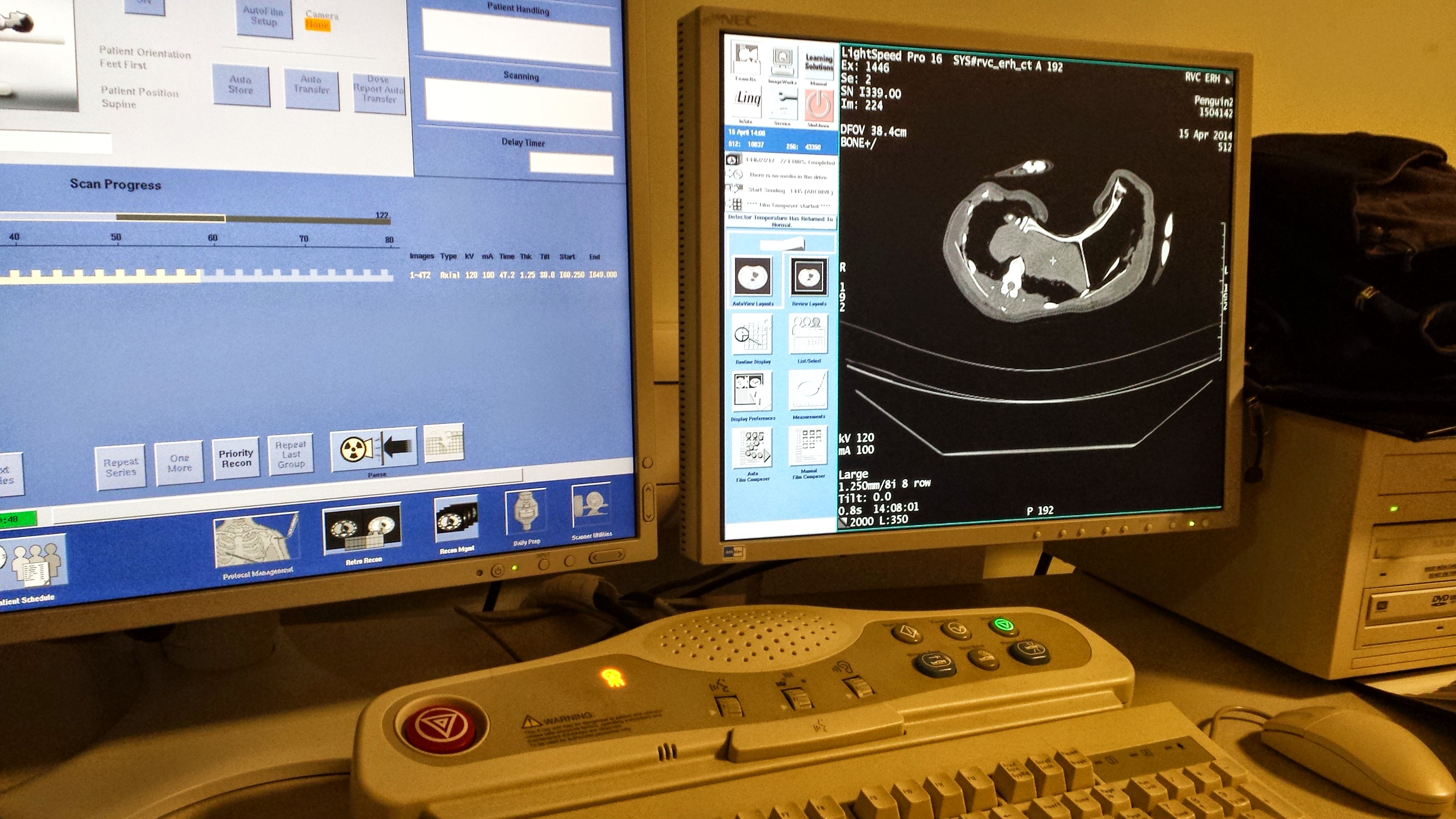
Sorry about the title. It’s the best I could do. In case you missed it on our Anatomy to You blog, we unleashed a hefty database of CT (and some MRI) scans of our frozen crocodile cadavers last week, for free public usage. In total, it’s about 34 individuals from 5 species, in 53 databases constituting around 26,000 individual DICOM file format slices of data. This page has a table of what the data/specimens are. I am writing this post to share some more images and ensure that word gets out. We’re thrilled to be able to finally release this first dataset. We have plans to let loose a LOT more such data in the future, for various organisms that we study.
Stomach-Churning Rating: 2/10- be glad that these data don’t come with an olfactory component, especially the five rotten, maggot-ridden Morelet’s croc specimens, which are among the stinkiest things I’ve dealt with.
Crocodiles are no strangers to this blog, of course, as these past links testify. Indeed, most of the crocodile images I’ve blogged with come from specimens that are in this scan dataset. We even released a “celebrity crocodile, “WCROC” or FNC7 in our dataset, which is the 3.7m long Nile croc from “Inside Nature’s Giants”. It broke our CT scanner back in ~2009 but we got the data, except for the torso, and we also got some MRI scans from it, so we’re chuffed.
Above: The only spectacled caiman (Caiman crocodilus); and indeed the only alligatorid; in our dataset. To watch for: stomach contents/gastroliths, and all the damn osteoderms that I did/didn’t segment in this quickly processed file. This specimen had its limbs dissected for one of our studies, so only the right limbs are visible.
Above: rotating Crocodylus moreletii (specimen FMC5 from our database) in a happy colour.
Sharing these kind of huge datasets isn’t so easy. Not only do few websites host them cheaply, and with reasonable file size limits, and limited headaches for what info you have to provide, and with some confidence that the websites/databases will still exist in 5-20 years, but also we were hesitant to release the dataset until we felt that it was nicely curated. Researchers can now visit my lab and study the skeletons (or in some cases, the still-frozen specimens) matched up with the scan data, and known body masses or other metadata. We’re not a museum with dedicated curatorial staff, so that was not trivial to reliably organize, and I still worry that somewhere in the dataset we mis-identified a specimen or something. But we’ve done our best, and I’m happy with that for now.
Above: rotating Osteolaemus tetraspis (specimen FDC2 from our database), which was obviously dissected a bit postmortem before we could scan it, but still shows some cool features like the extensive bony armour and the cute little doglike (to me, anyway) skull. I worked with these animals (live) a bit >10 years ago and came to love them. Compared to some other crocodiles we worked with, they had a pleasant demeanour. Like this guy:
Osteolaemus (resting) set up with motion capture markers for a yet-to-be-published gait study that we did in 2005 (ugh!). It wasn’t harmed by this.
Anyway, as a person who likes to maintain quality in the science we do, I also was hesitant to “just” release the DICOM file data rather than beautiful segmented 3D skeletal (or other tissue) geometry that is ready for 3D printing or animation or other uses, or interactive online tools like Sketchfab. Other labs (e.g. Witmerlab) do these kind of things better than we do and they inspire us to raise our game in the future, but I am sure that we will be forgiven for releasing big datasets without gorgeous visuals and more practical, processed files — this time. 🙂 We agree with many other scientists that sharing data is part of modern, responsible science– and it can be fun, too! Oddly enough, in this case we hadn’t used the CT/MRI data much for our own studies; most of the scans were never fully digitized. We just scan everything we get and figured it was time to share these scans.
Enjoy. If you do something cool with the data that we’ve made accessible, please let us know so we can spread the joy!
And if you’re a researcher headed to ICVM next week, I hope to see you there!
Last year we finally, after about 14 years of slow work, released our biomechanical model of an ostrich’s hindleg. We showed how it informed us about the potential leverages (moment arms; contributions to mechanical advantage of the joints) of all of the muscles. It was a satisfying moment, to understate it, to finally publish this work from my postdoc at Stanford. Today, we begin to deliver on that model’s promise. And it only took 4 years or so, roughly? The journal Royal Society Interface has published our study of how we used this musculoskeletal model to simulate walking and running dynamics. Those simulations join an intimidatingly broad and complex literature using similar models to study human (and some other primate) locomotion or other functions at the level of individual muscles (for whole limbs/bodies) in vast detail and growing rigor. I have Dr. Jeffery Rankin, a research fellow finishing up his post with me after ~6 years of hard work on many projects, to thank for driving this work forward, and Dr. Jonas Rubenson (now at PennState) for his patient collaboration that has continued since the early 2000’s.
Stomach-Churning Rating: 2/10; computer models of muscle actions. The underlying anatomical data are goopy, as prior ostrich-dissection-focused posts show!
Our model; in right side view (on the left) and frontal view (on the right), with muscles in red and the leg’s force as the blue arrow; frozen at the middle of a running step.
Simulations like these predict things that we can’t easily measure in living animals, such as how much force muscles and tendons generate, how quickly those tissues change length, how much mechanical energy they thus contribute to the joints, limbs and whole body, how much metabolic energy their actions cost, and much more. There are more ways to use these tools than I have space or time to explain, but simply put we forced our ostrich simulation to match experimental measurements of the motions and forces of a representative walking and running ostrich stride, from contact of one foot until the next time that foot hit the ground. It then used optimization methods (minimizing target criteria like muscle stress) to estimate how the muscles and tendons were used to generate those motions and forces. This is a ways ahead of some prior ostrich simulations such as this one that I recall from classes during my PhD studies.
Any modeller worth their salt knows that their models and simulations are wrong at some level. This is much as most science is “wrong”; i.e. a simplification of reality with some errors/noise introduced by assumptions, variation, methods and such. But generally these kinds of musculoskeletal dynamic simulations hold up pretty well against experimental data. A standard “validation” is to test how closely the simulations’ predictions of muscle activity match the “real” (measured in life, also with some uncertain error) activity of muscles. Science still lacks those data for ostriches, but fortunately measurements from other birds (by Steve Gatesy and colleagues) indicate that muscles tend to follow fairly conservative patterns. Grossly speaking, avian leg muscles tend to either be active mainly when the foot is on the ground (stance phase) or off the ground (swing phase). Some studies acknowledge that this is an oversimplification and other muscles do act across those two phases of a stride, either in multiple pulses or as “transitional” (stance-to-swing or swing-to-stance) switch-hitters in their activations. Our ostrich predictions matched the qualitative patterns for avian muscle activations measured to date, so that’s good. The results also reinforced the notion of transitional or multi-phasic muscle activation as still having some importance, which bears more study.
Yet what did the simulations with our ostrich model tell us that other ostrich experiments or other bird species didn’t? Three main things. First, we could calculate what the primary functions of muscles were; they can act as motors (generating energy), brakes (absorbing energy), springs (bouncing energy back and forth) or struts (just transmitting force). We could then sum up what whole muscle groups were doing overall. The image below shows how these broad functions of groups vary across the stance phase (swing phase is harder to condense here so I’ve left it out).
Positive work can speed things up; negative work can slow things down. Solid bars are running; striped bars are walking. (from our Fig. 13) You may need to click to em-broaden this image for the gory biomechanical details.
There’s a lot going on there but a few highlights from that plot are that the hip extensor (antigravity) muscles (biarticular hip/knee “hamstrings”) are acting like motors, the knee extensors (like our quadriceps) are mainly braking as in other animals and the ankle is fairly springy as its tendons (e.g. digital flexors; gastrocnemius) suggest. We often characterize birds as “knee-driven” but it’s more accurate biomechanically to say that their hips drive (power; i.e. act as motors) their motion, whereas their knees still act as brakes — in both cases as in many other land vertebrates. Thus in some ways bird legs don’t work so unusually. Birds like ostriches are, though, a little odd in how much they rely on their hamstring muscles to power locomotion (at the hip) rather than their caudofemoral muscles, which are reduced. Zooming in on some particular muscles such as parts of the hip or knee extensors, the functions sometimes weren’t as predictable as their similar anatomy might suggest. Some muscles had parts that turned on during swing phase and other parts used during stance phase. Neural control and mechanics can produce some unexpected patterns.
Second, we looked at one important methodological issue. Simulations of musculoskeletal dynamics can vary from simple static (assuming each instant of a motion is independent from the others; e.g. ignoring acceleration, inertia, tendon effects, etc.) to more complex grades of fully dynamic flavours (e.g. assuming rigid or flexible tendons). We looked across this spectrum of assumptions, for both walking and running gaits, with the expectation that more static assumptions would work less well (vs. more dynamic ones; by various criteria) for running vs. walking. This also showed us how much tendons influence our simulations’ estimates of muscle mechanics—a fully rigid tendon will make the muscle do all of the work (force times length change) whereas a flexible tendon can literally take up some (or even all) of that slack, allowing muscles to remain closer to their isometric (high force-generating, negligible length change) quasi-optimum.
Nicely, our predicted muscle functions weren’t influenced much by these methodological variations. However, static assumptions clearly were in some ways less appropriate for running than for walking, as were flexible tendons. Somewhat surprisingly, making the simulations more dynamic didn’t lower the levels of activation (and thus presumably the metabolic costs) of muscles, but actually raised those levels. There are good reasons why this might be realistic but it needs further study. It does muddy the waters for the issue of whether assuming that rapid locomotion can be modelled as static is a “bad” thing such as for estimating maximal speeds—yes, tendons can do more (elastic energy storage, etc.) if more dynamic models are used, but co-contraction of antagonistic muscles against each other also brings in some added costs and might lead to slower speed estimates. We’ll see in future work…
Finally, one often overlooked (sometimes even undiscussed!) aspect of these simulations is that they may silently add in extra forces to help muscles that are struggling to support and move their joints. The justification is typically that these extra “reserve actuators” are passive tissues, bony articular forces and other non-muscular interactions. We found that the hip joint muscles of ostriches were very weak at resisting abduction (drawing the thigh away from the body) and this needed resisting during the stance phase, so our simulations had very high reserve actuators switched on there. That fits the anatomy pretty well and needs more investigation.
Want to know more? Happily, not only is the paper free for anyone to view but so are all of the data including the models (modified slightly from our last paper’s). So, although the software (Opensim) isn’t ideal for 4-year-olds to play with (it is fancy engineering stuff), if you have the interest and dilligence it is there to play with and re-use and all that. But also watch this space for future developments, as there is more to happen with our steadily improving models of ostriches and other beasties. Anyway, while this paper is very technical and challenging to explain I am not too bashful to say it’s one of the finer papers from my career; a big stride forward from what we’ve done before. I have been looking forward for a long time to us getting this paper out.
P.S. Our peer reviewers were splendid- tough but constructive and fair. The paper got a lot better thanks to them.
Happy Darwin Day from the frozen tundra sunny but muddy, frosty lands of England! I bring you limb muscles as peace offerings on this auspicious day. Lots of limb muscles. And a new theme for future blog posts to follow up on: starting off my “Better Know A Muscle” (nod to Stephen Colbert; alternative link) series. My BKAM series intends to walk through the evolutionary history of the coolest (skeletal/striated) muscles. Chuck Darwin would not enjoy the inevitable blood in this photo-tour, but hopefully he’d like the evolution. Off we go, in search of better knowledge via an evolutionary perspective!
