woolly rhinos (Coelodonta)? Cast of a mummified woolly rhino from the NHM’s entry hall. More of these finds are likely, I’d say.
At this writing (17 October, 2013), I am headed home after a 10-day trip to China as part of an RVC delegation participating in a London Universities International Partnership (LUIP) event (celebrating London innovations, especially those developed with Chinese input) as part of a broader UK/London-China trade mission. I am still processing what has been an astonishing, exhausting, exhilarating, chaotic, lavish, smog-ridden, and inspiring visit. As a simple scientist, I’ve found myself in the midst of major global politics, business and science policy, with little time to assimilate what has happened but still learning plenty about how the bigger world, way beyond my lab, operates. I thought I’d share that experience, by way of pictures illustrating key – or just unusual or interesting – events and places from my journey. It was surreal, in so many ways…
Stomach-Churning Rating: 0/10 except for a couple of odd statues. No squat-toilets; I will spare you those.
Odd decoration above entrance to the art gallery building that housed the LUIP event.
Several months ago the RVC selected me to help RVC Access director Nina Davies and colleagues set up an exhibit, as part of the LUIP event, featuring the work that my team has done, and is still doing, with Chinese collaborators at the IVPP in Beijing (exemplified by this past post). Dinosaurs and 3D computer modelling were thought to be a good potential draw for the public (ya think?) as opposed to more controversial subjects such as avian flu, with which the RVC also has research strengths and Chinese collaborations. I saw it as a great chance to go spend time at the IVPP’s spectacular fossil collection and develop ongoing collaborations with scientists there like Drs. Zhou Zhonghe and Xu Xing. Subsequently, I learned that it was a small enough event that I’d probably be meeting Boris Johnson (Mayor of London) there as well, possibly even presenting our research to him.
Hallway lined with art galleries, one of which is the Yang Gallery, which the event was held in.
The preparations for the exhibit were full of surprises, as you might expect a long-distance interaction between UK and Chinese people to be, especially if you’ve spent time in China and know some of the broad-brush cultural differences (e.g. “Yes” can mean no, and “maybe” usually means no). There were many cooks involved! Artists, policymakers, scientists, universities… and then the Mayor’s office got thrown into the action, and then it snowballed, with UK Higher Education and Science minister Rt Hon MP David Willetts coming to the LUIP event, and UK Foreign Chancellor George Osborne then scheduling a related trip to China at the same time. Meanwhile, I just supplied some images (courtesy of Luis Rey) and a video (by Vivian Allen and Julia Molnar) from our past paper to illustrate what we’re doing with Chinese collaborators.
There wasn’t time to prepare a fancy exhibit with lots of bells and whistles, but I was pleasantly surprised by what the LUIP organizers cooked up from what we provided, as photos below show. The addition of four great casts of fossils on loan from the IVPP was crucial and made us stand out from all the other exhibits in a big way! The event was held in the trendy 798 Art District in eastern Beijing, which is an old industrial area converted to a surprisingly bohemian, touristy area that still sports its rusting old industrial infrastructure, but bedecked with modern art! That really worked for me as a setting. This was my third visit to Beijing/China but my first time in this gritty area of the city, which I recommend spending an afternoon in sometime if you visit– the streets are lined with cafes and art galleries.
Boris bike and nice design of exhibits (placed on/around the giant letters LONDON). The back wall sports a Communist slogan, partly painted over, exhorting the workers to give their full effort for the glory of Chairman Mao or something (seriously). The building was once a weapons factory, I was told.
All the work we put into this event was a big deal to me, but as the event developed, and the schedule for my 10 day visit shifted almost daily as various political factions shuffled the LUIP and UK trade mission plans, I became aware of the vastly broader issues at play, and humbled by their scope. Sure, studying the 3D changes of dinosaur body shape across >225 million years is truly awesome to conduct, but the socio-political issues around the LUIP event boggled and baffled me. Issues like “How do we get more Chinese students to come study at London universities?”, “How do Chinese parents feel about their students studying to become veterinarians?” and “What are the key obstacles limiting UK-Chinese collaborations and how can they be resolved?” gradually eclipsed the technical, scientific issues in my mind, and I started to feel lost. I learned a lot from this eye-opening experience.
The rest of his post is mostly a photo blog to illustrate the goings-on, but I consider some psychological/philosophical matters toward the end.
The London innovation event lighting gets tested out– and looks sweet.
Boris arrives, and proceeds to tour the exhibits rather than give his speech as planned. But it worked out OK in the end; he had two exhibit tours and a speech in the middle.
Minister Willetts arrives and prepares to speak about UK higher education for Chinese students.
I give Minister Willetts a tour of our fabulous fossil casts.
