Jason Anderson, vertebrate paleontologist and anatomist at the University of Calgary (Canada), shared these two intriguing photos with me, and agreed for me to share them with you. Yay, thanks Jason! Good timing for a badly needed Mystery Dissection post:
What are they (species ID) *AND* from what region of the body?
(they are the same region, same specimen, same animal)
RULE: Your answer must be in the form of a winter holiday song (at least four lines)!
If you’ve been following some of my recent tweets, I’ve been tweeting about the “joys” of increased academic paperwork around holiday-time; e.g. this one:
Here, I give you a long-planned post on the patella (“kneecap bone”) of birds, which was my Royal Society Senior Research Fellowship sabbatical project for 2012-13. This is only a brief introduction to the anatomical issues at hand, err, I mean at knee…
Stomach-Churning Rating: 6/10; mostly skeletons/fossils, but there are a few images of the dissection of a guineafowl, which is fresh and meaty.
Archaeopteryx, the Berlin specimen. Helluva fossil, but nary a patella!
The question I am exploring, first of all, is simply how the patella evolved, because it seems to be present in almost all living birds. However, it is absent in all non-avian dinosaurs, and indeed most Mesozoic birds, too. There is barely a hint of any precursor structure (a “patelloid”) in other reptiles, but lizards evolved their own patella that is quite different (a flattened lozenge, not a rectangular structure lying tightly confined in a “patellar groove” on the femur as it is in birds). Mammals evolved the knobbly, hemispherical kind of kneecap that we’re familiar with, possibly on several occasions (a different story!). So the patella evolved at least three times in the lizard, mammal and bird lineages– and possibly more than once in each of these groups. And that’s about it for almost 400 million years of tetrapod evolution, except for a few very rare instances in fossils and sort-of-patella-like things in some frogs or other weirdos.
Fossil birds exhibit no clear presence of a patella until we come very close to modern birds on the avian stem of the tree of life (see below). And then, suddenly in modern birds, there is a lot of variation and not much good documentation of what kind of patella exists. This makes it challenging to figure out if the patella is ancient for modern birds or if it evolved multiple times, or how it changed after it first evolved– let alone bigger questions of what the patella was “for” (performance benefits, functional consequences, etc.; and developmental constraints) in the birds that first evolved it.
Considering that the patella is such an obvious bone in some birds, and certainly affects the mechanics of the knee joint (forming a lever for the muscles that cross it; homologous to our quadriceps muscles) and hence locomotion, it is a compelling research topic for me.
What follows is a pictorial guide to the patella of some birds, in sort of an evolutionary/temporal sequence (see my earlier post for a recap of some major groups), with a focus on animals I’ve studied more intensively so far (with >10,000 species, there is a lot that could be done):
The early Cretaceous bird Gansus (from the IVPP in Beijing), represented by many beautifully preserved specimens, all of which lack a patella. This absence is characteristic of other stunningly preserved fossil Chinese birds, indicating that this is almost certainly an ancestral absence of a patella, until…
The famed Cretaceous diving (flightless) bird Hesperornis, from Wikipedia/Smithsonian. Note the massive, conical/crested patella in front of the knee (jutting up and overlapping the ribs/vertebrae close to the pelvis; see also below). That elongate patella is characteristic of many diving birds that use foot-propelled swimming; it has evolved many times in this fashion. Other hesperornithiform birds show some transformational states in their anatomy toward this extreme one.
Check this out! More Hesperornis (cast), with the femur on the left and the patella on the right. The bloody patella is almost as long as the femur! That’s nuts. With kind permission from the Natural History Museum, London.
Exhibited ostrich (Struthio camelus) skeleton in left side view showing the patella (white arrow), on exhibit atThe Natural History Museum at Tring, Hertfordshire, UK. Ostriches are remarkable in that they have this elongate patella (actually a double patella; there is a smaller, often-overlooked second piece of bone) and yet are rather basal (closer to the root of the modern avian family tree)– however, they obviously are specialized in ways other than this double patella, most notably their very large size, flightlessness, and elongate legs. So the unusual patella is more likely linked to their odd lifestyle than a truly primitive trait, at least to some degree (but stay tuned: what happened with the patella in other members of their lineage, the ratites/palaeognaths, is much less well understood!).
Note that ostriches and Hesperornis together hint that the presence of a patella might have been an ancestral trait for living birds, but their patellae are so different that the ancestral state from which they evolved must have been different, too; perhaps simpler and smaller. Hence we need to look at other birds…
Skinned right leg of a Helmeted Guineafowl, Numida meleagris, above. That whitish band of tissue in the middle of the screen, on the front of the knee, is part of what is concealing the patella. That is an aponeurosis (connective tissue sheet, like a thin tendon) of the muscles corresponding to our “quads” or our tensor fascia latae, detailed more below. Guineafowl are fairly basal and well-studied in terms of their bipedal locomotion, so they are an important reference point for avian form and function.