There is, inarguably, no cooler muscle than M. caudofemoralis longus, or CFL for short. It includes the largest limb muscles of any land animal, and it’s a strange muscle that confused anatomists for many years– was it a muscle of the body (an axial or “extrinsic” limb muscle, directly related to the segmented vertebral column) or of the limbs (an “abaxial” muscle, developing with the other limb muscles from specific regions of the paraxial mesoderm/myotome, not branching off from the axial muscles)? Developmental biologists and anatomists answered that conclusively over the past century: the CFL is a limb muscle, not some muscle that lost its way from the vertebral column and ended up stranded on the hindlimb.
The CFL is also a muscle that we know a fair amount about in terms of its fossil record and function, as you may know if you’re a dinosaur fan, and as I will quickly review later. We know enough about it that we can even dare to speculate if organisms on other planets would have it. Well, sort of…
Stomach-Churning Rating: 8/10. Lots of meaty, bloody, gooey goodness, on and on, for numerous species. This is an anatomy post for those with an appetite for raw morphology.
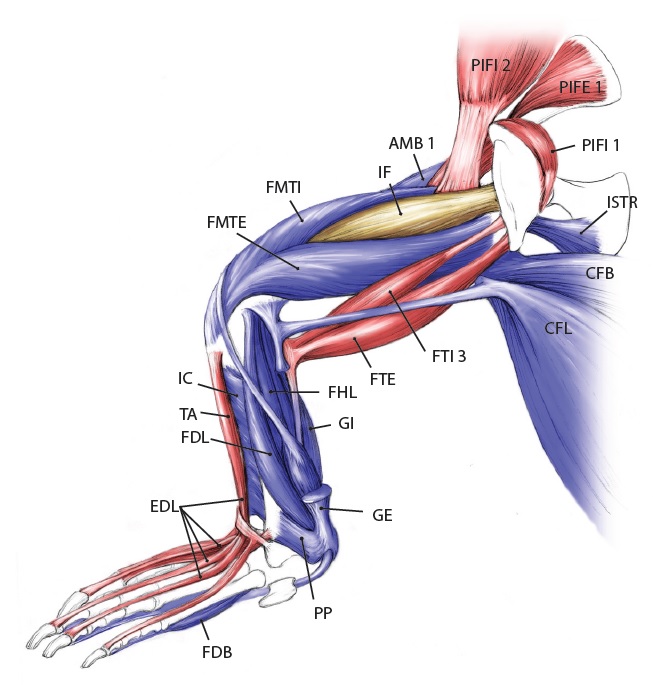
Let’s start from a strong (and non-gooey) vantage point, to which we shall return. The CFL in crocodiles and most other groups is (and long was) a large muscle extending from much of the front half or so of the tail to the back of the femur (thigh bone), as shown here:
Julia Molnar’s fabulous illustration of Alligator‘s limb muscles, from our 2014 paper in Journal of Anatomy. Note the CFL in blue at the bottom right.
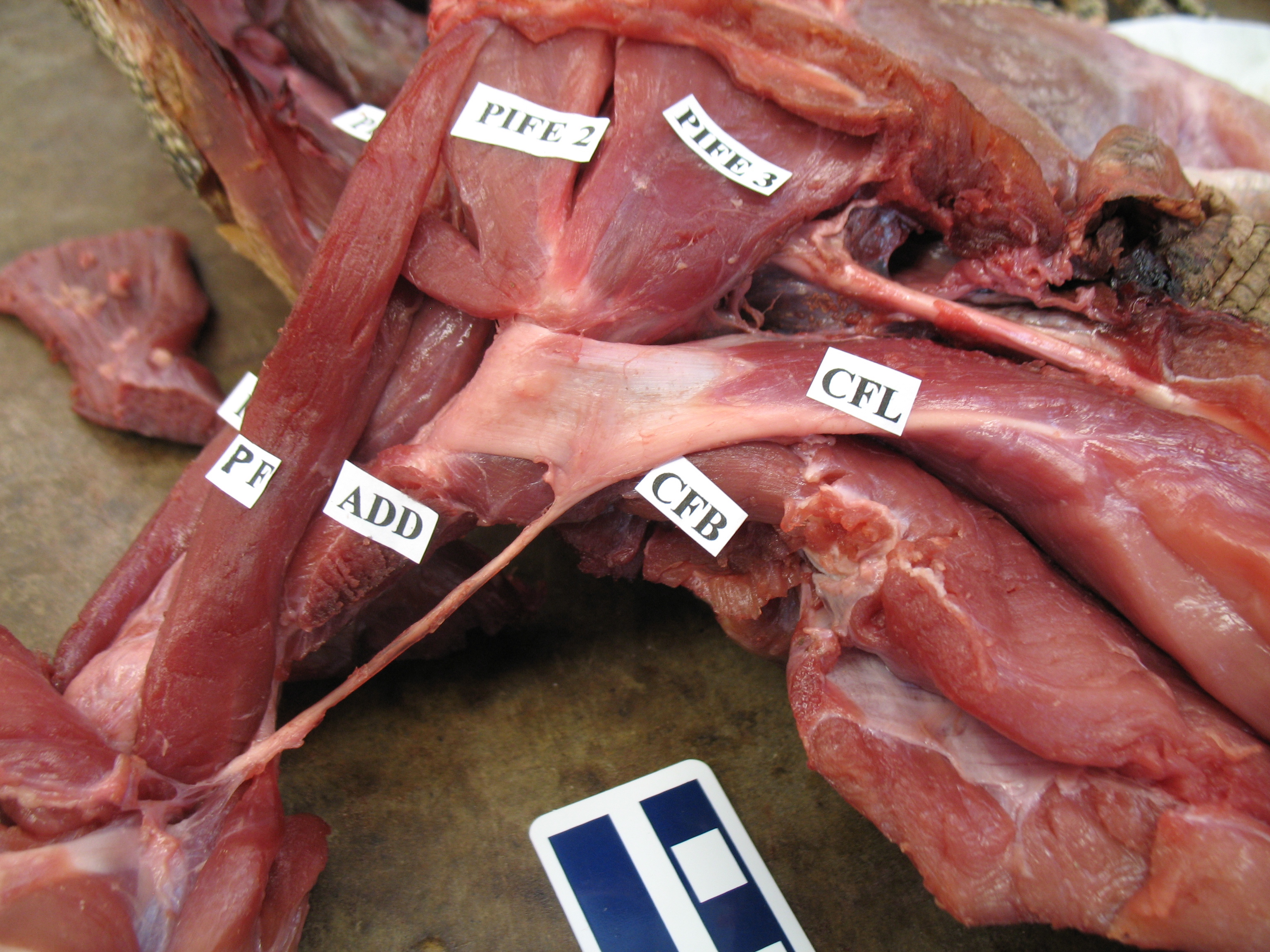
As the drawing shows, the CFL has a friend: the CFB. The CFB is a shorter, stumpier version of the CFL restricted to the tail’s base, near the hip. The “B” in its name means “brevis”, or runty. It gets much less respect than its friend the CFL. Pity the poor CFB.
But look closer at the CFL in the drawing above and you’ll see a thin blue tendon extending past the knee to the outer side of the lower leg. This is the famed(?) “tendon of Sutton“, or secondary tendon of the CFL. So the CFL has two insertions, one on the femur and one (indirectly) onto the shank. More about that later.
Together, we can talk about these two muscles (CFL and CFB) as the caudofemoralis (CF) group, and the name is nice because it describes how they run from the tail (“caudo”) to the femur (“femoralis”). Mammal anatomists were late to this party and gave mammal muscles stupidly unhelpful names like “gluteus” or “vastus” or “babalooey”. Thanks.
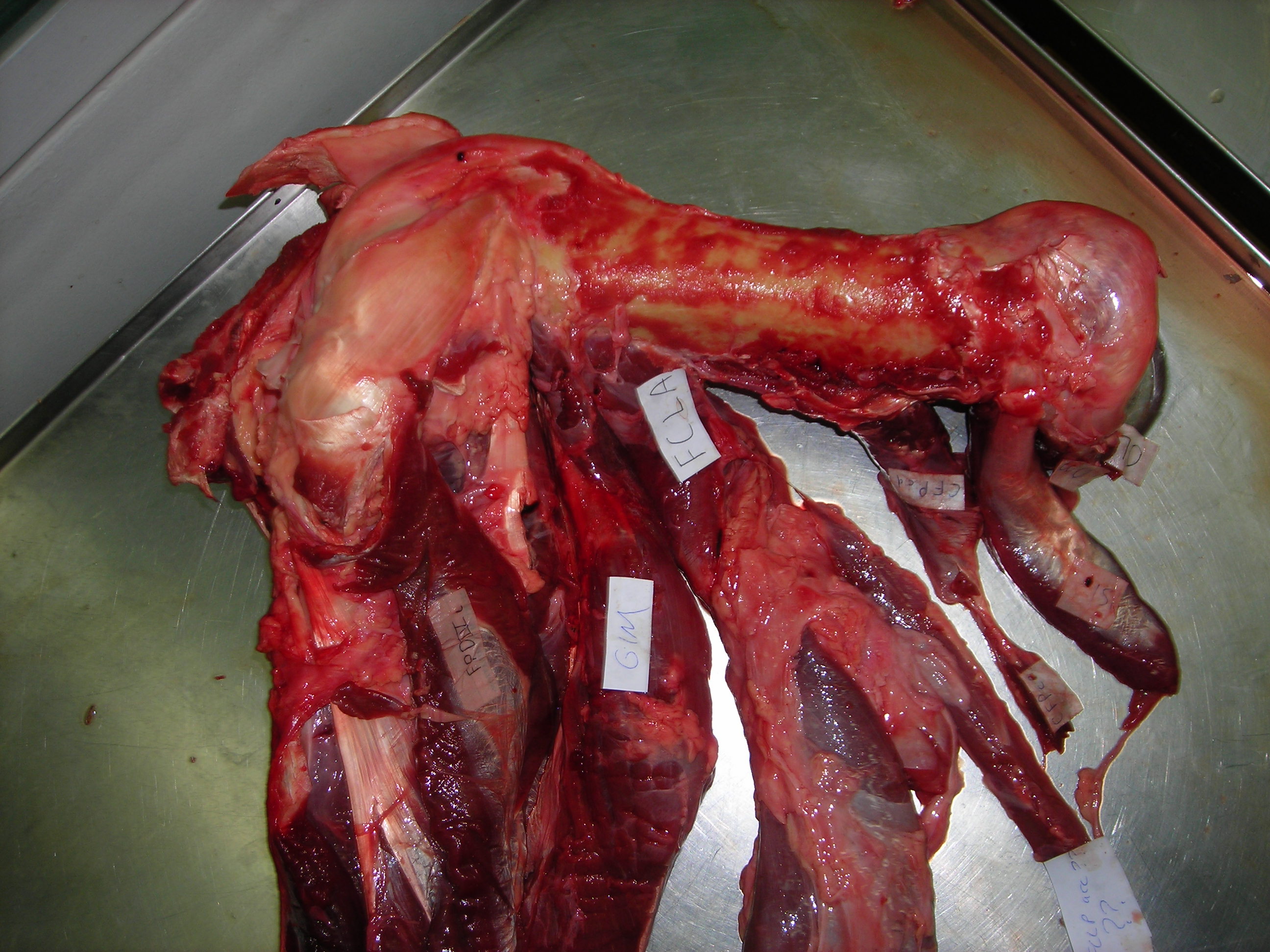
But enough abstract drawings, even if they rock, and enough nomenclature. Here is the whopping big CFL muscle of a real crocodile:
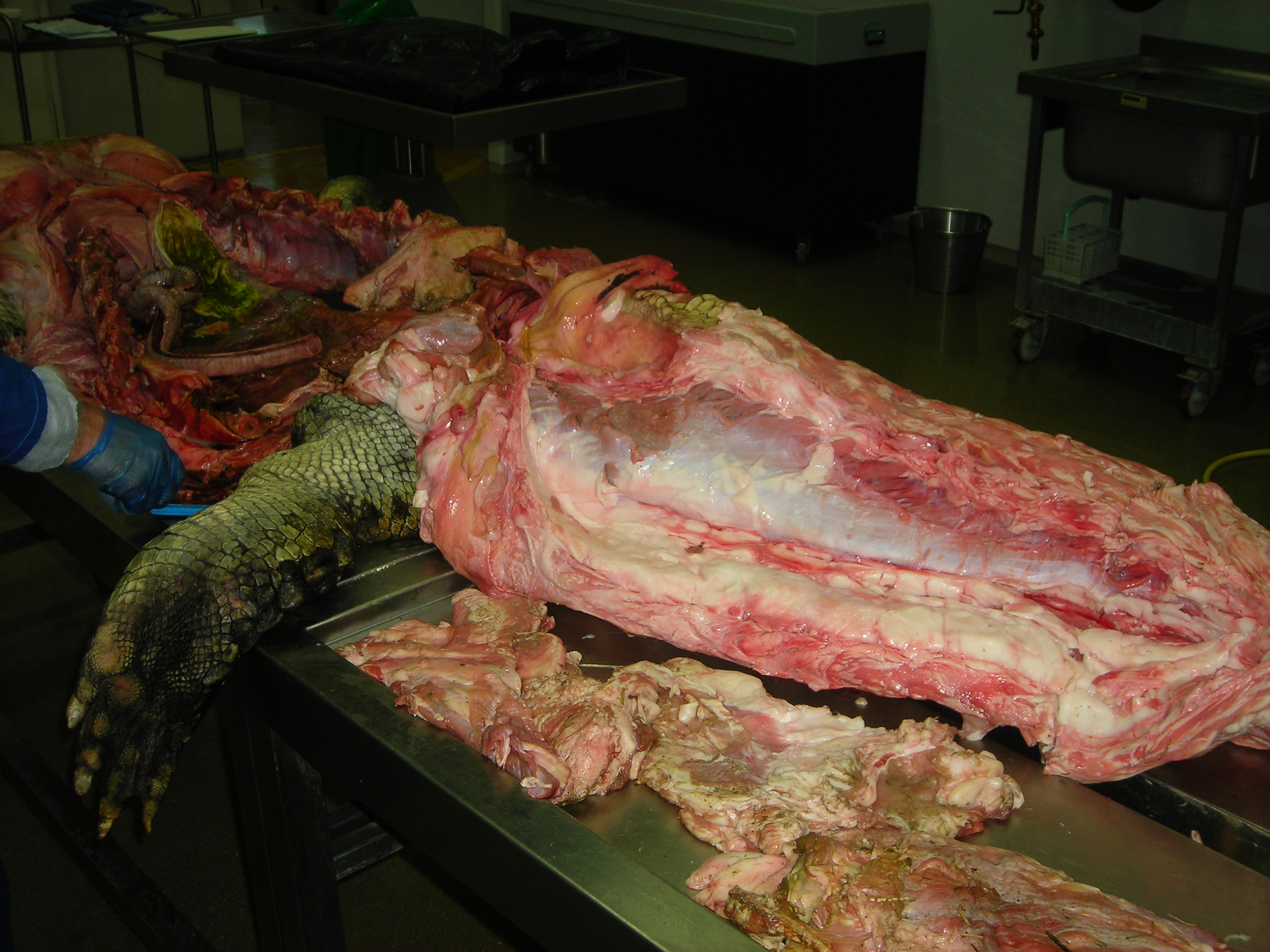
Huge Nile crocodile, but a relatively small CFL.