Left to right = back in time through avian evolution, represented by Yixianornis, Pengornis, Jeholornis and Microraptor casts, courtesy of the IVPP.
Arguably one of the most important fossil finds (ever?), the “four-winged” dinosaur Microraptor.
Added benefit of thaw in UK-Chinese relations: Microraptors for everyone!!! Well, for me anyway. And a cast, not a real one. But still pretty damn cool, and now it’s in my office for comparative research and teaching. See?
Darwin greets Microraptor in my office.
Like I said at the start, I don’t have a profound insight from this trip, not yet if ever. But it has obviously made a strong impression on me. It has reinforced some thoughts about Big Life Stuff. With the jetlag, the big geopolitical issues, the foreign country, the opulence, and my research thrown into that heady brew (ahem, along with some Tsingtao beer), I became lost. And I liked it, even though I was totally clueless at times, just looking around wide-eyed at the events unfolding and hearing about the political manoeuvring behind the scenes (e.g. how would big figures like Boris and Willetts share the limelight? And the news media was playing up the question of whether Boris’s or Osborne’s contingents were “winning” in some sense of some struggle, even though ostensibly they are on the same Tory team).
But we’re all clueless; we’re all lost. In some ways that’s a good thing. We have work to do; broad landscapes to explore whether evolutionary or socioeconomic or whatnot. There are big questions left, and no easy answers sometimes. That’s a bad thing, too; if we were less lost in major issues like climate change or habitat destruction or gross imbalance in wealth/power, the world would be a better place.
Quite apropos! Rockin’ artwork found in the 798 art district surrounding the Yang Gallery.
I find it helpful at times to ground myself in the knowledge that I am lost just like everyone else. There are different ways we can get lost: such as in pondering how dinosaur anatomy and physiology transformed over the Mesozoic era, or in throwing ourselves into weighty issues of business and diplomacy in the real world. To pretend we’re not lost risks becoming foolhardy; to exemplify the Dunning-Kruger effect.
It might be helpful for others to remind themselves of this sense of being lost, and that we all feel it or at least should at times. Students may sometimes look to their professors and think they have some monopoly on wisdom, but they’re lost too, and surely in some ways more lost than any of their students.
Smaller scale dino art in a local shop.
Boris got a bit lost, too, when he came to my exhibit – pondering the dinosaur-bird fossils, he pondered out loud “There’s some bone that birds and reptiles both have that shows they’re related… the, umm, the ischium?” Not understanding what he meant by this (all tetrapods have an ischium), I redirected him, along with a reassuring comment that he’d done his homework. I did this a bit clumsily as the multitude of news cameras and lights and boom-mikes hovered around us in eager anticipation of Something Interesting Happening, and as his minders began to urge him to move onward through the LUIP exhibit. I noted the wrist of a dinosaur like Microraptor and how it already had the unusual wing-folding mechanism that modern birds now use during flapping flight or to keep their feathers off the ground when standing. He seemed to sort of like that, then shook my hand and said something like “very impressive, well done” and moved on to the next exhibit. (Willetts fared a bit better and stayed longer, but science is his business)
Random artwork from the Yang Gallery and around the 798 Art District follows… I liked the style. My kind of funky art. The statue above combines childlike toy aspects with sinister jingoistic imagery. And the next one, well… see for yourself.
In that brief, frantic conversation, we were both lost, and I think none the less of Mayor Johnson for it. He’d come off the plane, rushed to hotel and to the LUIP event, gave an impassioned speech about London and China, and then was whisked around between a dozen or so exhibits, pursued all the while by a throng of media and minders and gawkers- was he expected to know all the sundry details of maniraptoran evolution at that point? No. But we had some fun and smiled for the cameras and then it was all over as we spun off, reeling into our different orbits. I wouldn’t be surprised if, from time to time, a politician like Boris pinches himself and thinks privately, “Wow, these issues I am embroiled in are so convoluted. I am totally, utterly lost.” I think that’s a healthy thing, and I enjoyed repeated doses of that feeling during my trip. In science, we often deal with a sense of awe or wonder—that is the sunny side of being lost. The other side, which can coexist sometimes in duality with awe/wonder, is the more fearful/anxious side, like when you’re stuck in a foreign city far from your hotel; surrounded by alien, fantastic scenery; and night is falling but no taxis are around to take you back, and the locals are starting to watch you to see if you’ll do something stupid (this was me, briefly, after doing some evening mall-shopping in Shanghai). How we react to that duality is, in some way, our choice. I point to a scientist studying evolution and a creationist freaking out about the subject as a good example of two polar opposites in how an awesome topic in science can evoke very different reactions within that duality. A seasoned traveller who likes to throw themselves into a city and experience blissful, unpredictable immersion, and a worrisome tourist who can’t stray far from their tour group provide analogous examples. But I digress; this post is in danger of becoming lost… Enjoy some cool statues as the denouement. Get lost in the comments—what makes you have that sense of awe, or being lost, and how do you deal with it?