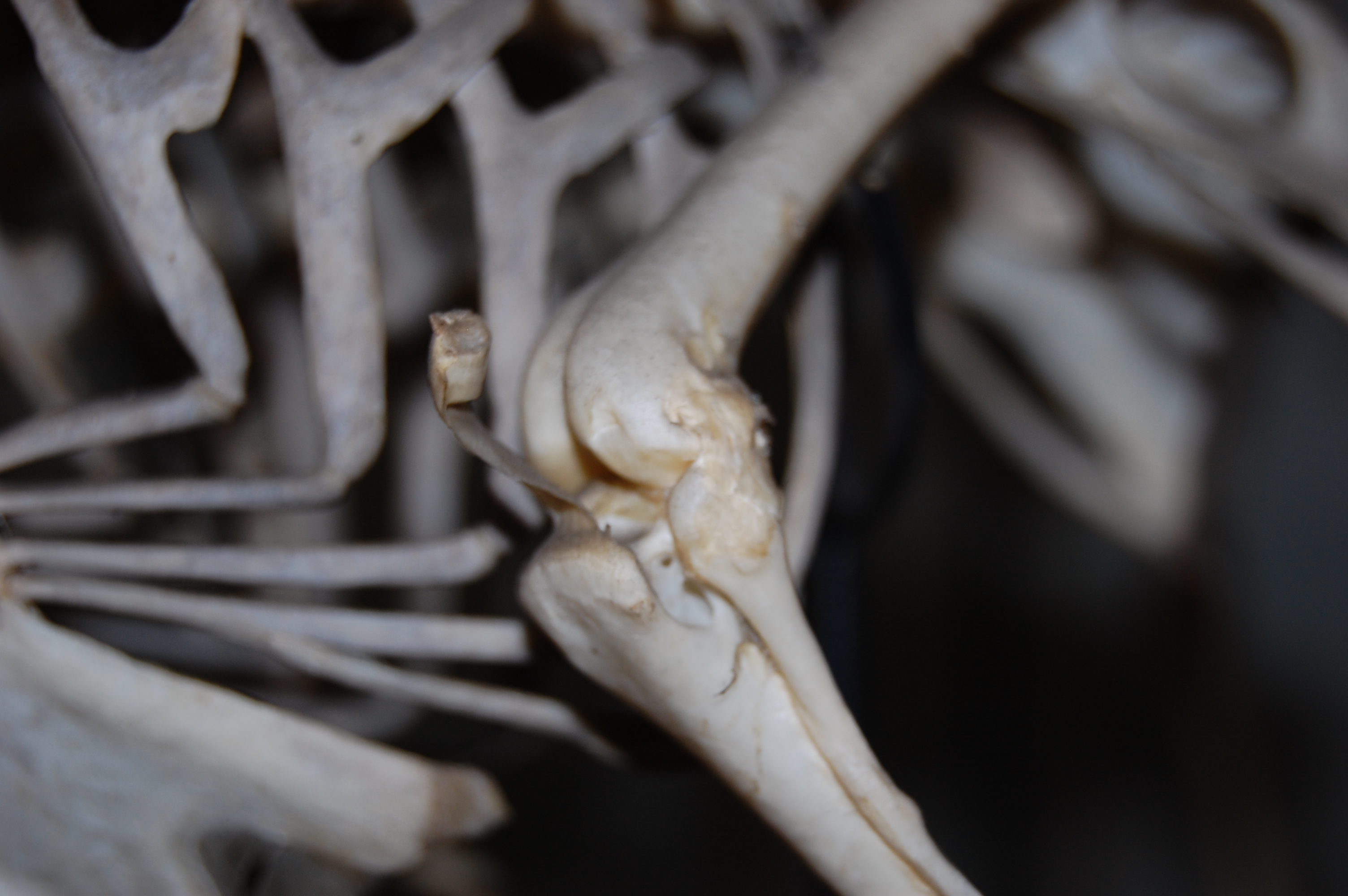
Right guineafowl leg, with patella exposed. Here I’ve peeled away that white band of tissue and associated muscles, which have been reflected toward the bottom of the screen (AIL and PIL labels corresponding to the anterior and posterior parts of the Iliotibialis lateralis muscle). The tip of the scalpel is contacting the patella. It’s not much to see, but lies atop the bright yellow fat pad that cushions it against the femur. You should be able to see a groove in the end of the femur just above that fat pad, which is where the patella sits and slides up and down as the knee moves/muscles contract. This is called the patellar groove, or sulcus patellaris.
Left leg of another guineafowl (with right tibiotarsus behind it, on the left) showing the patellae in articulation; in medial (inside) and cranial (front) views, respectively. The patella is the little rectangular bit of bone in the top middle of the screen, interposed between femur (thigh) and tibiotarsus (shank).With kind permission from the Natural History Museum, London.
Right leg of a Cape Penguin (Spheniscus demersus) from the University Museum of Zoology in Cambridge, showing the big lumpy patella in this wing-propelled diver. They still walk long distances on land, so presumably a patella plays some role in their gait, helping to explain its large size, which like the ostrich and Hesperornis seems to be a novel trait. Notice the groove across the patella, made by the tendon of the ambiens (like our sartorius/”tailor’s muscle”), which crosses from the inside to the outside of the leg via this route. This groove is often considered a useful phylogenetic character in modern birds, as its contact with the patella (sometimes via a hole, or foramen) varies a lot among species.
A hornbill, Buceros sp., from the UMZ Cambridge museum as well. This displays the possibly-more-typical, little rounded patellar nubbin that many birds have. See below for more.
Closeup of the knee/patella of the hornbill, Buceros sp., from above. Not much to squawk about, patella-wise, but it’s there.
And so we complete our quick tour of the avian patella, in its grand variation and humble beginnings.
Why does an ostrich have a patella and a Tyrannosaurus, Edmontosaurus or Triceratops did not? Why were birds the only bipedal lineage to evolve a patella (mammals and lizards gained a patella as small quadrupeds), and why did some bipeds like kangaroos “lose” (reduce to fibrous tissue, apparently) their patella?
These are the kinds of mysteries my group will now be tackling, thanks to a generous Leverhulme Trust grant on sesamoid bone ontogeny, mechanics and evolution. My group is now Dr. Vivian Allen as the postdoc, Sophie Regnault as the PhD student, and Kyle Chadwick as the technician and MRes student, along with numerous collaborators and spin-off projects. We’re looking forward to sharing more! But for now, I hope that I’ve engendered some appreciation for the avian patella, as the silly title indicates (“fella” used in the general sense of anyone!). This work is all unpublished, but some of this should be out in not too long, in much more lavish detail! Much as the patella is the “forgotten lever “of the avian hindlimb, it is the fulcrum about which a substantial part of my research group’s activity now pivots.
Well lookie here… it’s a new Mystery CT slice challenge! And it’s appearing while Society of Vertebrate Paleontology members are busy drinking and eating at the conference’s welcome reception– how naughty of me!
What is it, what species, etc.– tell me what you can.
RULE: Your answer must involve excessive alliteration!!!
Prodigious perambulations of appropriate prose promise to procure prodigious points!