Bigger crocs have smaller legs and thus smaller leg muscles, relatively speaking. CFL at the top, curving to the left.
The giant Nile croc’s CFL muscle removed for measurements. 2.35 kg of muscle! Not shabby for a 278 kg animal.
However, maybe crocodile and other archosaur CFL muscles are not “average” for leggy vertebrates? We can’t tell unless we take an evolutionary tack to the question.
Where did the CFL come from, you may ask? Ahh, that is shrouded in the fin-limb transition‘s mysteries. Living amphibians such as salamanders have at least one CF muscle, so a clear predecessor to the CFL (and maybe CFB) was present before reptiles scampered onto the scene.
But going further back through the CF muscles’ history, into lobe-finned fish, becomes very hard because those fish (today) have so few fin muscles that, in our distant fishy ancestors, would have given rise eventually to the CF and other muscle groups. With many land animals having 30+ hindlimb muscles, and fish having 2-8 or so, there obviously was an increase in the number of muscles as limbs evolved from fins. And because a limb has to do lots of difficult three-dimensional things on land while coping with gravity, more muscles to enable that complex control surely were needed.
OK, so there were CF muscles early in tetrapod history, presumably, anchored on that big, round fleshy tail that they evolved from their thin, finned fishy one — but what happened next? Lizards give us some clues, and their CFL muscles aren’t all that different from crocodiles, so the CFL’s massive size and secondary “tendon of Sutton” seems to be a reptile thing, at least.
Courtesy of Emma Schachner, a large varanid lizard’s very freshly preserved CFL and other hindlimb muscles.
Courtesy of Emma Schachner, zoomed in on the tendons and insertions of the CFL muscle and others. Beautiful anatomy there!
Looking up at the belly of a basilisk lizard and its dissected right leg, with the end of the CFL labelled. It’s not ideally dissected here, but it is present.
An unspecified iguanid(?) lizard, probably a juvenile Iguana iguana, dissected to reveal its CFL muscle near its attachment to the femur. The muscle would extend further, about halfway down the tail, though.
Let’s return to crocodiles, for one because they are so flippin’ cool, and for another because they give a segue into archosaurs, especially dinosaurs, and thence birds:
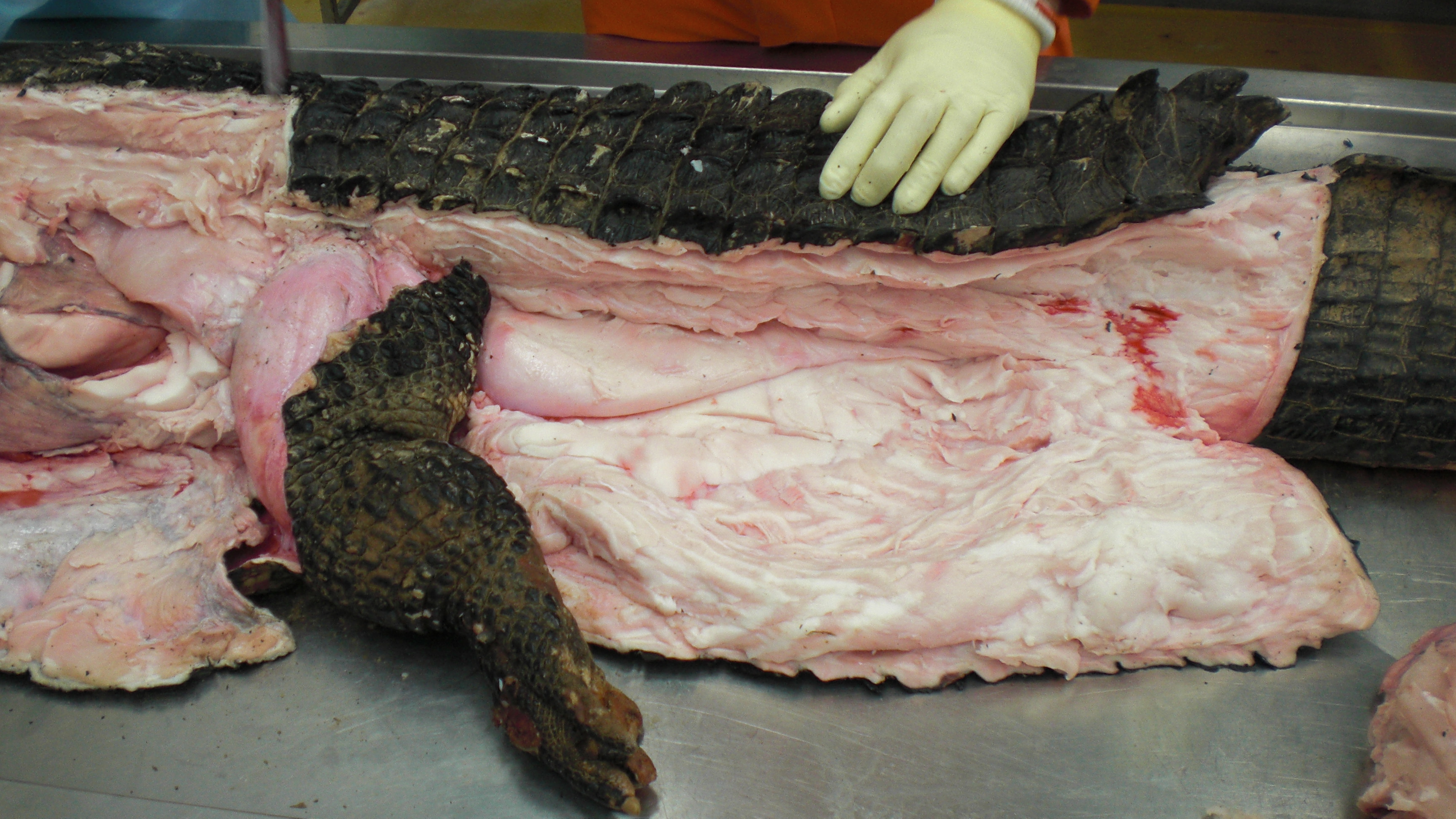
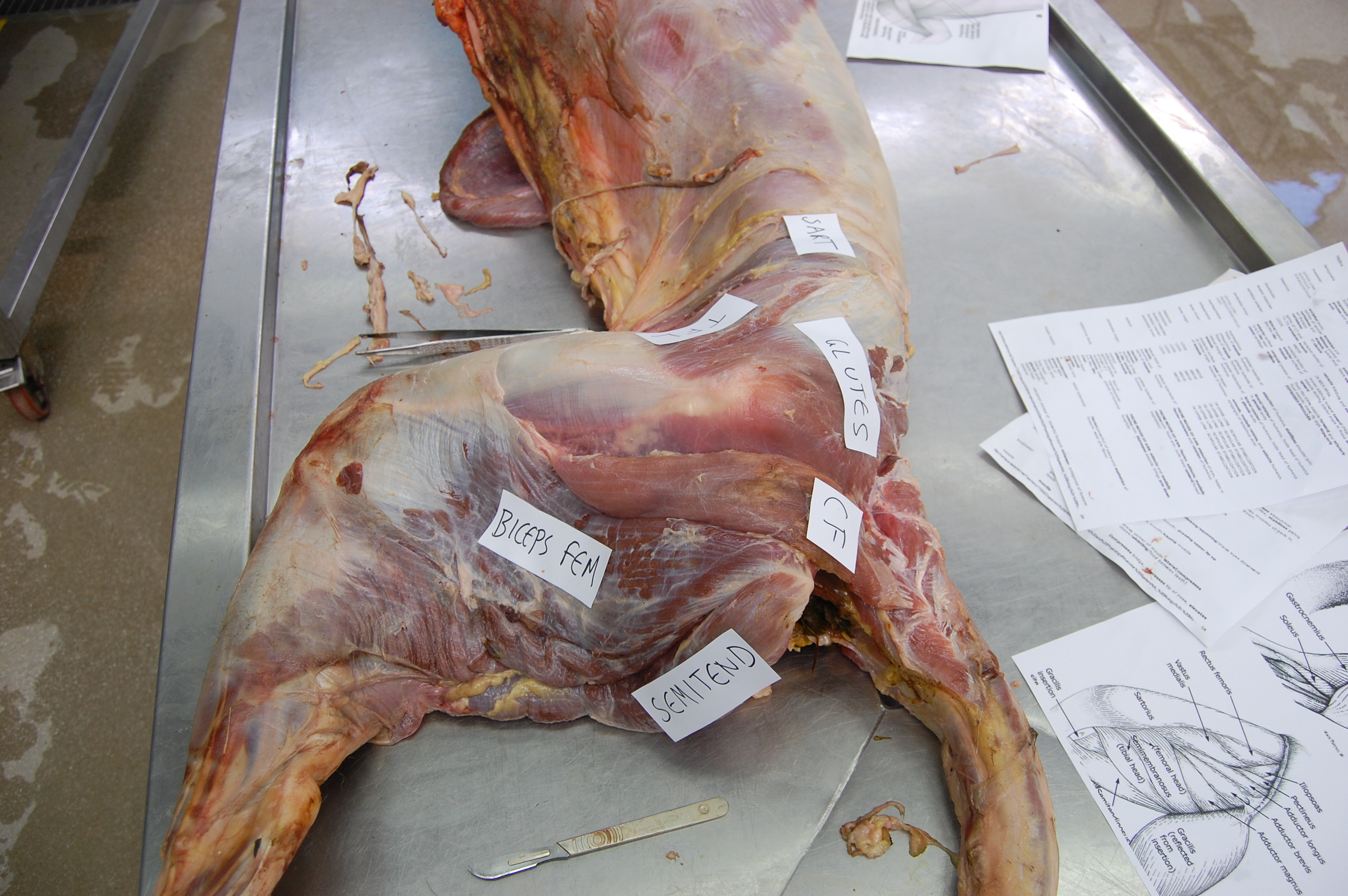
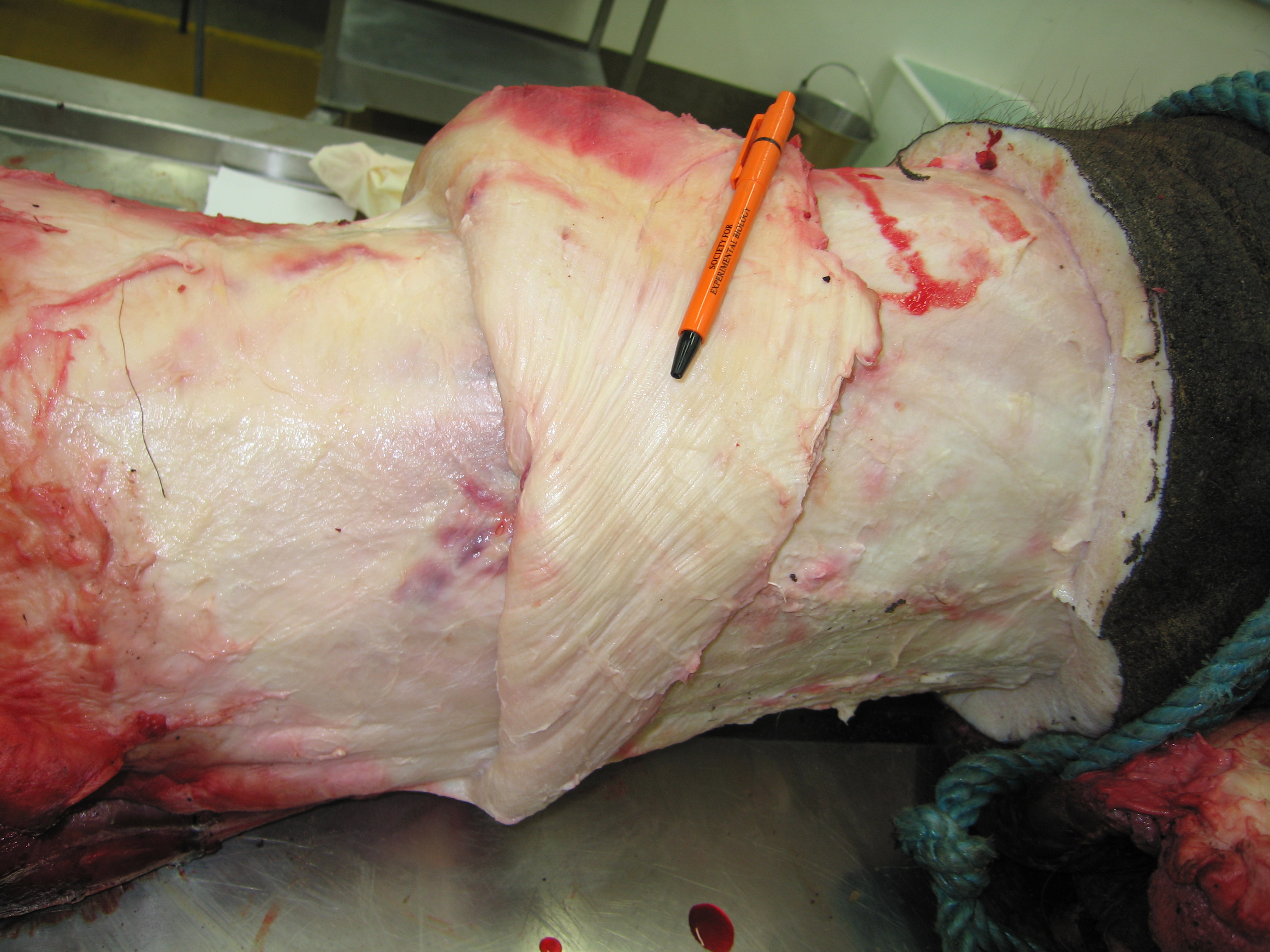
A moderate-sized (45kg) Nile crocodile with its CFL muscle proudly displayed. Note the healthy sheath of fat (cut here) around the CFL.
American alligator’s CFL dominates the photo [by Vivian Allen].