Yesterday I encountered the question that, as a scientist who has studied a certain chunky Cretaceous carnivore a lot, most deflates me and makes me want to go study cancer therapeutic methods or energy sources that are alternatives to fossil fuels (but I’d be useless at either). I will explain why this is at the end of the post.
The question stems from a new discovery, reported in Proceedings of the National Academy of Sciences (PNAS) and thus expected to be one of the more important or exciting studies this year (no, I’m not going to get into the issue here of whether these “high impact” journals include the best scientific research or the most superficial or hyped “tabloid” science; they publish both, and not in mutual exclusivity). It’s a broken Tyrannosaurus rex tooth embedded in a duckbill dinosaur’s tail bone, which healed after the injury, showing that the animal survived the attack.
If you’re with me so far, you might be making the logical leap that this fossil find is then linked to the hotbed of furious controversy that still leaves palaeontology in crisis almost 100 years after Lambe suggested it for the tyrannosaur Gorgosaurus. If the hadrosaur survived an attack from a T. rex, then T. rex was a habitual predator and OMG JACK HORNER AND OTHERS BEFORE HIM WERE WRONG!
And you’d be right.
My encounter with the question stemmed from an email from a science journalist (Matt Kaplan) that, as is normal practice, shared a copy of the unpublished paper and asked for comments from me to potentially use in an article he was writing for the science journal Nature’s news site. Here, then, was my off-the-cuff response:
“Ooh. I do have a pretty strong opinion on this. Not sure if you’d want to use it but here goes. I may regret it, but this hits my hot buttons for One of the Worst Questions in All of Palaeobiology!
The T. rex “predator vs. scavenger” so-called controversy has sadly distracted the public from vastly more important, real controversies in palaeontology since it was most strongly voiced by Dr Jack Horner in the 1990s. I find this very unfortunate. It is not like scientists sit around scratching their heads in befuddlement over the question, or debate it endlessly in scientific meetings. Virtually any palaeontologist who knows about the biology of extant meat-eaters and the fossil evidence of Late Cretaceous dinosaurs accepts that T. rex was both a predator and scavenger; it was a carnivore like virtually any other kind that has ever been known to exist.
While the discovery is nice evidence, it is not particularly exciting in a scientific sense and is only one isolated element from species that lived for hundreds of thousands of years, which to me changes nothing and allows no generalizations about the biology of any species, only the statement that at one point in time a Tyrannosaurus bit a hadrosaur that survived the encounter. There is no real substance to the controversy that T. rex was “either” a predator or scavenger. It is just something that scientists drum up now and then to get media attention. I hope that soon we can move on to more pressing questions about the biology of extinct animals, but the media needs to recognize that this is just hype and they are being played in a rather foolish way; likewise scientists that still feel this is an exciting question need to move on. Maybe this specimen will allow that. But somehow my cynical side leads me to suspect that this “controversy” will just persist because people want it to, regardless of logic or evidence. (bold font added; see below)
Great galloping lizards, I am so tired of this nonsense. Maybe there is educational value in showing how science deals with provocative half-baked ideas about celebrity species, but scientists in the community need to speak up and say what the real science is about. It’s not about this “controversy”. Modern palaeontology is so much better than this.
Sorry for the rant. Maybe it’s too extreme but I’m just fed up with this non-issue! I suspect a huge proportion of our field feels similarly, however.”
(I later redacted a bit of it where I got a little too excited and used the word “curmudgeon”; a mistake, as that could be seen as ad hominem rather than a term of endearment, and this issue is about the science and not the people, per se. That bit is redacted here, too. I’ve also redacted a sentence in which I made an opinion on whether the paper should have been published in PNAS; that is mostly irrelevant here. I was not a reviewer, and authors/reviewers/editors have to make that decision. This would be a massive tangent away from what this blog post is intended to be about! I know some of the authors and don’t want to offend them, but this is about the science and how it is represented to the world, not about these particular authors or even this paper itself.)
Importantly, Kaplan’s story did include my skeptical quote at the end. I am curious to see how many other news stories covering this paper go that far.