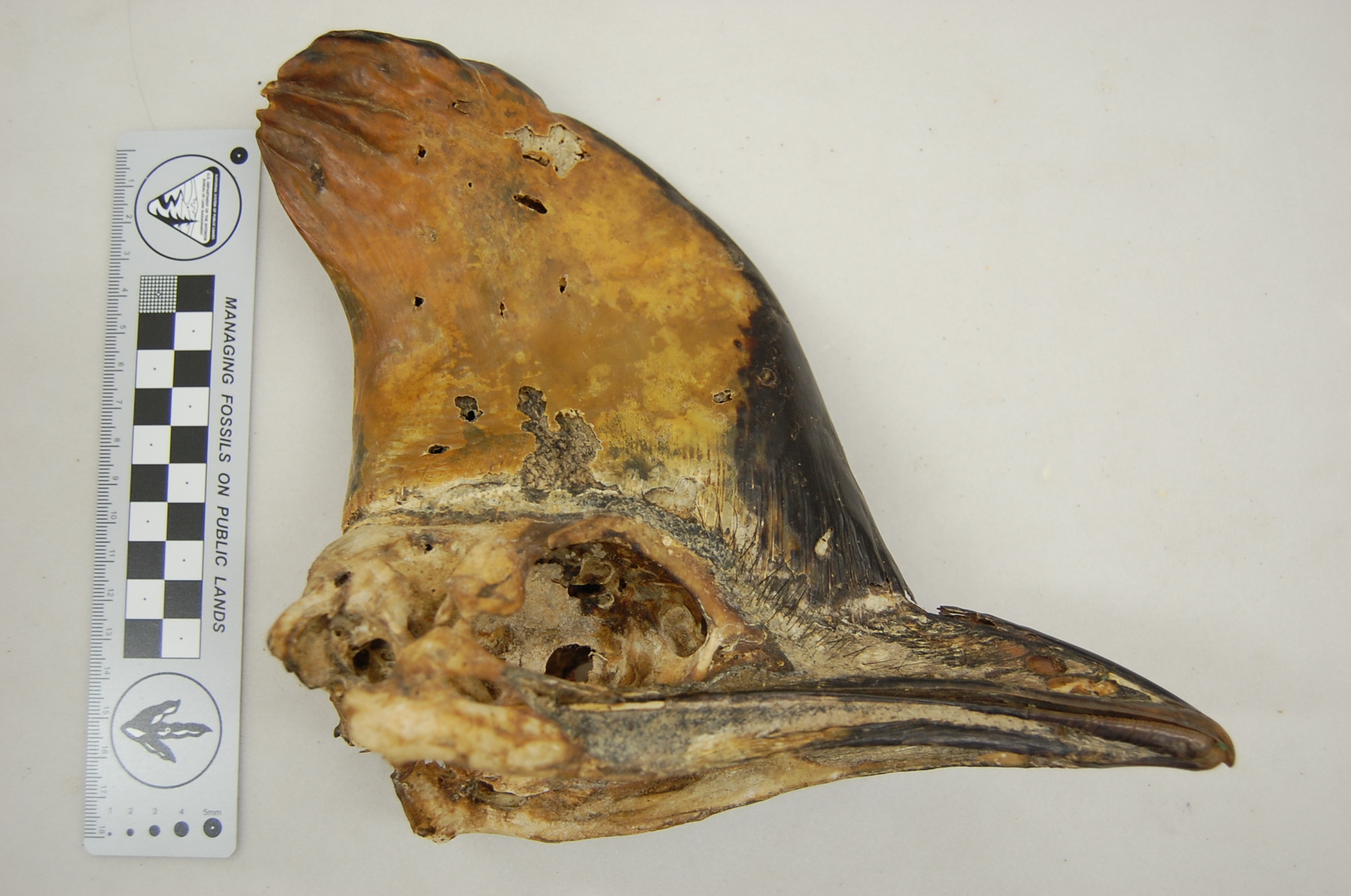
EDIT: These images give the answer and show some cool features. It’s an Asian elephant skull, NHMUK 1984.516, of a juvenile animal (probably a UK zoo animal). Gotta love that pneumaticity!
Hey, Christofer Clemente sent me these photos of a ~10cm long skull found by a student on a beach in Perth, Australia ( sent to Chris by Christine Cooper from ECU in Perth). What is it?
I figured I’d feed it to our mystery anatomy gurus for some fun, and see what comes up. You must justify your answer to get points. Convincing us with links to other photos/images will help. I have my ideas what it is, but they kind of suck, because I suck at skull anatomy.
Bonus points for incorporating Australian slang into your answer or beachy imagery/jokes.
No rhyming, unless the rhyming is using silly Australian-ish lingo like adding “-idoo” to the end of a word, or making up Dr-Seussian Oz-ish words.
My team had a new technician arrive, Kyle Chadwick from Uni. Virginia, and NSF Postdoctoral Research Fellow, Dr. Ashley Heers (see here for an example of new stuff she’s starting here at the RVC!), started working with me at the RVC, and then these guys showed up…
First a tiger salamander (Ambystoma) paid a visit, for filming an episode of the Windfall Films/PBS documentary “Your Inner Fish” (a la the famous book):
So cute! Tiger salamander, soon to be a TV celebrity.
And that gorgeous salamander was a star performer in strutting his stuff for the camera to demonstrate the locomotion of modern tetrapods, including some lovely slo-mo footage from our lab cameras:
(if that’s too slow for you, try the normal-speed footage. I’ll admit, salamanders don’t really need slo-mo video for normal walking, but I like it)
So cool!
But then we got a special package… with three frozen fire salamanders (Salamandra salamandra) from colleagues in Germany!
Three new occupants of the freezers, for planning our studies of salamander locomotion
This marks the start of an exciting new period in my team’s work in the lab. I’ve always liked salamanders and newts, and we’ve scanned and modelled plenty (e.g. this old post), but now we’re going to work with live fire salamanders (a first for me)! We are using the dead ones to plan the new studies with the live ones– these new studies will involve lots of high speed videos and force platform analysis (as shown above), in conjunction with XROMM (biplanar fluoroscopy/3D skeletal motion analysis) and other techniques including computer simulations. We got initial approval this week to work with these salamanders, and found a reputable source this week too, so it was definitely Salamander Week in my group!
This research all will feed into our upcoming studies of extinct tetrapods: we’re using salamanders to figure out how salamanders move and what limits their speed and gait, and then we’re using the same sorts of computer tools to try to estimate how extinct tetrapods may have moved and how locomotion evolved, in much more specific detail than our prior work had done, which was mainly about using 3D reconstructions of anatomy to show what those animals could not do. More about the project here.
Watch this space for more scampering salamanders!
UPDATE: And here’s one! Not quite scampering, but…
Setting up our two fluoroscopes for a test run of our gait studies– but with one of the deceased salamanders. Gotta get good images before any live animal work begins!
An example of the kind of footage we’re aiming for (single 2D fluoroscope view from Nadja Schilling’s team’s research; see XROMM website for more details on the methodology)
UPDATE 2:
I did a CT scan with a normal medical grade CT scanner at the highest resolution we can manage (0.625 mm slices). Check out the results below, which amuse me:
Looks like a toy; too crude resolution. But we can see major structures, and we can very nicely see the “microchip” (which looks HUGE) that was placed in this animal’s back when in captivity, and then another structure is visible near the pelvis which might be another chip or else remains of some food, pathology, or a really odd pelvis– I am not totally sure!
So this is why we tend to use microCT, which can go down to as low as ~5 micron resolution, to get 3D anatomy of animals this small. It’s no surprise to me, but it is fun to see how far we could push our normal CT machine. The results aren’t horrid but wouldn’t have much scientific value for us. They did confirm for us that this specimen is heavily ossified, so the faint images of bone that we are getting in our x-ray fluoroscopes (above) are due to something going wrong with our camera system, not the animal’s immature skeleton. Stay tuned for more updates as the science happens!
UPDATE 3:
20 wonderful adult Fire Salamanders have joined our team and are relaxing over the coming week before we start taking them for walks. Here is one exploring its new home:
UPDATE 4:
August 11-15, 2014 we are in Jena, Germany using their fancy biplanar radiography system (“x-ray video”) to study our salamanders, at last! Follow the tweets starting here, for more information as it happened! https://twitter.com/JohnRHutchinson/status/500187568416518144
and this video of “Jabba” the corpulent salamander walking-
Hola from Barcelona, where 500ish of us are telling each other about the latest research in the field of morphology (like anatomy, but broader, deeper, more explanatory; but if you prefer to think of it as anatomy that’s OK by me)!
