I’m letting the dogs out today. Science gone barking mad! Hopefully my puns will not screw the pooch.
Stomach-Churning Rating: 4/10; a dog cadaver’s leg (not messy), then just tame digital images of anatomy.
I am working with Rich Ellis, a former MSc student at Univ. Colorado (see his cool new paper here!), for a fun new collaboration this year. He was awarded a prestigious Whitaker Foundation scholarship to do this research, which focuses on how different animals stand up from a squatting position, with the legs about as bent as they can be.
We want to know how animals do this standing up movement, because it is in some ways a very demanding activity. Very flexed/bent limb joints mean that the muscles (and some tendons) are stretched about as far as they ever will be. So this places them at disadvantageous lengths (and leverage, or mechanical advantage) for producing force. We know almost nothing about how any animal, even humans, does this-– how close to their limits of length are their muscles? Which muscles are closest? Does this change in animals with different numbers of legs, postures, anatomy, size, etc? Such fundamental questions are totally unaddressed. It’s an exciting area to blaze a new trail in, as Rich is doing. So far, we’ve worked with quail, humans, and now greyhounds; in the past I did some simple studies with horses and elephants, too. Jeff Rankin from my team and other collaborators have also worked on six species of birds, of varying sizes, to see how their squat-stand mechanics change. Thus we’ve covered a wide diversity of animals, and now we’re learning from that diversity. “Diversity enables discovery,” one of my former PhD mentors Prof. Bob Full always says. Too true.
Greyhounds are interesting because they are medium-sized, long-legged, quadrupedal, quite erect in posture, and very specialized for fast running. Fast runners tend to have big muscles with fairly short fibres. Short fibres are bad for moving the joints through very large ranges of motion. So how does a greyhound stand up? Obviously they can do it, but they might have some interesting strategies for doing so- the demands for large joint motion may require a compromise with the demands for fast running. Or maybe the two demands actually can both be optimized without conflict. We don’t know. But we’re going to find out, and then we’ll see how greyhounds compare with other animals.
To find out, we first have to measure some dogs standing up. We’ve done that for about 8 greyhounds. Here is an example of a cooperative pooch:
Those harmless experiments, if you follow me on Twitter, were live-tweeted under the hashtag #StandSpotStand… I dropped the ball there and didn’t continue the tweeting long after data collection, but we got the point across– it’s fun science addressing useful questions. Anyway, the experiments went well, thanks to cooperative pooches like the one above, and Rich has analyzed most of the data.
Now the next step involves the cadaver of a dog. We could anaesthetize our subjects and do this next procedure to obtain subject-specific anatomy. But it really wouldn’t be ethically justified (and if I were an owner I wouldn’t allow it either!) and so we don’t. A greyhound is a greyhound as far as we’re concerned; they’ll be more like each other than either is like a quail or a human. Individual variation is a whole other subject, and there are published data on this that we can compare with.
We get a dead dog’s leg — we don’t kill them; we get cadavers and re-use them:
We study the hindlimb because birds and humans don’t use their forelimbs much to stand up normally, so this makes comparisons simpler. We’re collecting forelimb data, though, as we work with quadrupeds, for a rainy day.
We then CT scan the leg, getting a stack of slices like this– see what you can identify here:
It’s not so clear in these images, but I was impressed to see that the muscles showed up very clearly with this leg. That was doggone cool! Perhaps some combination of formalin preservation, fresh condition, and freezing made the CT images clearer than I am used to. Anyway, this turned out to be a treat for our research, as follows.
We then use commercial software (we like Mimics; others use Amira or other packages) to “segment” (make digital representations in 3D) the CT scan data into 3D anatomy, partitioning the greyscale CT images into coloured individual objects– two views of one part of the thigh are shown below.
What can you identify as different colours here? There are lots of clues in the images (click to embiggen):
And here is what the whole thigh looks like when you switch to the 3D imaging view:
Quite fetching image, eh?!
The next steps after we finish the limb segmentation are to apply the experimental data we observed for greyhounds of comparable size by importing the model and those data into biomechanics software (SIMM/OpenSim). We’ve done about 40 models like this for various species. I detailed this procedure for an elephant here.
Then, at long last, science will know how a greyhound stands up! Wahoo! Waise the woof! Stay tuned as we hound you with more progress on this research-as-it-happens. Rich just finished the above thigh model this week, and the rest of the leg will be done soon.
Many thanks to Rich Ellis for providing images used here. And thank you for persevering my puns; they will now be cur tailed.