Would a T. rex prey on, or just scavenge — or have a great time racing — a giant chicken? (art by Luis Rey)
I will stop right here and acknowledge that I’ve published a lot on a somewhat related topic: how fast a T. rex could run or if it could run at all. To me, that’s a great scientific question that has consequences not only for the predator/scavenger false dichotomy, but also for general theories of locomotor biomechanics (can an animal the size of a large elephant run as well as or better than said elephant? What are the thresholds of size and maximal running/jumping/other athletic abilities and how do they vary in different evolutionary lineages? And so on.). I’ll defend the validity of that question to the bitter end, even if it’s a question I’ve grown a little (but only a little) tired of and generally feel is about as well settled as these things can be in palaeontology (see my review here). I’ll also defend that it has been a real controversy (I have plenty of old emails, formal rebuttals submitted by colleagues, and other discourse as evidence of this) since I tackled it starting in 2002 and sort of finishing by 2011. I am sensitive about the issue of hyping my research up– this is something I’ve been careful about. I set a reasonable bar of how much is too much, check myself continuously with reflective thought, and I do not feel I have ever really crossed that bar, away from science-promotion into darker realms. This is partly why I’ve stopped addressing this issue in my current work. I feel like the science we’ve done on this is enough for now, and to keep beating the same drum would be excessive, unless we discovered a surprising new way to address the questions better, or a very different and more compelling answer to them.
“T. rex: scavenger or predator?” was controversial back in 1994 when Horner published “The Complete T. rex”, where he laid out his arguments. Brian Switek covered this quite well in his post on it, so I will not review that history. There was a big Museum of the Rockies exhibit about it that toured the USA, and other media attention surrounding it, so Horner’s name became attached to the idea as a result. Other such as Lambe and Colinvaux had addressed it before, but their ideas never seemed to gain as much currency as Horner’s did. But this post is not about that.
What this post is about is a consideration of why this is still an issue that the media report on (and scientists publish on; the two are synergistic of course), if most scientists aware of past debates are in good agreement that a T. rex was like most other carnivores and was opportunistic as a switch-hitting scavenger-predator, not a remarkably stupid animal that would turn down a proper meal that was dead/alive. Indeed, the Nature news piece has a juicy quote from Horner that implies (although I do not know if it was edited or if important context is missing) that he has been in favour of the opportunistic predator-scavenger conclusion for some time. Thus, as Switek’s article notes, even the strongest advocates of the obligate scavenger hypothesis(?) have changed their minds; indeed, that 2011 blog post intimates that this had already happened at least 2 years ago.
For many years, nothing has been published in the main peer-reviewed literature that favours that extreme “obligate scavenger” hypothesis. If I am wrong and there is a scientific debate, where are the recent papers (say within the past 5 years) that are strong, respectable arguments in favour of it? I contend that it is a dead issue. And if it is just about the middle ground; i.e. what percent of its time did a T. rex spend hunting vs. scavenging; we have no clue and may never know, and it’s not a very interesting question.
But who then is feeding off of this moribund equine; this defunct tyranno-parrot?
In thinking about my reply to the journalist over the past 2 days, I am reminded again of my general feeling that this is no longer a question of scientific evidence; the important bit in bold font above. Maybe we just like this “hypothesis” or the “controversy”, or maybe we’re lazy and don’t want to have to hunt for real debates in science.
But who are “the people?” I do not feel that The Public should be blamed; they are the people that The Scientists and The Media ostensibly are seeking to inform about what the state of modern knowledge and uncertainty is in science. So when I get asked about the controversy after a public lecture, I always try to go into detail about it. I don’t sigh and say “go Google it”. Nor do I do this to a journalist. Indeed, I’ve generally headed this issue off at the pass and added a blurb to press releases/webpages explaining my T. rex research to explain how it relates to the non-controversy; example here.
I have to begin turning my finger of accusation away from scientists and toward some of the media, because they must play a huge role in the shennanigans. Yes, scientists should know better then to play this up as a valid, heated, modern controversy. That is true. Yet I have a feeling that the balance of blame should also fall heavily on the side of media (general and science news) that continue to report on this issue uncritically as a real controversy. Thus the general public thinks it still is, and scientists/journals keep issuing papers/press releases that it is, leading to more reporting on this “controversy”, and the beast refuses to die. Switek’s article is a good counter-example of balanced coverage with clear application of critical thinking.
This is trivially different from other non-controversies in palaeontology such as whether birds evolved from a subgroup of theropod dinosaurs and hence are dinosaurs by virtue of descent (consensus = yes). So it is reflective of a broader problem of not calling a spade a spade.
And it’s embarassing, to a scientist, as my quote above expressed, to see dead controversies trotted out again and again, feeding the public perception that they are not dead.
That’s what leaves me frustrated. When do the shennanigans end?
I am reminded of a quote from a Seinfeld episode:
“Breaking up is like knocking over a Coke machine. You can’t do it in one push. You gotta rock it back and forth a few times, and then it goes over.”– Jerry, from the episode “The Voice”.
But this predator/scavenger relationship-from-hell leaves me, as a specialist working in this general area, feeling like I am trapped under that fridge. Help!