#ICVM and #ICVM2013 (favoured) are the hashtags, and http://icvm2013.com/ is the website, and there’s Facebook and all that too! You can read the full programme and abstracts here. It’s the best damn conference in the universe and I am not remotely biased. It happens every 3 years somewhere in the world and is always chock full of 5 days of glorious new information on animal form and function and much more, with just too many interesting talks to ever be able to take it all in.
I am speaking a few times and want to share a talk that is about sharing the glory of morphology in public.
Morphology research, that is; please put your clothing back on!
It’s a text-heavier talk than my rules-of-conference-talks normally would allow, but I’m going for it, as that makes it better for sharing because my dulcet tones will not accompany the version I am sharing online. Someday in the future, at a conference venue that is better set up for reliably live-broadcasting a talk (this is NO FAULT of the excellent organizing committee of ICVM/ISVM!), I would just do it live, but not today, not here.
The point of the talk should be obvious from the first slide (as in my last post). But I’ll presage it by saying that another subtext, which might not come through so strongly in the slides as opposed to my spoken words, is that we need to tell people that we’re doing morphology/anatomy research! We should not be shy of that label because deans or geneticists or conventional wisdom or what/whomever might say (very, very wrongly!) that it is a dead or obsolete science.
While natural history, evolution, palaeontology and other fields allied to morphology do pretty well in the public eye, I don’t see people often reminded that what they are being told about in science communication is a NEW DISCOVERY IN ORGANISMAL MORPHOLOGY and that we are still discovering such new things about morphology all the freaking time! (e.g. my team’s research on elephant false sixth toes, or Nick Pyenson‘s team’s research on whale chin sense organs to name just 2 such studies, both published on the same day in Science!)
Indeed, many of those discoveries such as new fossils/exotic living things with cool features, cool developmental mechanisms that produced complex structures, or insights into how organisms are able to do amazing things are implicitly morphological discoveries, but the fanfare too often goes to natural history, palaeontology, evo-devo or some other area rather than explicitly to morphology.
In contrast, I too often hear people poo-pooing anatomical research as yesterday’s science.
Vesalius’s classic skeleton (from Wikipedia), which is great but to me also conjures misleading connotations of anatomy as a defunct discipline that old dead dudes did.
We need to sell ourselves better not only in that regard, of a renaissance of discoveries and insights in our field, but also in the sense of being in a renaissance that is driven by TOTALLY AWESOME TECHNOLOGICAL AND METHODOLOGICAL ADVANCES, especially computerized tools. We’re just as fancy in terms of techy stuff as any other biologists, but we don’t shout it from the rooftops as much as other disciplines do.
We’re not just primitive scientists armed only with scalpels and maybe a ruler now and then, although that simple approach still has its sublime merits. We’re building finite element models, running dynamic computer simulations, taking high-resolution CT or synchotron scans, manipulating embryos, digging up fossils, sequencing genes– you name it, morphologists may be doing it! (For similar views see Marvalee Wake’s recent review of herpetology & morphology; I’m by far not the first person to make the arguments I’m making in my talk, but I am putting a personal spin on them)
And of course, as the talk is being delivered by me, you might rightly expect that I’ll say that we need to do more of this kind of cheerleading where we have maximal visibility and interaction, which includes online via social media, etc. I’ll discuss one other venue which has featured prominently here on this blog, too: documentaries. Oh I’m not done with that hobby horse, no sirree, not by a long shot!!
Anyway I should get back to preparing my talk but here is the link to the slideshow (props to Anne Osterrieder for the inspiration to put my slides up here):
Please discuss anything related to this topic in the Comments– I’d love to hear what you think!
I am happy to clarify what my shorthand notes in the slide text mean if needed. There are links in the talk to other sites, which you can click and explore.
A photo blog post for ya here! I went to Dublin on a ~28 hour tour, for a PhD viva (now-Dr Xia Wang; bird feather/flight evolution thesis) earlier this month. And I made a beeline for the local natural history museum (National Museum of Ireland, Natural History building) when I had free time. So here are the results!
Stomach-Churning Rating: Tame; about a 1/10 for most, but I am going to break my rule about showing human bodies near the end. Just a warning. The bog bodies were too awesome not to share. So that might be 4/10-8/10 depending on your proclivities. They are dry and not juicy or bloody, and don’t look as human as you’d expect.
Simple Natural History museum entrance area.
Adorable frolicking topiaries outside the NHM.
Inside, it was a classical Victorian-style, dark wood-panelled museum stuffed with stuffed specimens. It could use major refurbishment, but I do love old-fashioned exhibits. Get on with it and show us the animals; minimize interpretive signage and NO FUCKING INTERACTIVE COMPUTER PANELS! So by those criteria, I liked it. Some shots of the halls: And on to the specimens!
Giant European deer (“Irish elk”). I looked at these and thought, “why don’t we see female deer without antlers ever? then noticed one standing next to these (you can barely see it in back); too bad my photo is crappy.
Superb mounted skeleton of giraffe (stuffed skin was standing near it).
A sheep-y or a goat-y beastie; I dunno but it shows off a nice example of the nuchal ligament (supports the head/neck).
Yarr, narwhals be internet gold!
Giant blown glass models of lice!