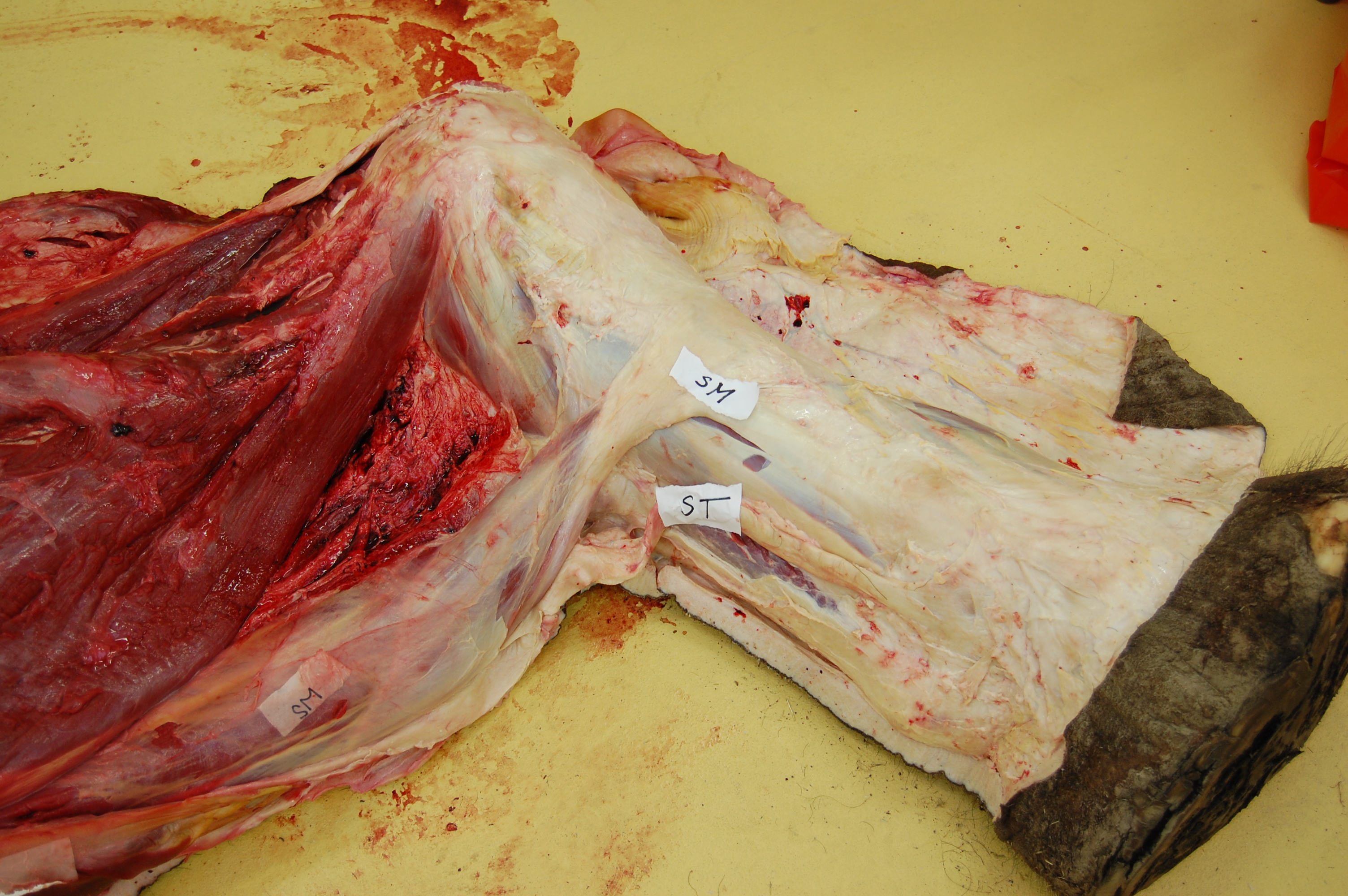
Black caiman, Melanosuchus, showing off its CFL muscle (pink “steak” in the middle of the tail near the leg), underneath all that dark armour and fatty superficial musculature.
A closer look at the black caiman’s thigh and CFL muscle.
Like I hinted above, crocodiles (and the anatomy of the CFL they share with lizards and some other tetrapods) open a window into the evolution of unusual tail-to-thigh muscles and locomotor behaviours in tetrapod vertebrates.
Thanks in large part to Steve Gatesy’s groundbreaking work in the 1990s on the CFL muscle, we understand now how it works in living reptiles like crocodiles. It mainly serves to retract the femur (extend the hip joint), drawing the leg backwards. This also helps support the weight of the animal while the foot is on the ground, and power the animal forwards. So we call the CFL a “stance phase muscle”, referring to how it mainly plays a role during ground contact and resisting gravity, rather than swinging the leg forwards (protracting the limb; i.e. as a “swing phase muscle”).
The “tendon of Sutton” probably helps to begin retracting the shank once the thigh has moved forward enough, facilitating the switch from stance to swing phase, but someone really needs to study that question more someday.
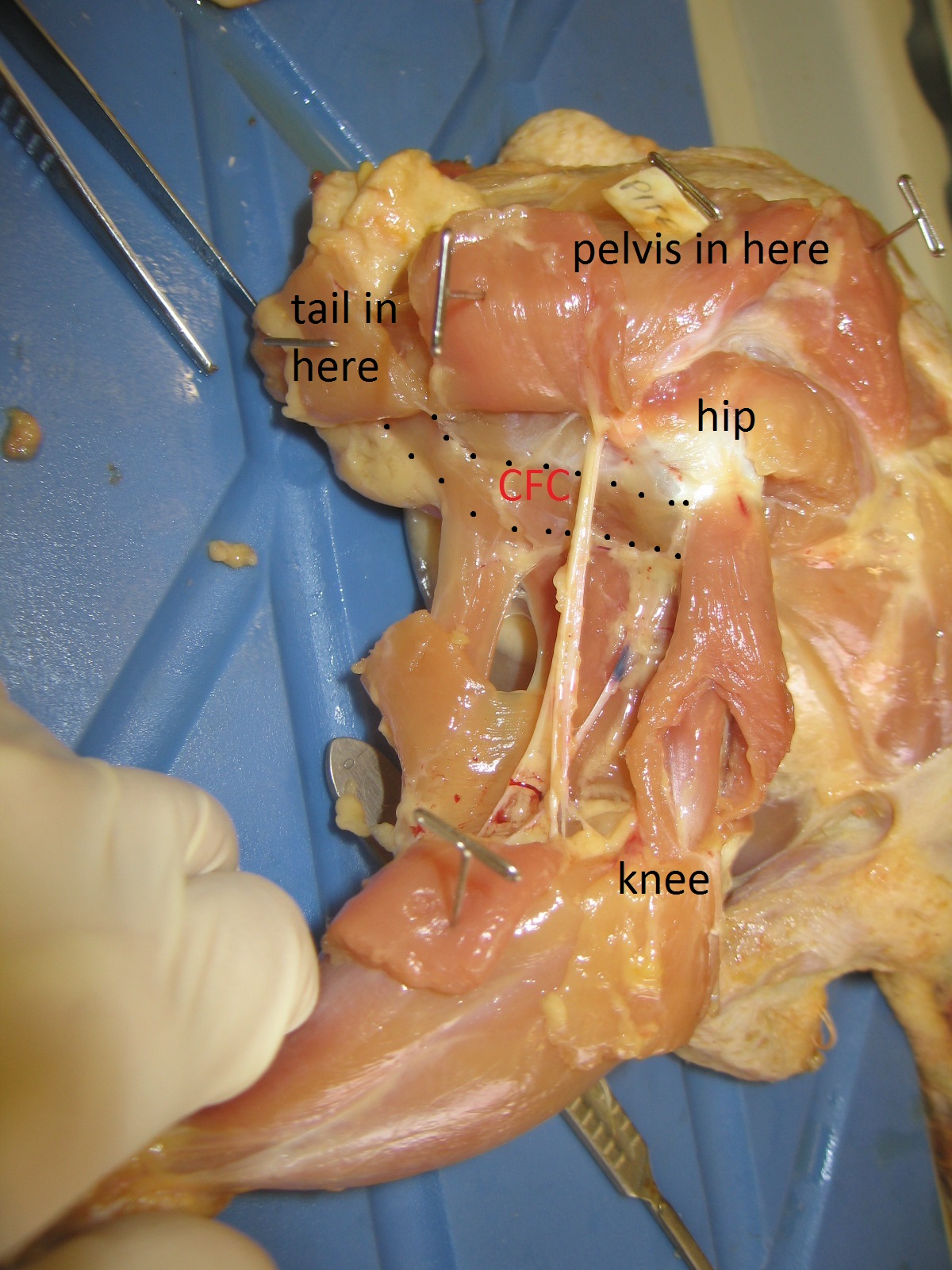
And thanks again to that same body of work by Gatesy (and some others too), we also understand how the CFL’s anatomy relates to the underlying anatomy of the skeleton. There is a large space for the CFL to originate from on the bottom of the tail vertebrae, and a honking big crest (the fourth trochanter) on the femur in most reptiles that serves as the major attachment point, from which the thin “tendon of Sutton” extends down past the knee.
Femur bones (left side; rear view) from an adult ostrich (left) and Nile crocodile (right). Appropriate scale bar is appropriate. The fourth trochanter for the CFL is visible in the crocodile almost midway down the femur. Little is left of it in the ostrich but there is a bumpy little muscle scar in almost the same region as the fourth trochanter, and this is where the same muscle (often called the CFC; but it is basically just a small CFL) attaches.
That relationship of the CFL’s muscular anatomy and the underlying skeleton’s anatomy helps us a lot! Now we can begin to look at extinct relatives of crocodiles; members of the archosaur group that includes dinosaurs (which today we consider to include birds, too), and things get even more interesting! The “tendon of Sutton”, hinted at by a “pendant” part of the fourth trochanter that points down toward the knee, seems to go away multiple times within dinosaurs. Bye bye! Then plenty more happens:
A large duckbill dinosaur’s left leg, with a red line drawn in showing roughly where the CFL would be running, to end up at the fourth trochanter. Many Mesozoic dinosaurs have skeletal anatomy that indicates a similar CFL muscle.
We can even go so far as to reconstruct the 3D anatomy of the CFL in a dinosaur such as T. rex (“Sue” specimen here; from Julia Molnar’s awesome illustration as part of our 2011 paper), with a fair degree of confidence. >180kg steak, anyone?
As we approach birds along the dinosaur lineage, the tail gets smaller and so does the fourth trochanter and thus so must the CFL muscle, until we’re left with just a little flap of muscle, at best. In concert, the hindlimbs get more crouched, the forelimbs get larger, flight evolves and voila! An explosion of modern bird species!
Left femur of an ostrich in side view (hip is toward the right side) showing many muscles that attach around the knee (on the left), then the thin strap of CF muscle (barely visible; 2nd from the right) clinging near the midshaft of the femur.
Another adult ostrich’s CF muscle complex, removed for study. Not enough ostrich myology for you yet? Plenty more in this old post!Or this one!Or this one… hey maybe I need to write less about ostriches? The CF muscle complex looks beefy but it’s no bigger than any other of the main hindlimb muscles, unlike the CFL in a crocodile or lizard, which puts everything else to shame!
STILL not enough ostrich for you yet? Take a tour of the major hindlimb muscles in this video:
And check out the limited mobility of the hip joint/femur here. No need for much femur motion when you’re not using your hip muscles as much to drive you forwards:
But I must move on… to the remainder of avian diversity! In just a few photos… Although the CF muscles are lost in numerous bird species, they tend to hang around and just remain a long, thin, unprepossessing muscle:
Chicken’s right leg in side view. CFC muscle (equivalent of CFL; the ancestral CFB is confusingly called the CFP in birds, as it entirely resides on the pelvis) outlined and labelled.
A jay (species? I forget) dissected to show some of the major leg muscles, including the CFL-equivalent muscle; again, smallish. [Photo by Vivian Allen]
Finally, what’s up with mammals‘ tail-to-thigh CF-y muscles? Not much. Again, as in birds: smaller tail and/or femur, smaller CF muscles. Mammals instead depend more on their hamstring and gluteal muscles to support and propel themselves forward.
But many mammals do still have something that is either called the M. caudofemoralis or is likely the same thing, albeit almost always fairly modest in size. This evolutionary reduction of the CF muscle along the mammal (synapsid) lineage hasn’t gotten nearly as much attention as that given to the dinosaur/bird lineage’s CFL. Somebody should give it a thoroughly modern phylogenetic what-for! Science the shit outta that caudofemoralis…
Yet, oddly, to give one apparent counter-example, cats (felids) have, probably secondarily, beefed up their CF muscle a bit:
Cats have a pretty large CF muscle in general, and this jaguar is no exception! But mammals still tend to have fairly wimpy tails and thus CF muscles, or they even lose them (e.g. us?). [photo by Andrew Cuff, I think]
In summary, here’s what happened (click to embeefen):
Better Know A Muscle: the evolution of M. caudofemoralis (longus).
I hope you enjoyed the first BKAM episode!
I am willing to hear requests for future ones… M. pectoralis (major/profundus) is a serious contender.
P.S. It was Freezermas this week! I forgot to mention that. But this post counts as my Freezermas post for 2016; it’s all I can manage. Old Freezermas posts are here.
Tonight is the debut of the ballyhooed BBC1 programme “Attenborough and the Giant Dinosaur“, featuring Sir David and The Titanosaur-With-No-Name, at 6:30pm. Furthermore, this week I presided over a showing of “T. rex Autopsy” to our RVC undergrad Zoological Society, with a very enjoyable Q&A afterwards. So it seemed timely for me to do a post about a theme that links these two documentaries that I helped with, my own research, and science communication and palaeontological research more generally. But first let’s get this out of the way:
It was great. I could gush more but that’s not what this post is about.
Stomach-Churning Rating: ~7/10; mainly the elephant leg dissection that’s not far below, which is a bit messy.
For the titanosaur documentary with Sir David, and the incomparable Ben Garrod as well, we had an old elephant “friend” of mine (subject of many biomechanics studies we’d done) walk across a pressure pad to demonstrate how the elephant locomotion works and some of the basic similarities with how a giant sauropod dinosaur might walk:
A broader feature of that documentary, which elephants are linked into, is how we can use the skeleton to reconstruct some general aspects of the soft tissue anatomy, and thereby the physiology or even behaviour, of a giant titanosaur. Which brings me to this post’s subject: We dig up dinosaurs all the time, but what about digging into dinosaurs and using what’s preserved to reconstruct what isn’t?
Some of the “G-suit” compressive stocking that the fascia wrapped around elephant, and other large mammals, creates, and the underlying, interwoven muscles and tendons (hindlimb of a young Asian elephant that sadly died in captivity). Did some larger dinosaurs have something like this? I’d expect so.
Another view, more superficially, of that G-suit/stocking under the thick, tight skin of an elephant’s leg. You’ll hear more about this in the Attenborough show…
Once the documentary airs, I may be able to share some more images from it showing what they did for the titanosaur, but this BBC photo gives a good idea. Here, blood vessels and other tissues surrounding the skeleton. How would a titanosaur pump blood around its body? A good question.
I wanted to focus more on the end product; the imagery or even physical object; and how it conveys what we think we know about dinosaur anatomy. I’ll do that via examples, using photos of dinosaur anatomy that I’ve collected over the years from museums or other media. There won’t be any profound points or long musings; it’s mainly a photo blog:
What your (inner?) child most needs is a dinosaur to dissect yourself! Why not a T. rex toy like this?