That’s why I started off this long post talking about feeling deflated, or disappointed, when asked this question. I do feel that way. I have to admit, I sometimes evenfeel that way when a sweet young kid asks me that question. Deep inside, I wish they wondered about something else. I wish that science had reached them with a deeper, more contemporary question. But when a journalist asks me how I feel about a new paper that revisits the “controversy”, I feel embarassed for palaeontology. Can’t we get past this? It makes us look so petty, mired in trivial questions for decades. But we’re not like that. This is a dynamic, exciting, modern field, but every news story about non-issues in palaeontology just perpetuate bad elements of palaeontology’s image.
To the scientists— why don’t we put our foot down more and say enough is enough, this is a dead issue? We have a role not only in peer review, but also in communicating our views about published work to the media when asked (AND when not asked, as in this blog post). But if you call them on it, do they listen? Which brings me to…
To the media (science/general journalists etc; I know this is a huge category and please don’t think I am blaming 100% of journalists or assuming they are all the same; they are not!)– if scientists tell you that a “controversy” is not such, at what point do you accept their judgement and kill the story, or at least use that quote? Does that ever happen? In what way are you at the mercy of senior editors/others in such issues? What power do you have? Is a shift in the balance of editorial power needed, or even achievable, in your case or in good exemplar cases? I’d really like to hear your experiences/thoughts. I am sure there is a lot I am not understanding, and I know many journalists are in a tough situation.
To the public— You’re often being misinformed; you are the losers in this issue. How do you feel about all it? (While this post focuses on a very tiny issue, the T. rex scavenger/predator unending drama, it is also about a broader issue of how the media perpetuates controversies in science after they have already gone extinct.)
What did this post have to do with freezers? Nothing. I’m just (H)ornery. Although I was once filmed for a planned Discovery Channel film about scientists who find a frozen tyrannosaur in polar regions and have to decide what to do with it before it slips into a chasm and is lost forever. Probably better that this never aired; it was cancelled. Segue to this post.
The Berkeley cast of the Wankel (MOR555) specimen of T. rex. Will we ever see the end of the predator/scavenger non-issue?
Our 3D computer models of a basal dinosaur and bird, showing methods and key differences in body shape. The numbers at the bottom are museum specimen numbers.
At about the moment I’m posting this, our Nature paper (our more formal page here, and the actual article here) embargo is ending, drawing a 14+ year gestation to a close. The paper is about how dinosaur 3D body shape changed during their evolution, and how that relates to changes in hindlimb posture from early dinosaurs/archosaurs to birds; “morpho-functional evolution” sums up the topic. We used the 3D whole-body computational modelling that I, Allen and Bates (among others) have developed to estimate evolutionary changes in body dimensions, rather than focusing on single specimens or (as in our last study) tyrannosaur ontogeny. We’ve strongly supported the notion (dating back to Gatesy’s seminal 1990 Paleobiology paper) that the centre of mass of dinosaurs shifted forwards during their evolution, and that this shift gradually led to the more crouched (flexed) hind leg posture that characterizes living birds. Here is a movie from our paper showing how we did the modelling:
And here is a summary of our 17 computer models of archosaur bodies, shown as one walks along the tips of the phylogeny shown in the video (the models are not considered to be ancestral to one another; we used a common computer algorithm called squared-change parsimony to estimate ancestral state changes of body dimensions between the 16 numbered nodes of the tree):
But we’ve done much more than just put numbers on conventional wisdom.
We’ve shown, to our own surprise, that the shift of the centre of mass was largely driven by evolutionary enlargements of the forelimbs (and the head and neck, and hindlimbs, to a less strong degree), not the tail as everyone including ourselves has assumed for almost 25 years. And the timing of this shift occurred inside the theropod dinosaur group that is called Maniraptora (or Maniraptoriformes, a slightly larger group), so the change began in animals that were not flying, but not long before flight evolved (depending on whom you ask, what taxonomy they favour and what evidence one accepts, either the smaller clade Eumaniraptora/Paraves or the bird clade Aves/Avialae).
Now, if you don’t like the cliche “rewriting the textbooks”, do have a look through texts on dinosaur/early avian palaeobiology and you probably will find a discussion of how the tail shortened, the centre of mass moved forwards as a consequence, the caudofemoral musculature diminished, and theropod dinosaurs (including birds) became more crouched as a result. We did that to confirm for ourselves that it’s a pretty well-accepted idea. Our study supports a large proportion of that idea’s reasoning, but modifies the emphasis to be on the forelimbs more than the tail for centre of mass effects, so the story gets more complex. The inference about caudofemoral muscles still seems quite sound, however, as is the general trend of increased limb crouching, but our paper approximates the timing of those changes.
