The early, hippo-like mammal Coryphodon. I didn’t know it had a patella but it does. From Yale Peabody Museum.
I’m not shy about my fondness for the patella (kneecap) of tetrapod vertebrates, and neither are the other members of RVC’s “Team Patella”. We’ve had a fun 3+ years studying these neglected bones, and today we’ve published a new study of them. Our attention has turned from our prior studies of bird and lepidosaur kneecaps to mammalian ones. Again, we’ve laid the groundwork for a lot of future work by focusing on (1) basic anatomy and (2) evolutionary history of these sesamoid bones, with a lot of synthesis of existing knowledge from the literature; including development and genetics. This particular paper is a sizeable monograph of the state of play in the perusal of patellae in placental and other synapsids. Here’s what we did and found, focusing mostly on bony (ossified) patellae because that allowed us to bring the fossil record better to bear on the problem.
Reference: Samuels, M., Regnault, S., Hutchinson, J.R. 2016. Evolution of the patellar sesamoid bone in mammals. PeerJ 5:e3103 https://doi.org/10.7717/peerj.3103
Stomach-Churning Rating: 1/10; bones and more bones.
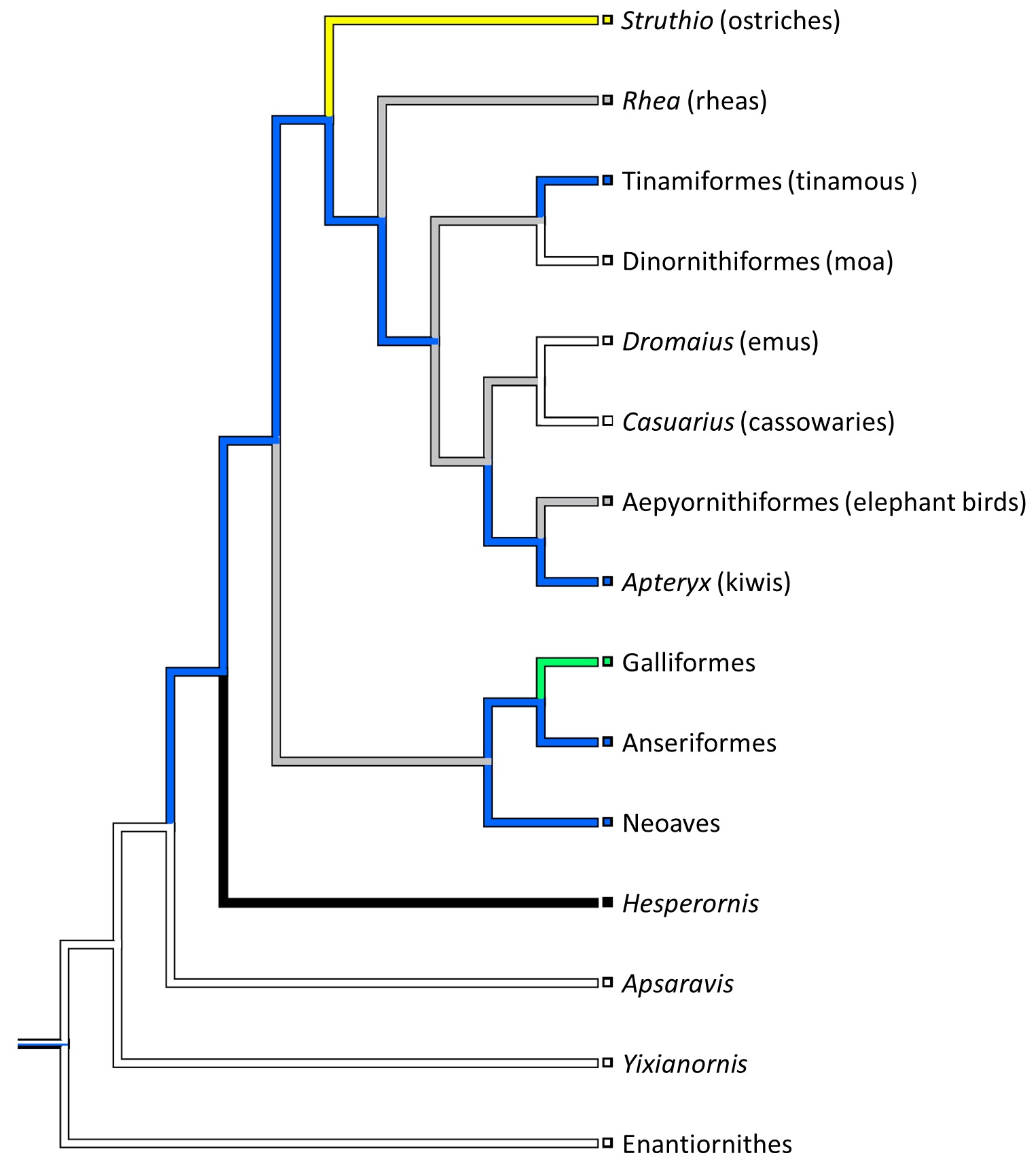
The short version of the story is that mammals evolved bony kneecaps about five times, with marsupials gaining and losing them (maybe multiple times) whereas monotremes (platypus and echidna) and placentals (us and other mammals) didn’t do much once they gained them, and a couple of other fossil groups evolved patellae in apparent isolation.