For the price (~$30 in USA), the 4D Vision dinosaurs deliver a pretty good bargain, and the anatomy is satisfactory. I’ve been collecting this series. I could quibble, but hey: it’s a dinosaur you get to build/dissect yourself, and with many major organs in reasonable positions! Not so easy to put/keep together, but it’s fine. I would not pay a ton for it, though.
Poster of Velociraptor’s anatomy I’ve had since grad school, adorning my office. For ~1996 (no feathers; “zombie hands“), it’s damn good, mostly… Closer views below (sorry, photo quality is crap– taking photos of wall poster turned out to be harder than I expected! Bad lighting.) :
Closeup of the leg muscles- hey, not bad! Pretty much the right muscles in the right places more or less, and plausible proportions. No air sacs in the torso, but again, this is mid-1990’s science shown. BUT…
I was happy with this poster until I got it home from the western-USA museum I bought it at and read this final bit of text… Oh, America! You silly place. (And unfortunately, these dinosaurs are not from the very end of the Cretaceous anyway, so “global catastrophe” is not implicated.)
Looking down onto the opened torso of the Barcelona ornithomimid. Strikingly realistic! Breastbone, heart, liver, intestines; not unreasonable positions and sizes. No feathers, but again hey– this was made in the earlier days.
Albertosaurus from the Drexel Academy of Sciences. I forget where I got this pic but I like the display– this is an impressive full-scale physical model. The transition from skeleton-only on the left to skinned in the middle to fully-fleshed and popping out atcha on the right is clever.
?T. rex? leg, photo that I took ages ago as a PhD student, if memory serves. Can anyone remind me where this was? California Academy of Sciences? I am embarrassed that I cannot recall. I remember geeking out about it. It has a toy-ish look, but I reckon they had to build this to withstand kids touching it.
Perhaps the best dino-dissection example I’ve seen in a museum: the AMNH’s sauropod Mamenchisaurus with internal organs and their functions projected onto it, in the “World’s Largest Dinosaurs” exhibit. Bravo! I stood and watched it for quite a while.
This is far from comprehensive– just several kinds of imagery that I mostly like. There’s the tension between showing too much realism, which science simply can’t back up, and being too cartoonish, losing the viewer’s immersion in the time-travelling fantasy. I do, however, like other kinds of more abstract, schematic depictions of dinosaur anatomy that simplify the details to focus on the basics of what organs should have been where and how they may have worked, such as this depiction from T. rex Autopsy, which also took the other extreme favouring ultra-realism (but with physical models, not so much with the CGI):
Air flow through a T. rex: simplified but clear. CGI used to explain, not abused. The real air sac anatomy would be too complex to show. You may see something similar with the titanosaur show.
That’s enough for now. I’ve stuck with relatively recent examples; of course in my particular field I also think back to Romer’s wonderful 1920’s drawings, which I covered in this post.
So, blog readers, help me out here: what examples of dinosaur internal, squishy anatomy from museums, documentaries or other not-entirely-done-by-nitpicky-scientists venues do you like, or not like so much? What works for you, or at least is memorable in some way?
Seeking adaptations for running and swimming in the vertebral columns of ancient crocs
A guest post by Dr. Julia Molnar, Howard University, USA (this comes from Julia’s PhD research at RVC with John & colleagues)
Recently, John and I with colleagues Stephanie Pierce, Bhart-Anjan Bhullar, and Alan Turner described morphological and functional changes in the vertebral column with increasing aquatic adaptation in crocodylomorphs (Royal Society Open Science, doi 10.1098/rsos.150439). Our results shed light upon key aspects of the evolutionary history of these under-appreciated archosaurs.
Stomach-Churning Rating: 5/10; a juicy croc torso in one small photo but that’s all.
Phylogenetic relationships of the three crocodylomorph groups in the study and our functional hypotheses about their vertebrae. * Image credits: Hesperosuchus by Smokeybjb, Suchodus by Dmitry Bogdanov (vectorized by T. Michael Keesey) http://creativecommons.org/licenses/by-sa/3.0
As fascinating as modern crocodiles might be, in many ways they are overshadowed by their extinct, Mesozoic cousins and ancestors. The Triassic, Jurassic, and early Cretaceous periods saw the small, fast, hyper-carnivorous “sphenosuchians,” the giant, flippered marine thalattosuchians, and various oddballs like the duck-billed Anatosuchus and the aptly named Armadillosuchus. As palaeontologists/biomechanists, we looked at this wide variety of ecological specializations in those species, the Crocodylomorpha, and wanted to know, how did they do it?
Of course, we weren’t the first scientists to wonder about the locomotion of crocodylomorphs, but we did have some new tools in our toolbox; specifically, a couple of micro-CT scanners and some sophisticated imaging software. We took CT and micro-CT scans of five fossil crocodylomorphs: two presumably terrestrial early crocodylomorphs (Terrestrisuchus and Protosuchus), three aquatic thalattosuchians (Pelagosaurus, Steneosaurus, and Metriorhynchus) and a semi-aquatic modern crocodile (Crocodylus niloticus). Since we’re still stuck on vertebrae (see, e.g., here; and also here), we digitally separated out the vertebrae to make 3D models of individual joints and took measurements from each vertebra. Finally, we manipulated the virtual joint models to find out how far they could move before the bones bumped into each other or the joints came apart (osteological range of motion, or RoM).
Our methods: get fossil (NHMUK), scan fossil, make virtual fossil and play with it.
Above: Video of a single virtual inter-vertebral joint from the trunk of Pelagosaurus typus (NHMUK) showing maximum osteological range of motion in the lateral direction (video). Note the very un-modern-croc-like flat surfaces of the vertebral bodies! (modern crocs have a ball-and-socket spinal joint with the socket on the front end)
While this was a lot of fun, what we really wanted to find out was whether, as crocodylomorphs became specialized for different types of locomotion, the shapes of their vertebrae changed similarly to those of mammalian lineages. For example, many terrestrial mammals have a lumbar region that is very flexible dorsoventrally to allow up-and-down movements during bounding and galloping. Did fast-running crocodylomorphs have similar dorsoventral flexibility? And did fast-swimming aquatic crocodylomorphs evolve a stiffer vertebral column like that of whales and dolphins?
Above: Video of how we modelled and took measurements from the early crocodylomorph Terrestrisuchus gracilis (NHMUK).
Our first results were puzzling. The Nile croc had greater RoM in side-to-side motions, which makes sense because crocodiles mostly use more sprawling postures and are semi-aquatic, using quite a bit of side-to-side motions in life. The part that didn’t make sense was that we found pretty much the same thing in all of the fossil crocodylomorphs, including the presumably very terrestrial Terrestrisuchus and Protosuchus. With their long limbs and hinge-like joints, these two are unlikely to have been sprawlers or swimmers!
So we started looking for other parts of the croc that might affect RoM. The obvious candidate was osteoderms, the bony scales that cover the back. We went back to John’s Freezer and got out a nice frozen crocodile to measure the stiffness of its trunk and found that, sure enough, it was a lot stiffer and less mobile without the osteoderms. If the fairly flexible arrangement of osteoderms in crocodiles had this effect on stiffness, it seemed likely that (as previous authors have suggested; Eberhard Frey and Steve Salisbury being foremost amongst them) the rigid, interlocking osteoderms running from head to tail in early crocodylomorphs would really have put the brakes on their ability to move their trunk in certain ways.
Testing the stiffness of (Nile) crocodile trunks to learn the effects of osteoderms, skin, muscles, and ribs. We hung metric weights from the middle of the trunk and measured how much it flexed (Ɵ), then removed bits and repeated. Click to em-croccen.
Another cool thing we found was new evidence of convergent evolution to aquatic lifestyles in the spines of thalattosuchians. The more basal thalattosuchians, thought to have been near-shore predators, had stiffness and RoM patterns similar to Crocodylus. But Metriorhynchus, which probably was very good at chasing down fast fish in the open ocean, seems to have had greater stiffness. (The stiffness estimates come from morphometrics and are based on modern crocodiles; see here again, or just read the paper already!) A stiff vertebral column can be useful for a swimmer because it increases the body’s natural frequency of oscillation, and faster oscillation means faster swimming (think tuna, not eel). The same thing seems to have happened in other secondarily aquatic vertebrate lineages such as whales, ichthyosaurs, and mosasaurs.
So, our results were a mixed bag of adaptations particular to crocs and ones that seem like general vertebrate swimming specializations. Crocodylomorphs are important because they are the only group of large vertebrates other than mammals that has secondarily aquatic members and has living members with a reasonably similar body plan, allowing us to test hypotheses in ways that would arguably be impossible for, say, non-avian dinosaurs and birds. The take-home message: crocodylomorphs A) are awesome, and B) can teach us a lot about how vertebrates adapt to different modes of life.
Well, that was an eventful week for me, although today’s post will focus on one event: the debut of the film Jurassic World. Briefly though, the awesome “T. rex Autopsy” documentary debuted (I was going to post more about it but all I’d have left to say is that I was very pleased with the result), I also showed up briefly in “Top 10 Biggest Beasts Ever” talking about the giant rhinocerotoid Paraceratherium and the stresses on its feet, our paper on ostrich musculoskeletal modelling was published (more in a future post) after ~12 years of me diddling around with it, and much more happened. Then to cap it all off, very shortly after I hit the “publish” button on my last post, I had four tonic clonic seizures in a row and spent a hazy night in the hospital, then the past week recovering from the damage. Nothing like another near-death (no exaggeration there, I’m afraid) experience to cap off an exciting week. But strangely, what I feel more interested in talking about is, like I said, Jurassic World, but this is not a review, as you will see here.
Stomach-Churning Rating: 0/10; just SPOILERS if you haven’t seen the film yet!
I guess I have to give a brief review of the film and say that I was entertained, to a degree, but it was not a great piece of film-making. It was a far cry from the original but then so were the sequels, and maybe it was better than them. The mosa-star was the most novel, memorable bit. I didn’t care for the Indominus villain, but then when you bring genetic engineering into a film like this, you’ve basically thrown out the rulebook and can make your dinosaurs as magical as you want; we’re already in “X-Men” territory here and almost in “Pacific Rim”-land.
Chris Pratt has signed on for at least another sequel to Jurassic World and the ending of the film already started that ball rolling. So I find it fun to speculate wildly, and certainly incorrectly, on what the sequel might do. What does the Jurassic future have in store?
First of all, who survived to re-appear in the next film? We’re left with the Bryce Dallas Howard character, who probably will return with Pratt to further develop their rather uninteresting social/romantic dynamic, rather than start afresh with someone else. The kids of course survived, as always, and as always they won’t return, as that’s not interesting and they didn’t have much to do except scream and (highly implausibly) hotwire an old car. Much as I’d like the parents from my hometown of Madison, WI to return, they won’t either for the same reasons. But we really only need Pratt and his high-heeled sweetie for the next film. Everyone else memorable(?) seems to have died, although it would be wonderful to bring Goldblum back for some smarmy wit (please!).
Second of all, the next film can’t be set in Jurassic World. There’s not much left to do there (JW already spent much of its time hearkening back to JP), and there’s no way the park would re-open. We need something new. I think by now we’re (very) tired of characters running around islands full of dinosaurs and the Blackfish parallel was milked dry in the latest movie. We need to spend a film with the dinosaurs amongst humanity (as Lost World briefly did), and much as I’d love to see the crazy drug lord/kidnapping plot happen, it won’t. But JW did set one thing up that has to happen now in its sequel: the paramilitary role of engineered, trained dinosaurs. We now know they can sort of train their dinosaurs and they can forge them to be anything they want to in terms of geno/phenotypes. They’ll learn from some mistakes of JW and engineer (or already did by the end of JW, at some remote site) some more compliant, deadlier animals, having largely given up on the public exhibition angle. The naked raptors and T. rex probably have to re-appear (sigh), but enough already of the giant uber-theropods like Spinosaurus and Indominus. The latter was already enough of a reprise of the former (plus psychic talents and chameleon powers etc.). Something truly novel is needed.
Unless they engineer a hyper-aggressive, intelligent sauropod or ceratopsian, which would admittedly be neat, I have this prediction (which is probably wrong but hey!): they have shown they can hybridize anything. There must be fewer and fewer “normal” (1990s…) dinosaurs now in the JW universe. So the next big step, which someone in the JW universe surely would do, is to hybridize dinosaurs and humans. Maybe some raptor-human hybrids, maybe also saving a tyranno-human hybrid for a surprise late appearance. But this is the sensible next step because it allows them to play with the (tired) Frankenstein monster trope but also touch on the hot topic of human cloning and human GM.
And by unleashing dino-human hybrids, or at least some freaky clicker-trained and engineered super-dinos, they could also explore the military theme, which the JW universe still hasn’t delved into much. What if those hyper-smart, deadly hybrid dinos, led by Pratt and Howard’s expert training, were used to combat an ISIS-analogue terrorist threat? Dino-Avengers in the badlands of Afghanistan or Iraq? Too predictable perhaps, but that’s a film that the public will want to see. Yeah there’s plenty of stupid there, but there’s no turning back– each film ups the ante, as JW ironically reminds us several times. We’re already in firmly in stupid-land, and the science has largely advanced to the point of magic. My idea is too uncomfortably close to the abandoned John Sayles plot, true. Darn. And (groan) kids have to be involved in some way to make it a family film so it rakes in the $$$ again, so either they get caught in the middle of the paramilitary mess or they are the ones that have to be saved… or the hybrid dinos are cute-ish kids themselves that Pratt and Howard must manage… (shades of Teenage Mutant Ninja Turtles/Kick-Ass?) I’ve found that more fun to think about than dwelling on the flaws of the movie, which is frankly too easy.
(Another good theme that Vivian Allen suggested to me would be climate change and invasive species—i.e. planet warms, dinos are already loose and go feral in waterlogged Central America, ecological disaster is looming and something must be done to round up the dinos… could work in some other bits like ecotourists or drug runners?)
That’s as far as my wildly speculative ruminating has taken me, but I wanted to turn it over to you, Freezerinos. If you were to make the next film (will it be “Jurassic World 2”? “Jurassic Army”? or as I’ve proffered in the post’s title, “Jurassic Future”?), what would it be (A) in your ideal world where you call all the shots (yes, lots of colourful feathery dinos, I know), vs. (B) in a more likely (less daring, more Hollywood) reality, along the lines of what I’ve tried to do here? (but I surely will be wrong, although we’ll see in 2-5 years!)
In early 2011, I got a fun email from a producer at National Geographic TV about a new project they were planning, which involved dissecting a full-scale model of a Tyrannosaurus rex in a documentary to be called “T. rex Autopsy.” Things fell silent for some months, then I got another email saying they were moving forward, then things fell silent again. Three and a half years later I got another email, this time from a producer at Impossible Factual films (working with NatGeo), saying that the show was finally moving forward for real. (This sort of thing is normal for documentaries; time scales can be long and unpredictable, or very fast-paced) This email invited me to be a primary scientific consultant in the design of the creature and show. Of course, anatomical dissection and T. rex are what I’m about as a scientist; two of my major research areas; so bringing them together was like a dream come true and I leapt into that dream with enthusiasm.