Figure 3 from our paper, showing how the centre of mass moved forwards (up the y-axis) as one moves toward living birds (node 16); the funny dip at the end is an anomaly we discuss in the paper.
A final implication of our study is that, because the forelimbs’ size influenced the centre of mass position, and thus influenced the ways the hindlimbs functioned, the forelimbs and hindlimbs are more coupled (via their effects on the centre of mass) than anyone has typically considered. Thus bipedalism and flight in theropods still have some functional coupling– although this is a matter of degree and not black/white, so by no means should we do away with helpful concepts like locomotor modules.
And in addition to doing science that we feel is good, we’ve gone the extra mile and presented all our data (yes, 17 dinosaurs’ worth of 3D whole body graphics!) and the critical software tools needed to replicate our analysis, in the Dryad database (link now working!), which should have now gone live with the paper! It was my first time using that database and it was incredibly easy (about 1 hour of work once we had all the final analysis’s files properly organized)– I strongly recommend others to try it out.
That’s my usual general summary of the paper, but that’s not what this blog article is about. I’ll provide my usual set of links to media coverage of the paper below, too. But the focus here is on the story behind the paper, to put a more personal spin on what it means to me (and my coauthors too). I’ll take a historical approach to explain how the paper evolved.
Embarassing picture of me before I became a scientist. Hardee’s fast food restaurant cashier, my first “real job”, from ~1999- no, wait, more like 1986. The 1980s-style feathered (and non-receding) hair gives it away!
Rewind to 1995. I started my PhD at Berkeley. I planned to use biomechanical methods and evidence to reconstruct how Tyrannosaurus rex moved, and started by synthesizing evidence on the anatomy and evolution of the hindlimb musculature in the whole archosaur group, with a focus on the lineage leading to Tyrannosaurus and to living birds. As my PhD project evolved, I became more interested and experienced in using 3D computational tools in biomechanics, which was my ultimate aim for T. rex.
In 1999, Don Henderson published his mathematical slicing approach to compute 3D body dimensions in extinct animals, which was a huge leap for the field forward beyond statistical estimates or physical toy models, because it represented dinosaurs-as-dinosaurs (not extrapolated reptiles/mammals/whatever) and gave you much more information than just body mass, with a lot of potential to do sensitivity analysis.
I struggled to upgrade my computer skills over the intervening years. I was developing the idea to reconstruct not only the biomechanics of T. rex, but also the evolutionary changes of biomechanics along the whole archosaur lineage to birds– because with a series of models of different species and a working phylogeny, you could do that. To me this was far more interesting than the morphology or function of any one taxon, BUT required you to be able to assess the latter. So Tyrannosaurus became a “case study” for me in how to reconstruct form and function in extinct animals, because it was interesting in its own right (mainly because of its giant size and bipedalism). (Much later, in 2007, I finally finished a collaboration to develop our own software package to do this 3D modelling, with Victor Ng-Thow-Hing and F. Clay Anderson at Honda and Stanford)
Me and a Mystery Scientist (then an undergrad; now a very successful palaeontologist!), measuring up a successful Cretaceous hypercarnivore at the UCMP; from my PhD days at Berkeley, ~2000 or so.
In all this research, I was inspired by not only my thesis committee and others at Berkeley, but also to a HUGE degree by Steve Gatesy, a very influential mentor and role model at Brown University. I owe a lot to him, and in a sense this paper is an homage to his trailblazing research; particularly his 1990 Paleobiology paper.
In 2001, I got the NSF bioinformatics postdoc I badly wanted, to go to the Neuromuscular Biomechanics lab at Stanford and learn the very latest 3D computational methods in biomechanics from Prof. Scott Delp’s team. This was a pivotal moment in my career; I became partly-an-engineer from that experience, and published some papers that I still look back fondly upon. Those papers, and many since (focused on validating and testing the accuracy/reliability of computer models of dinosaurs), set the stage for the present paper, which is one of the ones I’ve dreamed to do since the 1990s. So you may understand my excitement here…
Stanford’s Neuromuscular Biomechanics Lab, just before I left in 2003.
But the new paper is a team effort, and was driven by a very talented and fun then-PhD-student, now postdoc, Dr Vivian Allen. Viv’s PhD (2005-2009ish) was essentially intended to do all the things in biomechanics/evolution that I had run out of time/expertise to do in my PhD and postdoc, in regards to the evolution of dinosaur (especially theropod) locomotor biomechanics. And as I’d hoped, Viv put his own unique spin on the project, proving himself far better than me at writing software code and working with 3D graphics and biomechanical models. He’s now everything that I had hoped I’d become by the end of my postdoc, but didn’t really achieve, and more than that, too. So huge credit goes to Viv for this paper; it would never have happened without him.