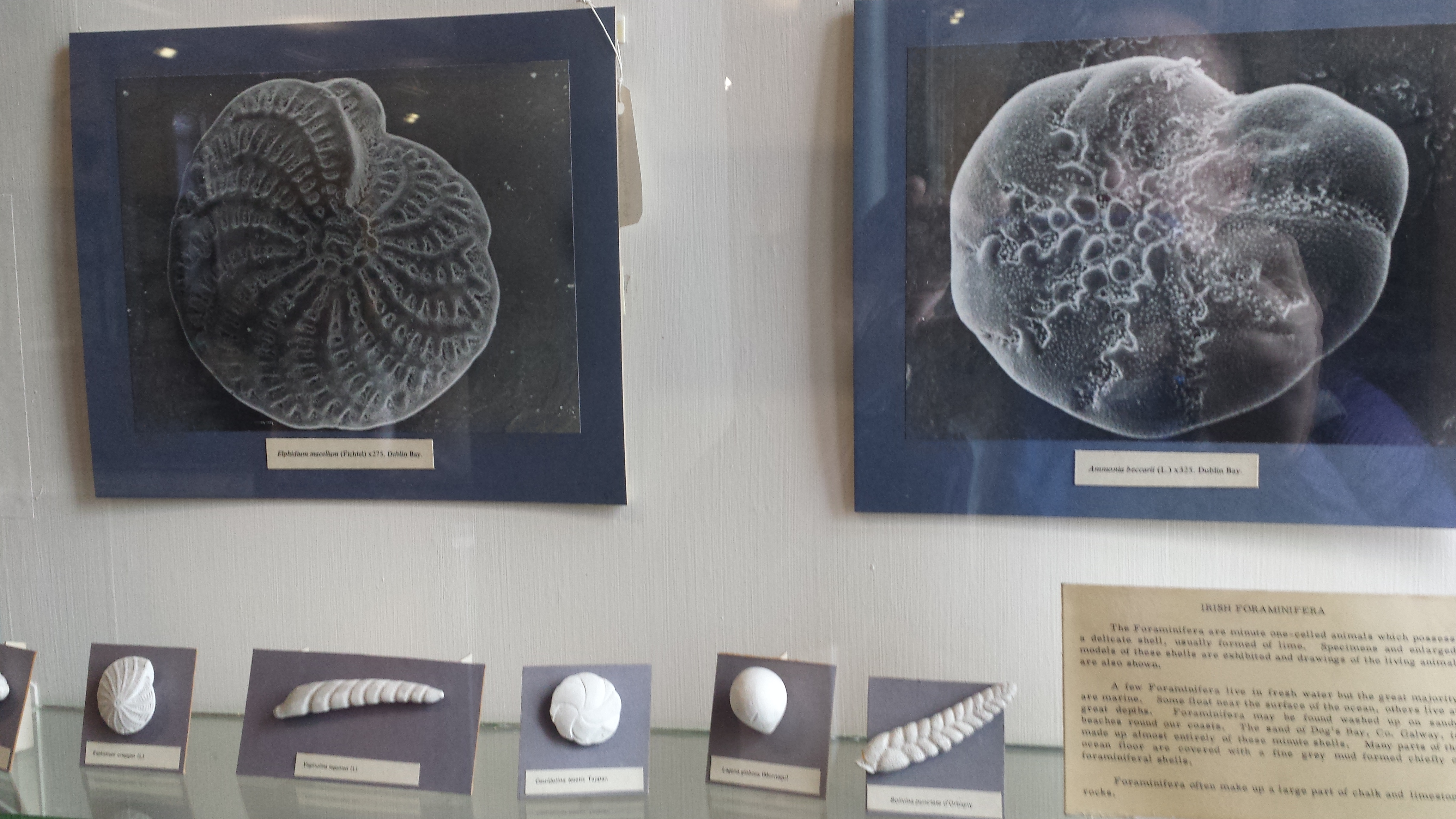
Who doesn’t like a good giant foramanifera image/model?
“That’s one bigass skate,” I murmured to myself.
“That’s one bigass halibut,” I quipped.
Tatty basking shark in entry hall.
Irish wolfhound, with a glass sculpture of its spine hanging near it, for some reason.
Stand back everyone! That beaver has a club!
Skull of a pilot whale/dolphin.
Nice anteater skeleton and skin.
Nice wombat skeleton and skin.
Sad display of a stuffed rhino with the horn removed, and signage explaining the problem of thefts of those horns from museum specimens of rhinos worldwide.
But then the stuffed animals started to get to me. Or maybe it was the hangover. Anyway, I saw this…
A proboscis monkey mother who seemed to be saying “Hey kid, you want this yummy fruit? Tough shit. I’m going to hold it over here, out of reach.” with a disturbing grimace. That got me thinking about facial expressions in stuffed museum specimens of mammals more, and I couldn’t help but anthropomorphize as I toured the rest of the collection, journeying deeper into surreality as I progressed. What follows could thus be employed as a study of the Tim-Burton-eseque grimaces of stuffed sloths. Click to emslothen.
Tree anteater has a go at the awkward expression game.
This completed my tour of the museum; there were 2 more floors of specimens but they were closed for, sigh, say it with me… health and safety reasons. Balconies from which toddlers or pensioners or drunken undergrads could accidentally catapult themselves to their messy demise upon the throngs of zoological specimens below. But the National Museum’s Archaeology collection was just around the block, so off I went, following whispered tales of bog bodies. There will be a nice, calm, pretty photo, then the bodies, so if peaty ~300 BCE cadavers are not your cup of boggy tea, you can depart this tour now and lose no respect.
Impressive entrance to the National Museum’s Archaeology building.
The bog bodies exhibit is called “Kingship and Sacrifice“. It is packed with cylindrical chambers that conceal, and present in a tomb-like enclosed setting, the partial bodies of people that were killed and then tossed in peat bogs as honoraria for the ascension of a new king. The peaty chemistry has preserved them for ~2300 years, but in a dessicated, contorted state. The preservation has imparted a mottled colouration and wrinkled texture not far off from a Twix chocolate bar’s. Researchers have studied the bejesus out of these bodies (including 3D medical imaging techniques) and found remarkable details including not just wounds and likely causes of death (axes, strangling, slit throats etc) but also clothing, diet, health and more.
Here they are; click to (wait for it)… emboggen:
Did you find the Celtic armband on one of them?
Finally (actually this happened first; my post is going back in time), I visited UCD’s zoology building for the PhD viva and saw a few cool specimens there, as follows:
Giant deer in UCD zoology building foyer, with a lovely Pleistocene landscape painted on the wall behind it.
Sika deer in an awkward posture (what is it supposed to be doing?) in Univ Coll Dublin zoology building’s foyer.
The pose of this ?baboon?mandrill struck me as very peculiar and menacing- reminiscent of a vampire bat’s pose.
A whole lotta chicken skeletons in a UCD teaching lab.
After the viva we went out for some nice Chinese food and passed some Dublin landmarks like this:
Trinity College entrance, I think.Former Irish Parliament; now the Bank of Ireland.
And we wandered into a very posh Irish pub called the Bank (on College Green), which displayed this interesting specimen, as well as some other features shown below:
Replica of illuminated 9th Century gospel manuscript “The Book of Kells”, with gorgeous Celtic art.
Vaults near toilets in the Bank pub. Almost as cool as having giant freezers down there.
Nice glass ceiling of the Bank pub.
And Irish pub means one big, delicious thing to me, which I will finish with here– much as I finished that night off:
This is a rant, but stick with me and this rant might have a silver lining toward the end, or at least a voice of reason within the roiling cloud of bitter blog-scowling. And there are pictures of cats.
My little tiger, Karmella.
Like probably almost anyone in the 21st century that does research in a field of biology, I grew up watching nature documentaries on TV, and that influenced me to become a scientist. Doubtless it remains a powerful influence on other people, despite the massive de-science-ification of certain cable channels ostensibly, or at least potentially, dedicated to communicating science and nature (Animal Planet and History Channel, we’re looking at you).
But now I’ve seen behind the curtain. There’s still magic to behold there (e.g. working with early episodes of Inside Nature’s Giants), to be sure. However, some of my experiences have led me to become increasingly discontented with the relationship between TV documentaries and scientists.
Black leopard with glowing motion capture markers and eyes; eerie image from our past studies.