(Meanwhile, circa 2010-11, another TV channel filmed me for a different programme in which a whole, fresh-ish T. rex was found weathering out of an Alaskan cliffside and scientists had ~2 days to study it before it fell into an abyss– it’s probably best that that show never happened… there were fundamental flaws.)
Stomach-Churning Rating: 0/10. Merciful. No gory images here, just text descriptions and 2 glamour shots, for various reasons. The TV show is not for small children, though. I am guessing that the final programme will be about a 7/10 SCR because of gooey, seemingly rancid, but rubbery (so it doesn’t look overly real, but still looks great) dinosaur vital organs. For more pictures, see the links to tweets, trailers and news stories below.
I introduced T. rex Autopsy to you in the previous post, I’ve been tweeting and retweeting extensively since then, and one of my later posts will be a “postmortem” of the show, which airs June 7 worldwide. My feeling is that, if what I’ve seen so far is indicative of the whole show, it will be a landmark moment in palaeontological documentary history. T. rex Autopsy fuses the best aspects of “Inside Nature’s Giants” with “Walking With Dinosaurs”, and without “Alien Autopsy” pseudoscience. Indeed, it seems to be a very science-based documentary (once you get past the requisite conceit that scientists could actually find a very fresh T. rex body– that’s the only sci-fi bit of the show, quickly dispensed with!). T. rex Autopsy is packed with evidence-based palaeobiology, and has consistently been so since I first spoke to producers, which was a great comfort to me.
This post is about my role in the show, my perspective on it, and an attempt at a spoiler-free prelude. I’m willing to go out on a limb a bit and urge people to watch it, because I’m already proud of what was attempted in the show– it was a bold vision by NatGeo and laborious execution by everyone involved. I especially want to give a big shout of respect out to creature designers Crawley Creatures (led by Jez Gibson-Harris [interview here], who helped create Jabba the Hutt and the Dark Crystal beings, among others). Around 14 people on Jez’s team worked full time for ~4 months to make the T. rex. The designers based the proportions on the Field Museum’s scans of “Sue”, which I helped them get access to (I’d used them for our PLOS ONE paper in 2011). That, and numerous comments on their draft dinosaur’s body proportions and limb positions (e.g. avoiding “bunny hands“), was some of my first major involvement in the programme.
Edwina revealed! (photo credit: National Geographic Channels/Stuart Freedman)
Over 200 emails (I was curious; I counted them!) and a bunch of phone calls and 7 months later, my input on the T. rex Autopsy film shoot and production was finishing. Just last week, I sent what supposedly was my last email of input on the show, about predatory habits (NOT the dumb scavenger debate we’re all tired of; more about ambush vs. pursuit habits). I’d spent many hours going over drafts of T. rex‘s anatomy and function and behaviour from head to tail with the superb Impossible Factual film production team (mainly Assistant Producer Cressida Kinnear). Very often, to their credit, they’d already done a lot of literature searching and speaking with key experts on dinosaur jaws or brains or breathing, so I just had to check the fine details, but in some cases I had to recommend experts to speak to and/or do my own sleuthing and educate myself about aspects of T. rex biology I’d never pondered much.
For example, how big was T. rex‘s heart? I’d been asked the same question about sauropods lately for another show so I had references and an Excel spreadsheet ready to go, and plugged in some values, but the estimates I got seemed too small relative to the thoracic cavity (mediastinum if you must). I had some interesting back-and-forth discussions with the producers and we settled on one size that seemed “right”. No one that I knew of had tried to scientifically estimate the size of a T. rex‘s heart, probably because there hadn’t been a good reason to try. Sauropods get all the dino-love in regards to blood pressure issues and heart size, for good reasons- for them, it should have been a serious biomechanical challenge to pump blood up the long neck to the brain. For an elephant-sized T. rex, it doesn’t boggle the scientific mind so much that blood pressure wasn’t such a major evolutionary design constraint. See the show and find out more about what the intrepid team of dissectors found…
Did T. rex have feathers? This was important to get right, I felt, and not just show T. rex as a leathery or scaly beast, which is outdated. As I put it, it’s more speculative to show T. rex without any feathery thingies than to show it with some. We passed around draft images and thoughts and agreed on a slightly fuzzy, bristly body, especially in some regions of the head/neck, arms and tail tip. I encouraged the design team to go for more colour (I wrote to the designers “Skin colouring: go nuts! Feathery things should be colourful. Big animals tend to be more drab in colour but that doesn’t mean a boring grey/green, and certainly there should be some regional patterning. I like the idea of there being brightly coloured areas on the face”). We can be confident that dinosaurs could see colour like most land animals (except many mammals!) can. All of this is pretty familiar to palaeo-artists and fans of modern dinosaur reconstructions, so I won’t belabour it more. I’m glad that much of this made it into the final design. It’s not your overly familiar Jurassic Park T. rex.
Cheeks, eye pupils, brain/senses, how big a mouthful of meat it could swallow, furcula (wishbone), gastralia (belly ribs- I gave a lot of detailed criticism here), reproductive anatomy and biology, eggs, body fat, growth, air sacs, stomach, and excretory system, among other things: we covered them all in discussing the dino’s design, and I learned a lot along the way.
A memorable part of my discussions with the designers, in early March, was about the intestines and cloaca (rear-end opening): they initially put the cloaca too far forward on the body, I got them to move it backward, then I later realized in a panic that, making a neophyte error, I’d missed a key anatomical feature in the hips that clearly would put the “vent” even further backward, so I send them a hasty email apologizing that I’d missed this and urging that they fix their graphics and animations. I felt bad about this as it was late in the design phase and I’m sure I stressed out the team to make this change, but I thought it would be embarrassing to get the position of that hole wrong. Yet it was also funny to me to be scrutinizing where the “poop hole” of a dinosaur should go, and worrying so much about getting it right… my scatological sense of humour was in overdrive. By the middle of March they had this detail right. Phew!
There is another dinosaur that makes an appearance in the documentary but I don’t want to spoil it. Suffice it to say that one dinosaur from another time period and continent was initially chosen, and I (echoed by Dave Hone, I know) urged them not to do that, choosing a more appropriate Hell Creek Formation dinosaur. Phew! Perhaps more about that later.
Finally, of course we talked about legs and muscles and locomotion. I was filmed at the RVC discussing this, and it looks like it will be a cool segment, including an explanation of how the bones reveal the anatomy of the soft tissues of limbs and other parts of the body (i.e. bread-and-butter from my PhD thesis work). I hope that makes the final cut! (Edit: I’m told it has; yay!) There may even be footage of me dissecting a chicken and talking about enlarged and reduced leg muscles in birds, in any “making of” side-programme.
But I was not one of the four people doing the T. rex dissections in the show. That arduous job (2 looooooong days of filming!) fell to vet Luke Gamble and palaeontologists Tori Herridge, Steve Brusatte and Matthew Mossbrucker, with a crew of assistants including some from Crawley Creatures. The clever idea the producers had, as they explained it to me, was to keep my and others’ scientific input on the show’s design separate from the dissectors’ knowledge, so that when the dissection team arrived and cut into the dinosaur, they’d be discovering things without much advance inside knowledge of what to expect to find. We’ll see how that worked when the programme airs– I’ve only seen the trailer and behind-the-scenes footage, as well as the first day of filming. Scientists like me aren’t Shakespearean actors so it’s hard to act surprised when you sort of know what’s coming and have to redo takes of that same surprise. But if you come to T. rex Autopsy expecting Oscar-worthy theatrics, you’ve missed the point. 🙂
A taxi drove me to Pinewood Studios (west of London; site of filming many blockbusters) on a Sunday morning in late April. I walked into the giant studio where a 12+ meter long T. rex carcass lay in dramatic lighting. Cue the freezing of my giant grin in place and my eyes wide open. I was stunned! It was gorgeous, and the scale of the carcass left me gobsmacked. I’d only seen various incarnations of it during the design phase, from wire mesh scale models to clay sculptures to full-on foam casts and CGI representations; and all of these just as digital files emailed to me. But to see “Edwina,” as she was called, in the pseudo-flesh, was a moment I may never forget. Emailed JPGs definitely didn’t prepare me for that visual splendour. Crawley’s team were still inserting some of the last ~20,000 goose feathers as bristles into the hide, one by one…
I was at Pinewood to spend a day hobnobbing with VIPs and international press visitors as a “tour guide” to the Edwina autopsy event, and then for a day to watch the initial half of filming with the press in a room overlooking the studio. I got excellent hospitality, was called the “on-screen talent” in documents, which felt really weird to me (I’d never been called that in >10 shows before), and I spent a lot of time explaining the show and dinosaur science to that receptive, inquisitive audience. And gawking at the unfolding spectacle before and during filming. And cracking jokes with journalists during long breaks between actual filming of the documentary. It was a surreal, awesome experience and I loved it. (And, as I’ve insisted scientists in documentaries are, I got paid for it.)
A few minutes after I met Edwina. Still in awe.
This documentary was a blast to be involved in and challenged all my skills as a dinosaur expert and biologist as well as a fan of documentaries, monster movies and anatomical artistry! I give a big hat-tip to NatGeo for taking the plunge on this adventure in the first place, to the amazing creature creators, to the film and production crew, to the many jovial journalists I met, and to the four faux-bloodied, surely exhausted dissectors starring in the show– and to Edwina. This was an impressive collaboration drawing together the best that the media, monster-makers and an international team of scientists (aside from the ones I’ve mentioned already, many others too!) can do together. I feel lucky to have been involved, and I think I’ll be looking back on this event as a highlight of my career, especially as a science communicator; much like consulting on Inside Nature’s Giants is a highlight.
I’m as excited as anyone to see how it turns out. Just 2.5 weeks to go — are you excited too? What would you want to see in a T. rex dissection? Where would your first cut be if you did the dissection? “Jurassic World”, what’s that?
This week was a great week for me and giant dinosaurs in many ways, so I’m sharing that experience via photos and a bit of backstory. I hope you like it.
Stomach-Churning Rating: 1/10. Big birds and bones but no barfing.
First, I attended the filming of a new documentary, “T. rex Autopsy” (due for release on 7 June on NatGeo TV, just in time to steal the thunder of get you excited for Jurassic World), on the edge of London. I’m allowed to post these two photos of it. Expect much, much more information later– and I think you will like that information when it comes! Not quite a 50′ tall bird, but… So. Damn. Cool.
Second, my team and I dissected a big animal I’ve mentioned here before. For various reasons, I won’t/can’t post images or details of it right now, but I hope to soon. It’s not a dinosaur, but it was giant as its kind goes, so I’m wedging it in here.
Third, and this is the main impetus for my post, I finally got to see the giant chicken! No, not this one that I recall from my childhood…
But this one! A 50’/13m tall chicken made by teacher Ben Frimet’s team of students and teachers at the City of London Academy!
Shortly after my first encounter. I’m still in a state of awed shock. And shadow.
The megachicken was unveiled at a “Chickenfest” event celebrating the sculpture’s completion. Chickenfest also prominently involved members of the “Chicken Coop” team who have drawn together scientists, humanities scholars, artists and more to investigate “Cultural & Scientific Perceptions of Human-Chicken Interactions” — more details here. Their theme helped unite the event’s various displays and lectures as well as some of the City of London Academy’s teaching topics, which inspired students to look at chickens from many angles. The event was so fun and truly integrative that it had me clucking with joy, but the anatomically accurate giant chicken art piece stole the show (as intended). Enjoy the photo tour below.
Pelvic/thigh region! (no patella, but hey)
Great views from up to 3 storeys around it.
Little chickens made of fast-food forks and stuff. Very clever.
One of our research chickens, a 30-day-old broiler, skeletonized by the Chicken Coop team and brought to the event. Chunky and funky!
Our RVC chicken research team (postdocs/fellows Drs. Heather Paxton, Jeffery Rankin, Diego Pereira Neves) presented a stall with motion capture demos and chicken bones, like this fun identification display.
What will happen to that giant chicken art piece? This is yet to be determined, and was the question asked of the lecture panel (including me, who gave a lame answer involving King’s Cross’s birdcage). It was unanimous that it must not be destroyed– as long as it does not go on a destructive rampage through London…
One of my favourite films of my teenage years, Beastmaster, lends me a phrase I’ll throw out here like a razor-edged boomerang-thing: “Life is a circle. We will meet again.” And so, at the Chickenfest event, past and present worlds collided. I happened to be there presenting a talk just before Luis Rey. Almost exactly 13 years ago, Luis had done this classic T. rex vs. giant chicken race for my “T. rex was not a fast runner” paper in Nature. He likewise has blogged about the Chickenfest event, so check that out!
Coincidentally, there was ANOTHER 50′ tall bird placed not far from that giant chicken in southeast London this week, for a very different reason- a huge Norwegian Blue parrot in celebration of the Monty Python reunion! And I’ve been a Monty Python fan since age ~11, so that rocks my world two times over.
IMAGE: FLICKR USER TAYLOR HERRING
Two giant birds in London in one week. It doesn’t get any better than that– unless there were three such birds– if I missed one, chime in in the Comments!
Stomach-Churning Rating: 5/10: cheesy songs vs. fatty chunks of tissue; there are no better Crimbo treats!
Today is a special day for palaeognath publications, principally pertaining to the plethora of published PeerJ papers (well, three of them anyway) released today, featuring my team’s research! An early Crimbo comes this year in the form of three related studies of hind limb anatomy, development, evolution and biomechanics in those flightless feathered freaks of evolutionary whimsy, the ratites! And since the papers are all published online in PeerJ (gold open access), they are free for anyone with internet access to download and use with due credit. These papers include some stunning images of morphology and histology, evolutionary diagrams, and a special treat to be revealed below. Here I’ll summarize the papers we have written together (with thanks to Leverhulme Trust funding!):
1) Lamas, L., Main, R.P., Hutchinson, J.R. 2014. Ontogenetic scaling patterns and functional anatomy of the pelvic limb musculature in emus (Dromaius novaehollandiae). PeerJ 2:e716 http://dx.doi.org/10.7717/peerj.716
My final year PhD student and “emu whisperer” Luis Lamas has published his first paper with co-supervisor Russ Main and I. Our paper beautifully illustrates the gross anatomy of the leg muscles of emus, and then uses exhaustive measurements (about 6524 of them, all done manually!) of muscle architecture (masses, lengths, etc.) to show how each of the 34 muscles and their tendons grew across a more than tenfold range of body mass (from 6 weeks to 18 months of age). We learned that these muscles get relatively, not just absolutely, larger as emus grow, and their force-generating ability increases almost as strongly, whereas their tendons tend to grow less quickly. As a result, baby emus have only about 22% of their body mass as leg muscles, vs. about 30% in adults. However, baby emus still are extremely athletic, more so than adults and perhaps even “overbuilt” in some ways.