We also got Karl Bates, another proven biomechanics/modelling expert, to contribute diverse ideas and data. Furthermore, Zhiheng Li (now at UT-Austin doing a PhD with Dr Julia Clarke) brought some awesome fossil birds (Pengornisand Yixianornis) from the IVPP in Beijing in order to microCT scan them in London. Zhiheng thus earned coauthorship on the paper — and I give big thanks to the Royal Society for funding this as an International Joint Project, with Dr Zhonghe Zhou at the IVPP.
That’s the team and the background, and I’ve already given you the punchlines for the paper; these are the primitive and the derived states of the paper. The rest of this post is about what happened behind the scenes. No huge drama or anything, but hard, cautious work and perseverance.
Me shortly after I moved to the RVC; video still frame from a dinosaur exhibit (c. 2004) I was featured in. Embarassingly goofy pic, but I like the blurb at the bottom. It’s all about the evolutionary polarity, baby!
The paper of course got started during Viv’s PhD thesis; it was one of his chapters. However, back then it was just a focus on how the centre of mass changed, and the results for those simple patterns weren’t very different from those we present in the paper. We did spot, as our Nature supplementary information notes, a strange trend in early theropods (like Dilophosaurus; to a lesser degree Coelophysis too) related to some unusual traits (e.g. a long torso) and suggested that there was a forward shift of centre of mass in these animals, but that wasn’t strongly upheld as we began to write the Nature paper.
On the urging of the PhD exam committee (and later the paper reviewers, too), Viv looked at the contributions of segment (i.e. head, neck, trunk, limbs, tail) mass and centre of mass to the overall whole body centre of mass. And I’m glad he did, since that uncovered the trend we did not expect to find: that the forelimb masses were far more important for moving the centre of mass forwards than the mass (or centre of mass) of the tail was– in other words, the statistical correlation of forelimb mass and centre of mass was strong, whereas changes of tail size didn’t correlate with the centre of mass nearly as much. We scrutinized those results quite carefully, even finding a very annoying bug in the 3D graphics files that required a major re-analysis during peer review (delaying the paper by ~6 months).
The paper was submitted to Nature first, passing a presubmission inquiry to check if the editor felt it fit the journal well enough. We had 3 anonymous peer reviewers; 1 gave extensive, detailed comments in the 3 rounds of review and was very fair and constructive, 1 gave helpful comments on writing style and other aspects of presentation as well as elements of the science, and 1 wasn’t that impressed by the paper’s novelty but wanted lots more species added, to investigate changes within different lineages of maniraptorans (e.g. therizinosaurs, oviraptorosaurs). That third reviewer only reviewed the paper for the first round (AFAIK), so I guess we won them over or else the editor overruled their concerns. We argued that 17 taxa were probably good enough to get the general evolutionary trends that we were after, and that number was ~16 more species than any prior studies had really done.
Above: CT scan reconstruction of the early extinct bird Yixianornis in slab conformation, and then Below: 3D skeletal reconstruction by Julia Molnar, missing just the final head (I find this very funny; Daffy Duck-esque) which we scaled to the fossil’s dimensions from the better data in our Archaeopteryx images. There is also the concern, which the reviewers didn’t focus on but I could see other colleagues worrying about, that some of the specimens we used were either composites, sculpted, or otherwise not based on 100% complete, perfectly intact specimens. The latter are hard to come by for a diversity of extinct animals, especially in the maniraptoran/early bird group. We discussed some of these problems in our 3D Tyrannosaurus paper. The early dinosauromorph Marasuchus that we used was a cast/sculpted NHMUK specimen based on original material, as was our Coelophysis, Microraptor and Archaeopteryx; and our Carnegie ??Caenagnathus??Anzu (now published) specimen was based more on measurements from 1 specimen than from direct scans, and there were a few other issues with our other specimens, all detailed in our paper’s Supplementary Information.
But our intuition, based on a lot of time spent with these models and the analysis of their data, is that these anatomical imperfections matter far, far less than the statistical methods that we employed– because we add a lot of flesh (like real animals have!) outside of the skeleton in our method, the precise morphology of the skeleton doesn’t matter much. It’s not like you need the kind of quality of anatomical detail that you need to do systematic analyses or osteological descriptive papers. The broad dimensions can matter, but those tend to be covered by the (overly, we suspect) broad error bars that our study had (see graph above). Hence while anyone could quibble ad infinitum about the accuracy of our skeletal data, I doubt it’s that bad– and it’s still a huge leap beyond previous studies, which did not present quantitative data, did not do comparative studies of multiple species, or did not construct models based on actual 3D skeletons as opposed to artists’ 2D shrinkwrapped reconstructions (the “Greg Paul method”). We also did directly measure the bodies of two extant archosaurs in our paper: a freshwater crocodile and a junglefowl (CT scan of the latter is reconstructed below in 3D).