Here’s a common flow of events, and how they sometimes veer into frustration or worse:
Once a month or so, especially concentrated around this time (May-June-ish), I get a call or email from a documentary producer or researcher who is fishing for expert advice as they build a proposal for a documentary. I’m always very happy to talk with them and direct them to the best researchers to speak to, or papers to read, or to aspects of my own work that fit in with their idea for a documentary. Sometimes their idea is a bad one and I’m not afraid to tell them that and try to steer them toward a better idea; on occasion that seems to work, but more often they have their plan already and are reluctant to deviate from it.
About 3/4 of the time, I either never again hear from these nascent documentaries or else hear back maybe one more time (even to meet for coffee or give them a tour of our campus)– presumably, the proposal fails at that stage as it doesn’t excite executives. I’ve easily grown to accept this status quo after some initial disappointments. Much like in science, some ideas just don’t pass the muster of “peer review”, and documentary makers are operating under more of a market economy than science tends to be. Sifting is inevitable, and the time I spend helping people at this stage is quite minimal, plus it’s fun to see the sausage being made in its earliest stages. All fair so far…?
Alexis and technician setting up gear for one of our past studies of how cats move.
The frustration naturally ramps up the more one invests in helping documentaries through their gestation period. I’m sure it’s very frustrating and stressful for TV makers, too, to spend days or months on a project and then have the rug pulled out from under them by those on high. Hopefully they are getting paid for their time; all I can speak to is my experience. My experience is that all this early input I regularly provide is pro bono.
I used to mention that my time is not cheap, and I had a policy (after a few disappointments and lost time) that I should get paid around £100/hour for my time, even at the early consulting stage. That fee went straight into my research funds to help send grad students to conferences or buy small consumables; it was definitely worth my effort and felt very fair. Since the 2008 economic downturn, I’ve rapidly abandoned that policy, because it seems clear to me that documentary makers of late tend to be working on more austere budgets. I’m sympathetic to that, and the payoff for a documentary that gets made with my input is often quite substantial in terms of personal satisfaction, PR/science communication, happy university/grant funders, etc. On rare occasions, I still do get paid for my time (albeit essentially never by the BBC); Inside Nature’s Giants was generous in that regard, for example.
How the leopard got glowy spots: motion capture markers from our past studies.
But at some point a line needs to be drawn, where the helpful relationship between scientists and documentary makers veers from mutualism into parasitism, or just careless disregard. I’ve been featured in roughly eight different TV documentaries since 2004, but there were almost as many (six or so) other documentary spots that went beyond the proposal stage into actual filming (easily 8+ hours of time) and never aired; either being cancelled entirely or having my scenes cut. All too frequently, I don’t hear about this cutting/cancellation until very late and after my inquiries like “Any news about the air date for your programme?”
Several times I’ve heard nothing at all from a documentary after filming, only to watch the programme and reach the end credits to find no sign of me or my team’s research (in one embarrassing case that really soured my attitude, the RVC had broadcast to the college to watch the show to see me in action, and upon watching we found out I was cut. Ouch!). At that point I really do wonder, is it all worth it? Hours or days invested in calls, emails, paperwork, travel, arranging and replicating an experiment, repeating filmed scenes and lines, working to TV producers’ scripts and demanding timetables. All that is totally worth it if the show gets made. But if the odds are ~60/40 or so that I get cut, I think I have cause to do more than shrug. The people I’ve worked with on documentaries can be wonderfully kind and full of thanks and other approbations, and they often impress me with their enthusiasm for the programme and their very hard, tenacious work making it all happen. It is jarring, then, to find out “Oh, you’ve been cut from the show, I’m so very sorry, the executives made that decision and it was a bitter pill for us to swallow, believe me– take care and I hope we can work together again.”
Above: Performance art illustrating what it’s like to have your science filmed for a documentary, then cut; graciously acted out by a cat (R.I.P.).
My aggravation has resurfaced after filming with BBC Horizon’s new documentary on “The Secret Life of the Cat,” airing right now. Alan Wilson’s team, from our lab, is featured prominently there, so that is fantastic for the Structure & Motion Lab (also check out his purrfectly timed Nature paper on cheetah agility vs speed, also from this week!). It’s hopefully going to be a nifty show; I’ve seen some of the behind-the-scenes stuff develop. (EDIT: I’ve seen it now and it was pretty good in terms of imagery and showing off Alan’s team’s technology, but the science was pretty weakly portrayed– even laypeople I’ve spoken to said “Cats avoid each other… duh!” and the evolutionary storytelling didn’t convince me as much as I’d like; it came across as arm-waving, which is a shame if the two featured cat researchers actually have built a scientifically reasonable case for it. One could not tell if the “changes” in 1 village’s cats evidenced by 1 week’s observation were happening within a cat’s lifetime or were truly evolutionary and recent. I don’t think I’ll watch the 2nd segment.)
I was filmed for a segment which probably would have been in the 2nd part of the show airing on Friday night, but I found out last week that it got cut with a week left before airing. I will be watching the show anyway, of course. I’m not that bitter. The segment featuring my team’s research was about how cats of different sizes do not do what other land mammals do, which is to straighten their legs as size increases across evolutionary spans. This helps support their body weight more effectively, but I explained in the filming segment that in cats, the lack of a change of posture in size may have other benefits despite the cost in weight support: it can make them more stealthy, more agile/maneuverable (segue to the cheetah paper cited above!), or even better able to negotiate rough terrain. Hence a domestic cat is in a biomechanical sense in many ways much more like a tiger than it should be for a “typical mammal”– an athlete, specialized for the hunt. And smaller cats are relatively much more athletic than bigger ones because they don’t suffer from the reduced ability to support body weight that bigger cats do. This may be, for example, why cheetahs are not very large compared with tigers or lions; they are at a “happy medium” size for agility and speed. But this all got cut, I am told.
Random cat that sidled up to us during some research into cat movements; so meta!
For my would-be-part in the show, we recreated experiments that I did with then-postdoc Alexis Wiktorowicz Conroy and others (a paper yet to be published, but hopefully coming very soon) that showed how cats large and small use such similar mechanisms in terms of postures as well as forces and moments (rotational forces). In these recreations, I got an RVC clinician to bring her cat Rocket (?IIRC) Ricochet over to be filmed walking over forceplates with high-speed video recording it. The cat didn’t do much for us; it probably found our huge lab a bit overwhelming; but it did give us at least one good video and force trace for the programme. Next we did the same thing with two tigers at Colchester Zoo, and got some excellent footage, including a tiger launching itself out of its indoor enclosure to come outside, while rapidly making a turn past the camera. The latter tiger “ate” (well, ripped to shreds, literally) the rubber mat that covered my pressure pad, too, which was mostly funny — and the film crew has reimbursed me for that as well as for the drive to/from the zoo. The filming experience was good; the people were nice; but the end result was a bummer.
Advantage of visiting Colchester Zoo for research: going behind the scenes and meeting a baby aardvark (that’s not a cat).