Evolution of the patella in mammals: broad overview from our paper. Click to zoom in.
The marsupial case is the most fascinating one because they may have started with a fibrocartilaginous “patelloid” and then ossified that, then reduced it to a “patelloid” again and again or maybe even regained it. There needs to be a lot more study of this group to see if the standard tale that “just bandicoots and a few other oddballs have a bony patella” is true for the Metatheria (marsupials + extinct kin). And more study of the development of patellae in this group could help establish whether they truly do “regress” into fibrocartilage when they are “lost” in evolution, or if other, more flexible patterns exist, or even if some of the cases of apparent “loss” of a bony patella are actually instances of delayed ossification that only becomes evident in older adults. Our paper largely punts on these issues because of an absence of sufficient data, but we hope that it is inspiration for others to help carry the flag forward for this mystery.

The higgledy-piggledy evolution of a patella in Metatheria, including marsupials. Click to zoom in.
Some bats, too, do funky things with their kneecaps, analogous to the marsupial “patelloid” pattern, and that chiropteran pattern also is not well understood. Why do some bats such as Pteropus fruit bats “lose” their kneecaps whereas others don’t, and why do some bats and other species (e.g. various primates) seem to have an extra thing near their kneecaps often called a “suprapatella”? Kneecap geeks need to know.

The short-nosed bandicoot (marsupial) Isoodon, showing a nice bony patella as typifies this group. From Yale Peabody Museum.
Otherwise, once mammals evolved kneecaps they tended to keep them unless they lost their hindlimbs entirely (or nearly so). Witness the chunky patellae of early whales such as Pakicetus and join us in wondering why those chunks persisted. The evolutionary persistence of blocky bits of bone in the knees of various aquatic animals, especially foot-propelled diving birds, may help answer why, as the hindlimbs surely still played roles in swimming early in cetacean evolution. Ditto for sea cows (Sirenia) and other groups.

Early whale Ambulocetus, showing hefty kneecaps.
But I’m still left wondering why so many groups of land vertebrates (and aquatic ones, too) never turned parts of their knee extensor tendons into bone. We know a bit about the benefits of doing that, to add leverage to those joints that enables the knee muscles to act with dynamic gearing (becoming more forceful “low gear” or more speedy “high gear” in function). Non-avian (and most early avian/avialan) dinosaurs, crocodiles, turtles, amphibians, early mammal relatives, and almost all other known extinct lineages except for those noted above got by just fine without kneecaps, it seems, even in cases where a naïve biomechanist would expect them to be very handy, such as in giant dinosaurs.

A quoll, Dasyurus, with what is probably a fibrocartilaginous “patelloid”. From Yale Peabody Museum.
However, tendons don’t turn to bone unless the right stresses and strains are placed upon them, so maybe kneecaps are a “spandrel” or “exaptation” of sorts, to abuse Gould’s ghost, whose adaptive importance is overemphasized. Maybe that adaptive myopia overshadows a deeper ontogenetic story, of how tissues respond to their history of mechanical loading environment. It has been speculated that maybe (non-marsupial) mammals have broadly “genetically assimilated” their kneecaps, fixing them into semi-permanence in their genetic-developmental programmes, whereas in contrast the few studies of birds indicate more responsiveness and thus less assimilation/fixation. That “evo-devo-mechanics” story is what now fascinates me most and we’ve poked at this question a bit now, with some updates to come- watch this space! Regardless, whether an animal has a bony vs. more squishy soft tissue patella must have consequences for how the knee joint and muscles are loaded, so this kind of question is important.

Giant marsupial Diprotodon (at NHM London); to my knowledge, not known to have had kneecaps- why?
In the meantime, enjoy our latest contribution if it interests you. This paper came about when first author Dr. Mark Samuels emailed me in 2012, saying he’d read some of my old papers on the avian musculoskeletal system and was curious about the evolution of patellae in various lineages. Unlike many doctors and vets I’ve run into, he was deeply fascinated by the evolutionary and fossil components of patellae and how those relate to development, genetics and disorders of patellae. We got talking, found that we were kindred kneecap-spirits, and a collaboration serendipitously spun off from that, soon adding in Sophie. It was a blast!