This pattern of rapidly growing, enlarged leg muscles seems to be a general, ancestral pattern for living bird species, reflecting the precocial (more independent, less nest-bound), cursorial (long-legged, running-adapted) natural history and anatomy, considering other studies of ostriches, rheas, chickens and other species close to the root of the avian family tree. But because emus, like other ratites, invest more of their body mass into leg muscles, they can carry out this precocial growth strategy to a greater extreme than flying birds, trading flight prowess away for enhanced running ability. This paper adds another important dataset to the oft-neglected area of “ontogenetic scaling” of the musculoskeletal system, or how the locomotor apparatus adapts to size-/age-related functional/developmental demands as it grows. Luis did a huge amount of work for this paper, leading arduous dissections and analysis of a complex dataset.
Superficial layer of leg muscles in an emu, in right side view. Click any image here to emu-biggen. The ILPO and IC are like human rectus femoris (“quads”); ILFB like our biceps femoris (“hams”); FL, GM and GL much like our fibularis longus and gastrocnemius (calf) muscles, but much much bigger! Or, perhaps FL stands for fa la la la la?
Data for an extra set of emus studied by coauthor Russ Main in the USA, which grew their muscles similarly to our UK group. The exponents (y-axis) show how much more strongly the muscles grew than isometry (maintaining the same relative size), which is the dotted line at 1. The numbers above each data point are the # of individuals measured. Muscle names are partly above; the rest are in the paper. If you want to know them, we might have been separated at birth!
2) Regnault, S., Pitsillides, A.A., Hutchinson, J.R. 2014. Structure, ontogeny and evolution of the patellar tendon in emus (Dromaius novaehollandiae) and other palaeognath birds. PeerJ 2:e711http://dx.doi.org/10.7717/peerj.711
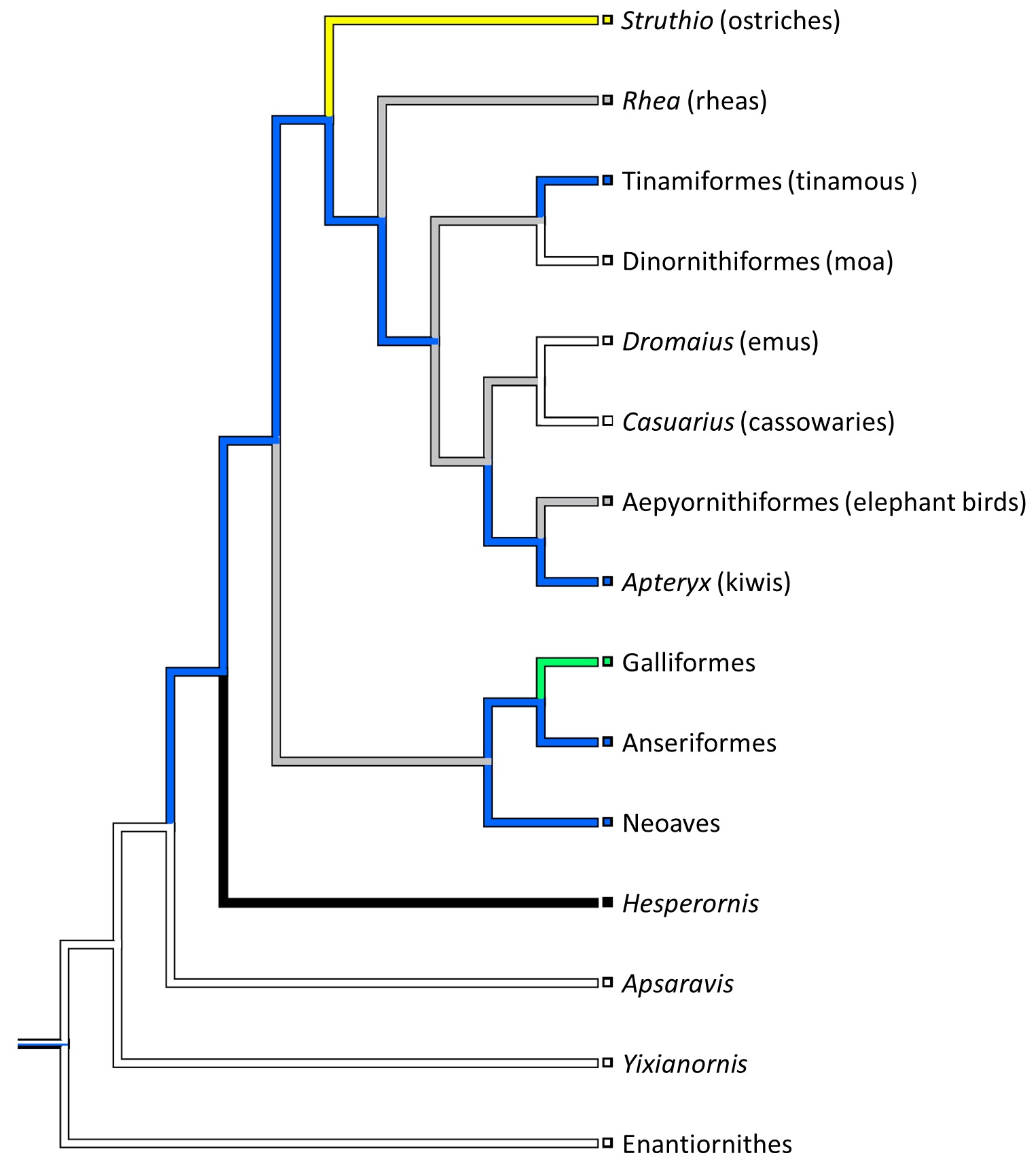
My second year PhD student Sophie Regnault (guest-blogger here before with her rhino feet post) has released her first PhD paper, on the evolution of kneecaps (patellae) in birds, with a focus on the strangeness of the region that should contain the patella in emus. This is a great new collaboration combining her expertise in all aspects of the research with coauthor Prof. Andy Pitsillides‘s on tissue histology and mine on evolution and morphology. This work stems from my own research fellowship on the evolution of the patella in birds, but Sophie has taken it in a bold new direction. First, we realized that emus don’t have a patella– they just keep that region of the knee extensor (~human quadriceps muscle) tendon as a fatty, fibrous tissue throughout growth, showing no signs of forming a bony patella like other birds do. This still blows my mind! Why they do this, we can only speculate meekly about so far. Then, we surveyed other ratites and related birds to see just how unusual the condition in emus was. We discovered, by mapping the form of the patella across an avian family tree, that this fatty tendon seems to be a thing that some ratites (emus, cassowaries and probably the extinct giant moas) do, whereas ostriches go the opposite direction and develop a giant double-boned kneecap in each knee (see below), whereas some other relatives like tinamous and kiwis develop a more “normal”, simple flake-like bit of bone, which is likely the state that the most recent common ancestor of all living birds had.
There’s a lot in this paper for anatomists, biomechanists, palaeontologists, ornithologists, evo-devo folks and more… plenty of food for thought. The paper hearkens back to my 2002 study of the evolution of leg tendons in tetrapods on the lineage that led to birds. In that study I sort of punted on the question of how a patella evolved in birds, because I didn’t quite understand that wonderful little sesamoid bone. And now, 12 years later, we do understand it, at least within the deepest branches of living birds. What happened further up the tree, in later branches, remains a big open subject. It’s clear there were some remarkable changes, such as enormous patellae in diving birds (which the Cretaceous Hesperornis did to an extreme) or losses in other birds (e.g., by some accounts, puffins… I am skeptical)– but curiously, patellae that are not lost in some other birds that you might expect (e.g., the very non-leggy hummingbirds).
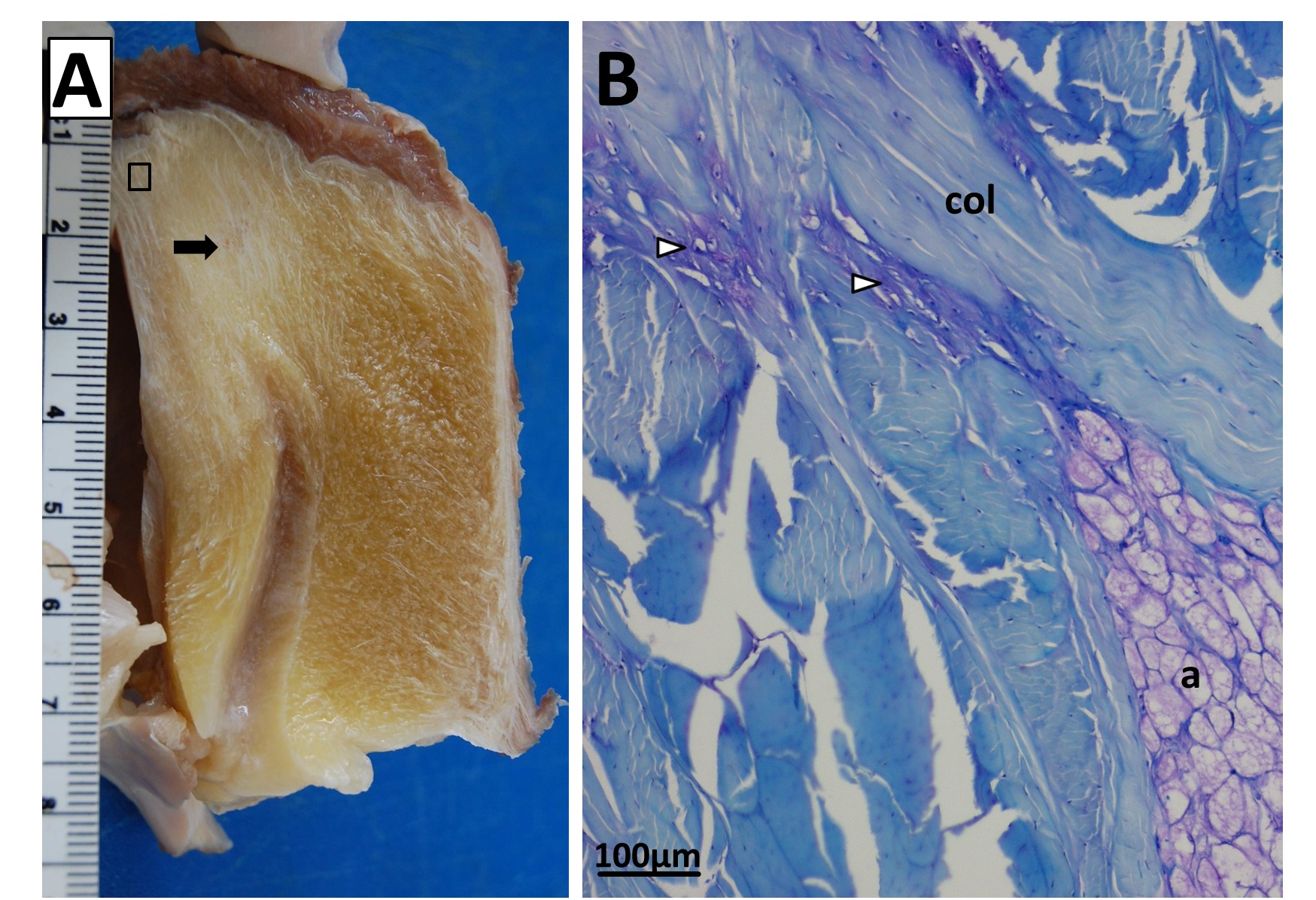
Fatty knee extensor tendon of an emu, showing the absence of a patella. The fatty tissue is split into superficial (Sup) and deep regions, with a pad corresponding to the fat pad in other birds continuous with it and the knee joint meniscus (cushioning pad). The triceps femoris (knee extensor) muscle group inserts right into the fatty tendon, continuing on over it. A is a schematic; B is a dissection.
Sectioning of a Southern Cassowary’s knee extensor tendon, showing: A, Similar section as in the emu image above. revealing similar regions and fibrous tissue (arrow), with no patella, just fat; and B, With collagen fibre bundles (col), fat cells (a), and cartilage-like tissue (open arrows) labelled.
Evolution of patellar form in birds. White branches indicate no patella, blue is a small flake of bone for a patella, green is something bigger, yellow is a double-patella in ostriches, black is a gigantic spar of bone in extinct Hesperornis and relatives, and grey is uncertain. Note the uncertainty and convergent evolution of the patella in ratite birds (Struthio down to Apteryx), which is remarkable but fits well with their likely convergent evolution of flightlessness and running adaptations.
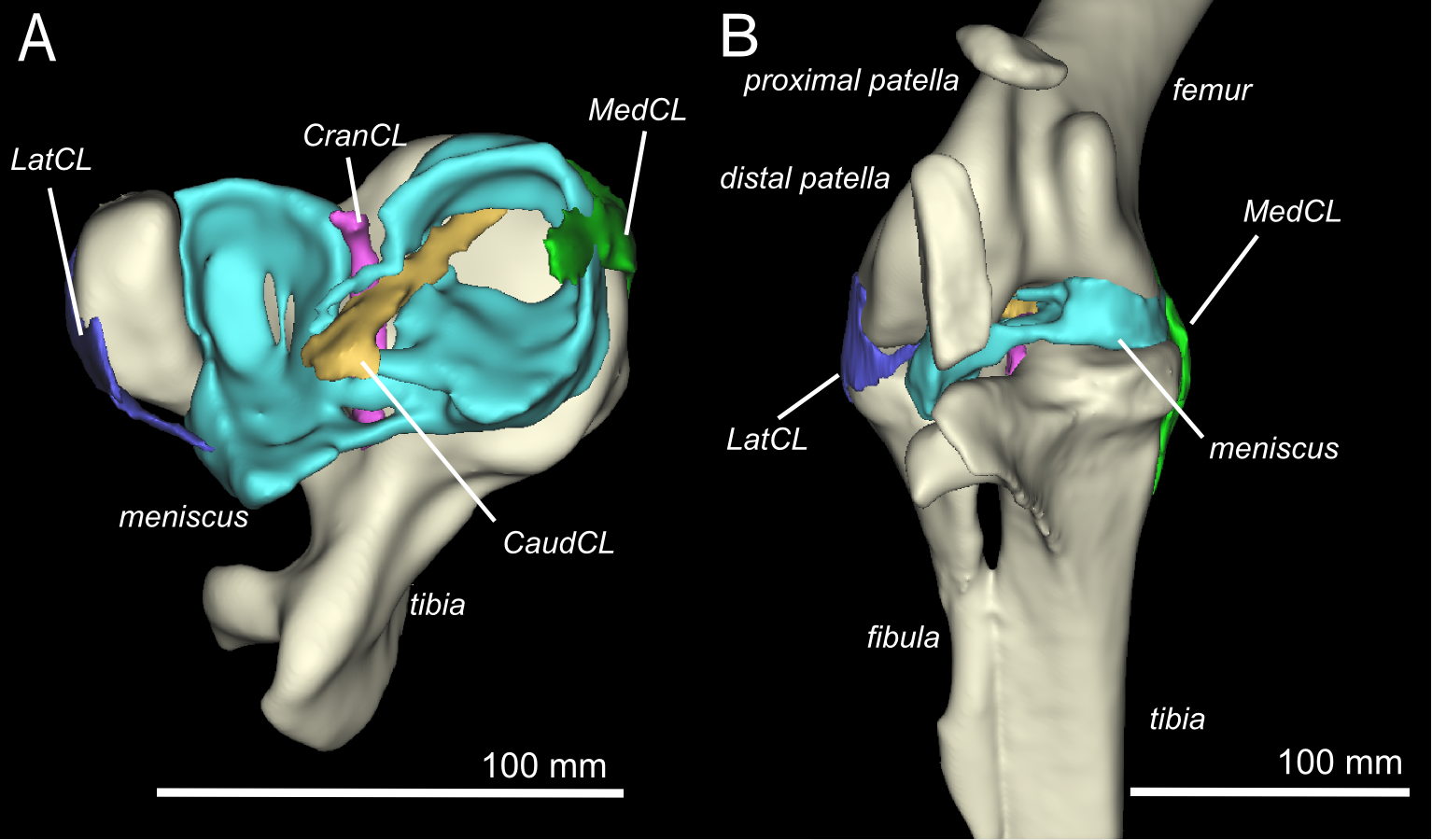
Finally, Kyle Chadwick came from the USA to do a technician post and also part-time Masters degree with me on our sesamoid grant, and proved himself so apt at research that he published a paper just ~3 months into that work! Vivian Allen (now a postdoc on our sesamoid bone grant) joined us in this work, along with Sophie Regnault. We conceived of this paper as fulfilling a need to explain how the major tissues of the knee joint in ostriches, which surround the double-patella noted above, all relate to each other and especially to the patellae. We CT and MRI scanned several ostrich knees and Kyle made a 3D model of a representative subject’s anatomy, which agrees well with the scattered reports of ostrich knee/patellar morphology in the literature but clarifies the complex relationships of all the key organs for the first time.