My segment, as far as I could tell, had cool footage and added a nice extra (if intellectual) context to the “secret life of cats” theme, so it’s a shame that it got cut. I heard that famed Toxoplasma-and-cat-behaviour researcher Prof. Joanne Webster‘s segment also got cut, so at least I’m in good company. I don’t have those cool videos of slo-mo cats and tigers with me now but will put them up early next week on my Youtube channel; stay tuned. They won’t ever show up on a documentary anyway; typically when footage gets cut it just vanishes into TV-land’s bowels.
So I’m not happy. Not at all. Bitter? Yeah, a bit. Spoiled brat scientist? I’d say that would be an overly cynical perspective on it. I do recognize that I am lucky that the research I do has a strong public appeal sometimes; many scientists will never be in a documentary or get much PR of any kind. But I think anyone has a right to examine their situation in life and ask, applying basic logic, whether it is fair treatment under the circumstances. Hence I have become disillusioned and angry about the relationship of documentary makers and scientists. Not just me, but us scientists in general. We’re unpaid actors playing sizeable roles and with major expertise. We give documentaries some sci-cred, too, simply by appearing onscreen with “Professor Snugglebunny from Smoochbridge University” in the caption. Supposedly, and often truly, we get good PR for it, when our segments don’t get cut or are not edited to obliterate the context or due credit. But it’s those latter instances that raise the question of fairness. If the segment gets cut, we simply have wasted our time. And to a busy scientist, that is like jabbing me with a hot poker.
Serenity now!
[Aside: I’m waiting to hear what has happened to another documentary I was filmed for, and again spent ~2 days on, Channel 5’s “Nature Shock: Giraffe Feast” which should be airing soon… no word yet if I’ve made the final cut but the show’s airing has been delayed; hopefully not a bad sign. I am crossing my fingers… it seemed like a great show with a cool idea, and my segment raised some fun anatomical and biomechanical issues about giraffes.]
I know I’m not alone. I’m going to end my rant and see what feedback it draws.
But don’t get me wrong— it’s not all sour grapes, not by any means. I’ve still had eight-ish pretty good TV documentary experiences (cough, Dino Gangs, cough!). I’ve had great experiences working with documentaries; indeed, Inside Nature’s Giants was one of the best experiences of my career to date. And I’m sure many other scientists have had positive experiences. In answer to my provocative “Why bother?” in the headline, there are plenty of good reasons to bother working with documentaries if you are a scientist whose research they want to feature… but only if you have some assurance that it will be worth your while, perhaps? How much of a gamble should we be bothering with? That brings me to my main point, a general query–
But what about the bad? And is it all worth it, in your views, given the risks of wasting time? Do we deserve some scientists’ bill-of-media-rights or something; a documentary-actor-scientists’ guild (90% joking here)? What should our rights be and should we push harder for them? Or do we just sit back and take the good with the bad, biting our lips? (I’m obviously not the type…)
I’d like to hear from not only the seasoned veterans who’ve experienced various ups and downs, but also from anyone that has views, anecdotes; whatever. I’m not aware of anyone collecting horror stories of documentary mishaps and mistreatments experienced by scientists, but that could start here. Please do share; even if you just got a call wondering if you’d want to help a documentary and then never heard back. Who knows where it would lead, but I think it’s helpful to bring these issues to the fore and discuss them openly.
Just a quickie here! I’m finishing a little sabbatical at Brown University and had a bit of downtime, then ran across this confusing image that seems to have loveable, sometimes-superhero Sesame Street character Grover in it, and also poses a tough but solveable Mystery CT Slice post! So go for it! Can you find Grover? (no points for that) and can you tell us (1) what the image is of (animal/species, region of anatomy, identifiable bits), and (2) what the heck is wrong with this image and why?
Stomach-Churning Rating: 1/10 unless you have bad childhood memories associated with Grover.
This is the mystery image below, not the Grover image above! You cheeky monkey.
No rhyming in your answers or you lose 10000000 points! Grover is grumpy today and hates rhymes. He had a bit too much Hefeweissen and polka music last night. Pity the poor creature.
Less words, more pictures in this post, and I’ll get the one lame cake joke out of the way early. I’ve nearly finished my research blitz through the postcranial material of the NHM-Tring’s osteological collection and have made some pit-stops for cake skulls now and then when I see one that pleases me. Now I shall present a survey of some of the species I’ve examined. I’ll proceed up from the base of the crown clade of living birds (Neornithes/Aves; the most recent common ancestor of living birds and all its descendants) and first take a tour of Palaeognathae; the ratites and kin; then move another step up into the Neognathae, first featuring the lineage featuring the ground fowl (Galliformes) and then the waterfowl (Anseriformes). If all this taxonomy and phylogeny is a bit much, check out this page for a brush-up on the bushy branches of bird biodiversity.
First, lots of bones of our cast of currasows, chachalacas, cassowaries and other kooky characters. And then, perhaps, a stop to the excessive alliteration. Finally, I will finish with some examples of species oddity (hat tip to Chris Hadfield).
Stomach-Churning Rating: 2/10- some bony pathologies but still just dry bones. Minimal cake jokes, and no filthy swearing this time.
BRING ON THE BONES:
My photographs are shown with kind permission from the Natural History Museum, London.
Exploded skull of an ostrich, Struthio camelus. This kind of careful preparation takes crazy skill, and creates a thing of rare beauty.
Imposing skull of a cassowary, Casuarius casuarius, with a rather worn head casque.
Mummified Owen’s Little Spotted Kiwi, Apteryx owenii. The feathers were still soft and fluffy, but I would not call this specimen cuddly.
Dorsal view of the back/hips of the Great Spotted Kiwi, Apteryx haasti. I like this photo and am not sure why. The symmetry and shading pleases me, I guess.
Front view of the back/hips of the Great Spotted Kiwi, Apteryx haasti, watching over my laptop and watching me while I write this blog on my laptop… so meta(ornithine)!
Wing of a kiwi, showing the fragile bones and feather attachments. “Apteryx” = “no wings”… well not quite. Click to emkiwi(?) so you can identify the individual bones, from the humerus right down to the fingers! I love this specimen.
The titanic left leg (in front view) of the Elephant Bird, Aepyornis maximus, from Madagascar, with a small moa nearby in left side view. There’s so much awesomeness about elephant birds I don’t know where to start, but this is one good place to do so.
The smaller end of the palaeognath scale: a mummified Undulated Tinamou, Crypturellus undulatus. Somehow the head got stuck into the abdominal cavity underneath the sternum, so this tinamou almost had its head up its arse. A tinamou with head in its proper position looks and sounds like this (video).
And now we take a left turn into the Galloanseres, most basal branch of the neognath birds, to see some of the neglected, strange early branches off from the “main line” that led to the modern diversity of ducks, geeses and swans (Anatinae, Anserinae).