One thing we still need to do, in future studies, is to look more carefully inside of the bird clade (Aves/Avialae) to see what’s going on there, especially as one moves closer to the crown group (modern birds). We represented modern birds with simply 1 bird: the “wild-type chicken” Red junglefowl, which isn’t drastically different in body shape from other basal modern birds such as a tinamou. Our paper was not about how diversity of body shape and centre of mass evolved within modern birds. But inspecting trends within Palaeognathae would be super interesting, because a lot of locomotor, size and body shape changes evolved therein; ostriches are probably a very, very poor proxy for the size and shape of the most recent common ancestor of all extant birds, for example, even though they seem to be fairly basal within that whole lineage. And, naturally, our study opens up opportunities for anyone to add feathers to our models and investigate aerodynamics, or to apply our methods to other dinosaur/vertebrate/metazoan groups. If the funding gods are kind to us, later this year we will be looking more closely, in particular, at the base of Archosauria and what was happening to locomotor mechanics in Triassic archosaurs…
Clickum to embiggum:
Australian freshwater crocodile, Crocodylus johnstoni; we CT scanned it in 3 pieces while visiting the Witmer lab in Ohio.
A Red junglefowl cockerel, spotted in Lampang, Thailand during one of my elephant gait research excursions there. Svelte, muscular and fast as hell. This photo is here to remind me to TAKE BLOODY PICTURES OF MY ACTUAL RESEARCH SPECIMENS SO I CAN SHOW THEM!
I’d bore you with the statistical intricacies of the paper, but that’s not very fun and it’s not the style of this blog, which is not called “What’s in John’s Software Code?”. Viv really worked his butt off to get the stats right, and we did many rounds of revisions and checking together, in addition to consultations with statistics experts. So I feel we did a good job. See the paper if that kind of thing floats your boat. Someone could find a flaw or alternative method, and if that changed our major conclusions that would be a bummer– but that’s science. We took the plunge and put all of our data online, as noted above, so anyone can do that, and that optimizes the reproducibility of science.
What I hope people do, in particular, is to use the 3D graphics of our paper’s 17 specimen-based archosaur bodies for other things– new and original research, video games, animations, whatever. It has been very satisfying to finally, from fairly early in the paper-writing process onwards, present all of the complex data in an analysis like this so someone else can use it. My past modelling papers have not done this, but I aim to backtrack and bring them up to snuff like this. We couldn’t publish open access in Nature, but we achieved reasonably open data at least, and to me that’s as important. I am really excited at a personal level, and intrigued from a professional standpoint, to see how our data and tools get used. We’ll be posting refinements of our (Matlab software-based) tools, which we’re still finding ways to enhance, as we proceed with future research.
Above: Two of the 17 archosaur 3D models (the skinny “mininal” models; shrinkwrapped for your protection) that you can download and examine and do stuff with! Dilophosaurus on the left; Velociraptor on the right. Maybe you can use these to make a Jurassic Park 4 film that is better, or at least more scientifically accurate, than Hollywood’s version! 😉 Just download free software like Meshlab, drop the OBJ files in and go!
Now, to bring the story full circle, the paper is out at last! A 4 year journey from Viv’s PhD thesis to the journal, and for me a ~14 year journey from my mind’s eye to realization. Phew! The real fun begins now, as we see how the paper is received! I hope you like it, and if you work in this area I hope you like the big dataset that comes with it, too. Perhaps more than any other paper I’ve written, because of the long voyage this paper has taken, it has a special place in my heart. I’m proud of it and the work our team did together to produce it. Now it is also yours. And all 3200ish words of this lengthy blog post are, as well!
Last but not least, enjoy the wonderful digital painting that Luis Rey did for this paper (another of my team’s many failed attempts to get on the cover of a journal!); he has now blogged about it, too!
Dinosaur posture and body shape evolving up the evolutionary tree, with example taxa depicted. By Luis Rey.
News stories about this paper will be added below as they come out, featuring our favourites:
Synopsis: Decent coverage, but negligible coverage in the general press; just science-specialist media, more or less. I think the story was judged to be too complex/esoteric for the general public. You’d think dinosaurs, evolution, computers plus physics would be an “easy sell” but it was not, and I don’t think we made any big errors “selling” it. Interesting– I continue to learn more about how unpredictable the media can be.
Regardless, the paper has had a great response from scientist colleagues/science afficionados, which was the target audience anyway. I’m very pleased with it, too– it’s one of my team’s best papers in my ~18 year career.