This ostrich knee model also takes Kyle on an important first step in his Masters research, which is analyzing how this morphology would interact with the potential loads on the patellae. Sesamoid bones like the patella are famously responsive to mechanical loads, so by studying this interaction in ostrich knees, along with other studies of various species with and without patellae, we hope to use to understand why some species evolved patellae (some birds, mammals and lizards; multiple times) and why some never did (most other species, including amphibians, turtles, crocodiles and dinosaurs). And, excitingly for those of you paying attention, this paper includes links to STL format 3D graphics so you can print your own ostrich knees, and a 3D pdf so you can interactively inspect the anatomy yourself!
Ostrich knee in side view: A, X-ray, and (B) labelled schematic.
3D model of an ostrich knee, showing: A, View looking down onto the top of the tibia (shank), with the major collateral ligaments (CL), and B, View looking straight at the front of the knee joint, with major organs of interest near the patella, sans muscles.
You can view all the peer review history of the papers if you want, and that prompts me to comment that, as usual at PeerJ (full disclosure: I’m an associate editor but that brings me £0 conflict of interest), the peer review quality was as rigorous at a typical specialist journal, and faster reviewing+editing+production than any other journal I’ve experienced. Publishing there truly is fun!
Merry Christmas and Happy Holidays — and good Ratite-tidings to all!
And stay tuned- the New Year will bring at least three more papers from us on this subject of ratite locomotion and musculoskeletal anatomy!
♬Should auld palaeognathans be forgot, And never brought for scans? Should publications be soon sought, For auld ratite fans!♪
It’s World Penguin Day! Watch your back though… these penguins aren’t as nice as they seem. But they need us to be nice to them!
Whether you watch a classic GIF like the one above, or a kid-friendly TV/film documentary, you might get the impression that penguins lead carefree, or at least silly or slapstick, lives– happy feet and all that. It works for Hollywood: a Charlie Chaplin comedy relief role to play. And that’s the vision of penguins I grew up with: they were living cartoons to me.
But what’s the reality? Plenty of documentaries, most notably to my mind the recent Attenborough’s “Frozen Earth” episodes or “March of the Penguins” film, have dealt with the darker side to these two-toned, tuxedo-toting antipodeans. And anyone who has experienced penguins in the wild has probably seen those not-so-light facets of penguinity firsthand. On realiizing just how compulsively horny young “hooligan cock” male penguins were, Natural History Museum ornithologist Douglas Russell wrote: ““just the frozen head of the penguin, with self-adhesive white O’s for eye rings, propped upright on wire with a large rock for a body, was sufficient stimulus for males to copulate and deposit sperm on the rock.”
Stomach-Churning Rating: 5/10; some tears may be shed over cute baby penguins and you might choke if you’re a rhea trying to swallow one, but the anatomy shown is mostly skeletal or dessicated. No penguin juices. Except those just mentioned above.
I’m quick to admit, I didn’t know much about penguins until recently. I couldn’t name many species or say much about their behaviour, anatomy or evolutionary history. When I was a graduate student at Berkeley, I was enthused by a now-classic, elegantly simple study (published in 2000) that fellow PhD student Tim Griffin and biomechanist Dr. Rodger Kram conducted on penguin waddling. They found that the waddling gait of penguins isn’t mechanically disadvantageous, as it appears, but rather is a way that they conserve energy while walking. It’s the short legs, instead, that make their gait metabolically expensive, because shorter legs mean that more frequent, costly steps need to be taken, incurring high costs due to rapid firing of leg muscles to support the body. My vicarious enjoyment of Griffin’s & Kram’s research began my scientific introduction to penguins. Fast forward to 2014: I get a crash course in penguinology.
Mostly-fledged Magellanic penguin
That’s what this post is about, and how it brought me in touch with The Existentialist Penguin— the haggard, storm-tossed, predator-harried, starved and bullied wanderer of wastelands.
My personal introduction to penguins over the past year has been initiated by a collaboration with PhD student James Proffitt and long-time colleague Dr. Julia Clarke, both at the University of Texas in Austin. They kindly invited me to collaborate on applying modern biomechanics to the surprisingly excellent fossil record of penguins (Sphenisciformes), among other extant water birds. Before diving into it all, I happened to go to Argentina.
Penguin tries to keep cool in the shade, opening its mouth to shed heat in the autumn sun.
Just before I travelled to Patagonia on unrelated business (to study sauropodomorph dinosaurs!), I did a little googling and came across Punta Tombo reserve, near the city of Trelew that I was visiting (more about that in a future post!). It’s where some 1+ million Magellanic penguins (Spheniscus magellanicus) gather every southern summer to breed and fledge before making a long ~5 month swim up to Brazil. I asked my host, Dr. Alejandro Otero, if we might take a day off to visit this spot, where guanacos, rheas and other wildlife were also said to be common, and he basically said “Hell yes!” as he’d never been there. My Flickr photostream gives a big set of my favourite photos from that trip, but here are some others below, to show some of my experiences. We rented a car and took a lovely 90-minute drive south across the Patagonian plains, observing wildlife like tinamous (yes! So exciting for me) as we went. You could get within 1.5m of the penguins according to park rules, and the penguins were very permissive of that!
This jaunty chap was staying put in his burrow while people walked by. We came closer and he kept rotating his head around, staring at us. I first took it as cute juvenile behaviour, but on later observations of penguins realized it was a threat- “My beak is sharp! Stay back, bro, or I’ll glock ya!”
The video below shows a penguin encounter that left me with no doubts that these animals don’t mess around. The smaller penguin escaped, losing its cool burrow and some of its tough hide, too. Indeed, penguins can be remarkable assholes to each other.
With battles like this erupting all around us, where the penguins struggled to find shade in the desert-like inland parts of the park, often hundreds of meters away from the cool ocean, it came as no surprise to find casualties. The juveniles (and some remaining adults; most having left by now while the ~1 year-old juveniles fledge) not only battled, but also fasted, and roasted in the heat as they shed their insulatory fluff for waterproofed streamlining. This poor little flat Spheniscus had been trodden a bit past streamlined:
Near the end of our visit, just after I saw an informative sign about the lesser rhea or “choique” (Pterocnemia/Rhea pennata), we managed to get very close to a rhea and follow it for a while, as penguins stood around in apparent disinterest. I’ll never forget that meeting: two flightless birds, yet adapted to such different lifestyles and habitats. The penguins were in the rhea’s domain; a hot, wind-blown, scree-scoured scrubland on the edge of the fertile ocean.
The choique soon found a dry old hatchling penguin carcass, no meatier than the surrounding thickets, and tried to swallow it. The loss of teeth by its distant ornithurine ancestors proved to be a bad move, because it struggled to get the jerky-like mass through its beak:
That Punta Tombo visit was an experience I’ll never forget. I returned to the UK, abuzz with excitement about penguins. I “got” them now, I felt, at least in a very unscientific, anthropomorphic way. It took the face-to-beak experience to drive that home, more than any emotive film treatment could. Whether enduring Antarctic wintery blasts or unforgivingly hot and dry, burrow-speckled coastal badlands, penguins are buggers with true grit. Survivors, as their >60 million year fossil record attests to. On my return, I delved through my photos of museum specimens to get a better appreciation for penguin anatomy, preparing to also get familiar with that fossil record; all as part of that ongoing work with Proffitt and Clarke. Here’s some of that anatomy:
My first encounter with a penguin in the wild (but not a live one) is probably this specimen washed up on a beach in Uruguay. I’m going with the tentative ID of a juvenile penguin skeleton (short foot; flat wing bones); probably Magellanic. The bevy of vertebrate morphologists investigating dead penguins on this beach during our conference in 2010 will not soon be forgotten!
Magellanic penguin skeleton, “flying” through the Punta Tombo visitor centre.
University Museum of Zoology Cambridge skeleton of a “great penguin” (do not confuse with the great pumpkin!) species of Aptenodytes; either King (patagonicus) or Emperor (forsteri). Characteristic features, in addition to the robust, dense skeleton, include the short neck, flattened but robust wings and scapulae, robust furcula (wishbone), stubby legs (with a big blocky patella) and thin but longish tail (supposedly used to balance with while walking/standing).
I’ll visit some more penguin anatomy in coming images- those photos are just teasers. And they set the stage for me to go back to my one-stop-shopping for awesome ornithological specimens, the Natural History Museum at Tring (images below presented with kind permission from the Natural History Museum, London; but I took the photos), to pick up an assortment of 11 frozen penguins from helpful curator Hein van Grouw! Such as this “gagged” King penguin:
And this handsome Emperor penguin, going through the Equine Imaging Centre’s CT scanner as I do my usual routine of (1) get cool critters, (2) barrage them with radiation to peek inside:
CT scanner monitors as I scan a penguin; mid-torso x-ray slice shown on the right.
Awwwwww… baby Gentoo penguin (Pygoscelis papua— EDIT: Probably Aptenodytes; see comments below). Unhappy feet, I’m afraid… Happy CT scanning, however– specimens like this are NOT easy to come by in these northern nether regions!
Because I love the CT scan images of these penguins so much (their skeletons are awesome and bizarre!), I’ll share the pilot scans of the best ones now:
Calling all penguin experts! What’s up with this? Is that really how much gastrolith (stomach stone; near bottom of image) volume a penguin carries (answer after some literature reading: maybe yes!), or did a museum curator stick rocks up its bum? It seems very caudal in position, and this is consistent with other animals I’ve seen (some below). A paper on this phenomenon and potential role in ballast is here. Another here.
Side view. Nice view of the head at least.
Young juvenile. Nice pose, and lots of anatomy is shown. And check it out- gastroliths?!? In such a young animal– is it even feeding yet? (presumably straight after hatching) And they are relatively big pebbles, too! If I noticed this 5 years ago, it would have been a nice paper to report- first recognition of gastroliths in penguin chicks seems to have been then. Indeed, that study observed some chicks intentionally swallowing stones.
Another youngun; the fluffy one from the photo above. More rocks up its wazoo.
Three wee little chicks, all with stomach stones.
CT reconstruction of adult skeleton. This specimen was gutted and flattened, so the gastroliths are few and scattered. Check out the long tail:
From recent skeletons to fossil ones, penguins have wacky anatomy; they break most of the “rules” of being a proper bird, putting other oddballs like rheas to shame. I can’t ably review the many penguin species we know of, but the ancient Palaeocene penguin Waimanu features prominently in recent scientific discussions of penguin evolution, such as the superb research and blog of Dan Ksepka as well as many workers in the southern hemisphere. I haven’t had a chance to inspect that creature’s bones, but while in Trelew, Argentina, I was very pleased to run into some excellent specimens of a later animal:
Part of the nice skeleton of Palaeospheniscus patagonicus, an Oligocene/Miocene largish penguin; from the MEF collections in Trelew, Argentina and collected nearby. The genus has been known since Ameghino’s description in 1891, and is closely related to living penguins, especially Aptenodytes. It was not a large penguin, but at about 5kg body mass was no slouch as birds go (roughly similar in size to a Magellanic penguin). I also got to see Madrynornis mirandus, a Miocene form.
For me, the diagnostic trait of a penguin skeleton: the very short, tobust tarsometatarsus. From Palaeospheniscus, as above. The great palaeontologist GG Simpson wrote of it: “Despite the innumerable variations in details, the tarsometatarsi, on which all species but P. robustus are based, are quite stereotyped in general structure and leave little doubt that the forms placed here by Ameghino do all belong to a natural group.” A ratio of length to proximal width of >2 is typical of most penguins. Synapomorphy FTW!
From beach skeletons, to mass suffering of landbound birds, to 3D imaging and fossil skeletons, I’ve had quite the immersion in penguinness lately. And through that experience, I’ve been drawn closer to penguins in more ways than one. I’ve been impressed by their adaptability and durability. In some ways, penguins’ adaptations to harsh freezing winters in wastelands also aid them to survive harsh baking summers in dry badlands.
Yes, those badlands are still coastal, and penguins can still drink the saltwater and excrete salt via their supraorbital glands, but those penguins in Punta Tombo were not having a keg party. They were clearly enduring some serious discomfort, and not all making it through the ordeal. I watched silently along with other penguins as one penguin lay prone in an awkward pose on a bleached-white stretch of hardpan soil, while one flipper meekly raised, then flopped down. It was not long for this world, and there was a host of large scavengers around ready to make the most of that, while penguin-eating giant petrels (a sister group to penguins) wheeled overhead.
Waddlers of the wastes
While penguins still spend most of their lives at sea, they retain a sometimes astonishing array of behaviours they use on land: burrowing, hopping/jumping, costly short-legged (but efficiently waddling) walking, and perhaps more that we haven’t yet discovered! Their unique anatomy reflects a compromise between all these factors, and we’re fortunate to have knowledge of their fossil record that shows a lot of detail on how they evolved it all. While penguins are a highly aquatic species, they show how aquatic and terrestrial adaptations can coexist in harmony; it’s not just a black-or-white issue. But with climate change in progress, the ~18 species of penguins have some rapidly altering challenges to adapt to, or go the way of Waimanu. This is a critical Kierkegaardian moment for The Existentialist Penguin.
I raise a glass in toast to that versatile, resilient, gravel-gizzarded Existentialist Penguin! May it persevere all the troubles our ever-changing world throws at it, as it has done since the Palaeocene. And may we draw inspiration from its tenacity, to face our own troubles, together on this crazy spinning globe!