Screamers (Anhimidae) are to Anseriformes as megapodes (see below; brush turkeys) are to Galliformes. By that I mean that both screamers and megapodes are very early branches off the main line of their respective lineages’ evolution, and both are quite strange when seen in that context… an unfair one, frankly; over-focused on the most familiar, “modern” or most speciose group. More about this issue further below.
This was my first hands-on experience with screamer anatomy; I was familiar from reading Tetrapod Zoology and other material about them. Check out the sound that gives them their name here! I’m now a big fan- they have so many strange features: oddly chunky but often very light bones, big feet with long toes, and then these switchblade-wrists, which would make Batman jealous:
Crested Screamer, Chauna torquata, showing the wicked spur (and smaller one) on the carpometacarpus.
Horned Screamer, Anhima cornuta; similar carpometacarpal spurs as in Chauna.
Torso of a screamer seen in top view. Nice narrow body, and no uncinate processes (spur-like bony struts that cross the ribs and act as levers for the muscles that move the ribcage during breathing)
The long, gracile, clawed toes of a screamer. Those toes, especially as they belong to an animal called a screamer, are spooky for me. Note also: very little toe-webbing for a “waterfowl.”
Not to be outdone, on the Galliformes side of Galloanserae, we have some funky headgear in the Maleo (a megapode bird/Megapodiidae; a very basal branch of “brush turkeys” and kin) and curassows (part of the Cracidae; odd South American birds whose males make booming sounds, presumably using their head-casques as resonating chambers?):
Australian brush-turkeys, Alectura lathami, at the Alma Park Zoo near Brisbane, Australia; they run wild there. Here they are doing what they are best known for: making a mound-like nest. We were doing kangaroo biomechanics experiments and they were everywhere. I was in awe to see such exotic (to me) birds; locals seemed not so enthused (the birds are loud and make a lot of mess).
Skull of Helmeted Curassow, Crax/Pauxi pauxi, showing that resonating chamber. Along with this boom-boom-room, the male uses a piece of food that he holds to draw in the female; if she takes it, then it’s sexy time.
Foot of a Siberian Black Grouse, Tetrao tetrix (nothing to do with a certain videogame), with and without flesh. Regard the broad, feathered feet, well insulated and with plenty of surface area for prancing around in the snow or moorlands. Tetrao engage in a cool display pattern called lekking, in which the males group together and show off to watching females.
A theme in the section above that is not to be missed is that there is some amazing disparity of anatomical forms in these basal lineages of poultry-relatives. Don’t dismiss the Galloanserae as just boring food-birds! Heaps of not-so-well-studied species exist here, surely with a treasure trove of cool neontological and evolutionary questions waiting for the right person to ask! Darwin’s chickens may get their share of neglect, but that pales in comparison to how little we understand about many basal Galloanserae.
What a lot of people think of as a “ground fowl” or galliform way of life is more of a way of life somewhat typical of the Phasanidae- chickens, pheasants and their familiar kin. Megapodes, curassows, guans, grouse and other Galliformes do not necessarily do things in the “typical” ground fowl way, much as the earlier branches of the Anseriformes don’t always look/act like “proper water fowl” (i.e. Anatidae). The phenomenon at play here is one of the great bugaboos in biology: essentialism— the often implicit misconception that variation away from some abstract ideal is negligible, uninteresting or just not conceivable due to mental blinders. When we say something like “the chicken is a fascinating species” we are sliding down the essentialistic slope. There is no “the chicken.” Not really. Oh dear, speaking of slippery slopes, I’d best stop here before I start talking about species concepts. And no one wants that to happen! Anyway, essentialism still pervades a lot of modern scientific thinking, and has its place as a conceptual crutch sometimes. But in biology, essentialism can be very insidious and misleading. It burrows in deep into the scientific mind and can be hard to root out. Unfortunately, it is entrenched in a lot of science education, as it makes things easier to teach if you sweep aside the exceptions to the essentialist “rules” in biology. I catch myself thinking in static, essentialist ways sometimes. The punishment is no cake for a week; so awful. 🙂
And speaking of “normal” or “typical,” morphology is of course often not that way even within a species, age class or gender. Pathology is a great example; by definition it is abnormal. It is a shattering of the “essence” of animals, brought on by some malady.
Next I’ve highlighted some of the amazing pathologies I’ve seen in the Tring skeletons. There have been so many I’ve been unable to keep track of them– some of these birds had the stuffing beaten out of them, and I’m not talking about Thanksgiving turkeys. Some were captive animals, in which the pathology might be blamed on living an inappropriate environment, but some were wild-caught — given the extreme pathologies, it’s a wonder those even survived to be found, but perhaps less a surprise that they were caught.
BONES GONE BONKERS:
View of left knee of a specimen of the Highland Guan, Penelopina nigra, showing some nasty osteoarthritis around the whole joint. Eew. A happier Guan sounds like this.
Femora and tibiotarsi of the Blue-throated Piping Guan, Aburria cumanensis. Amazing pathology involving the left femur (broken, rehealed) and tibiotarsus (secondary infection?). Interestingly, the non-fractured limb also showed some pathology, perhaps indicating general infection and/or arthritis in reaction to the severe damage to the other leg, or just increased load-bearing on that leg.
Little Chachalaca, Ortalis motmot, showing a broken and rehealed right femur and the tibiotarsus. As in the guan above, this animal was not walking for many weeks; its femur had snapped in two, but somehow melted back together. The tibiotarsus didn’t look too great, either; lumpy and bendy. In better times, the Chachalaca does the cha-cha like this.
These two specimens blew my mind. On the left is a normal Tetrao tetrix (Black Grouse); on the right is one hybridized with another (unknown) species.
In the picture above, what amazed me first was the very unusual flattened pelvis/synsacrum of Tetrao, which characteristically is light and wide. But in the hybrid this morphology was completely gone; the pelvis had a more standard “galliform” (read: Phasianid)-like shape, deeper and narrower and more solid in build. I am guessing that the hybrid was a cross with a pheasant like Phasianus itself, whose anatomy would be more like this. Somewhere in here there is a fantastic evo-devo/morphometrics project waiting to happen.
That’s my quick specimen-based tour of “basal birds”. Beyond these two clades of Palaeognathae and Galloanseres, there lies the forebidding territory of Neoaves: much of living avian diversity, and extremely contentious in its phylogenetic relationships. I’m tackling them next for my research on the evolution of the patella/kneecap. But first, I’ll be at the NHM-Tring today for a whirlwind tour through the respectably speciose “normal” Galloanseres clades of Phasianidae and Anserinae+Anatidae, so off I go! (It’s my wife’s birthday celebration, so cake may have to wait for later this time)
So what do you think? What’s your favourite neglected “primitive” bird group (more apropos: early branching avian lineage that may still be very specialized, rare and poorly understood), or cool factoid about palaeognaths and basal neognaths?
No quaggas were harmed during the writing of this post. Polly wanna quagga?