Happy Freezermas! Sing it: “On the fifth day of Freezermas, this blo-og gave to me: one tibiotarsus, two silly Darwins, three muscle layers, four gory hearts, a-and five stages modelling a doggie!” ♪♫
Today I’m shimmying down your interwebz with a late delivery. I’ve promised before to show how we clean up our nasty gooey skeletons to preserve them for future research to use. This is the intended final destination of all critters that are tenants of my freezers– the freezer is just a lovely holiday home, but bony heaven is the end result. I’ve accumulated a little museum of the bones of exotic animals I’ve studied, using these cleaned specimens. Here is how I do that preservation– there are four basic steps, and I’ll show them in four photos.
Stomach-Churning Rating: 8/10; first just dry bones, but then some gooey bones and by the end we ratchet it up to bloody organs.
Step 1) We get the deceased animal from various zoos and other EU sources, CT/MRI scan it, and dissect it. That’s what most of this blog focuses on, so I won’t show that. But I will show the end result, and then how I get to that:
Those are some elephant and rhino bones, some of which you saw on the 2nd day of Freezermas. Elephant bones are super greasy; it’s almost impossible to get rid of that brown grease visible in this photo (upper LH side) without making the bones brittle and over-bleached. The bones of the whiter white rhino on the right show what I’m usually aiming for. How do I get this done? Well, here’s an example for an elephant shank:
I take the elephant shank and make soup. (above) An Asian elephant’s patella, tibia and fibula were dissected, frozen for many years (queued up for cleaning; much freezer burn occurred on this specimen— it was jerky-fied), and then thawed. I put large specimens in this Rose cooker unit, which is a big ham cooker with a heater unit at the bottom. My baby, a Rapidaire MKV 5-ham unit is shown; oooh, ahhh!
The Rose cooker is filled up with tap water and been set it at around 60-90C. Then I let it cook away! A brothy soup develops, and sometimes it smells rather nice (my favourite aroma is giraffe leg). Sometimes… it’s not so nice. We check it every few hours to top up the water and remove stray tissue, and then change the water every day or so.
An elephant shank like this will take 2-3 days of cooking, longer if only switched on during work hours. The key thing is not to let it cook dry, which happened once with a faulty Rose cooker that did not do its normal auto-shutoff when the water ran low… showing up to work to encounter some fire trucks and unhappy college Health & Safety rep is not a good way to start your day, trust me!
This step is only slightly different for smaller (<30cm) specimens. Rather than the Rose cooker, we use the lovingly named “Croc Crock”, which isn’t visually impressive but you can see it here. As the name indicates, we’ve mainly used it for small crocodiles, and it is a crock pot. (a helpful thing is to add some detergent to the water for these small specimens; then bleaching isn’t so necessary)
Step 2) Then I empty out the water through the bottom spout, do the very nasty job of cleaning out the fat and other tissue that has accumulated (think 20 gallons of goo), hose off the bone, and set it in a ~10% bleach solution for at least a day, or up to a week or so for an elephant bone. Once it’s cleared up, I leave it out to dry (for big elephant bones, copious amounts of grease may be emerging for a few weeks). And then…
Step 3) I varnish the dry bones with a clear varnish, and let them dry. Here is how that elephant shank turned out. Pretty good! Finally, they get to join their friends:
Step 4) The prepared bones are labelled, given a number/name that I file in a world class comprehensive electronic database (cough, get on that John, cough!), and they become part of my humble mini-museum, shown above. Voila! The chef’s job is finished. Let science be served!
Happy Freezermas! Sing it: “On the fourth day of Freezermas, this blo-og gave to me: one tibiotarsus, two Darwin pictures, three muscle layers, a-a-and four steps of bone cookery!” ♪♫
Oh it’s Valentine’s day, so, err, have a heart today. Have four, actually!
For the previous days of Freezermas we first had 1 picture, then 2, now guess how many we have today? Right, we’ve settled into a groove and have three (plus one silly one). Today is fresh beefy anatomy day! No focus on bones, but on soft tissues– however, once again, I’m representin’ bird legs! And this time, no mystery things to identify; sorry. But if you want to muscle in on some myology, today is the day for you. I will unwrap the thigh of an ostrich and consider the major muscles that power rapid running in this biped, and how they illuminate the evolution of bipedal motion along the line of descent to birds. For more ostrich escapades, see this old post. And we’re off!
Stomach-Churning Rating: 7/10; plenty of fresh, red, meaty meat from ostrich leg muscles.
Here you are looking at a right hindlimb of an ostrich, in side/lateral view. To help orient yourself, the hip lies deep in the middle of the image and the knee is the rounded bump near the bottom right corner, with the shank angling sharply back toward the bottom left.
I’ve labelled six muscles in yellow. As usual for sauropsid (bird/reptile) pelvic limb muscles, they have sensible names that reflect their attachments. They don’t have so many silly old mammalian names like pectineus or latissimus, which tell you rather little about the muscles themselves. We can thank 19th century anatomists like two of my anatomist heroes, Hans Gadow and Alfred Romer (who refined Gadow’s earlier work and made it more popular among English-speakers and palaeontologists), for that enlightened nomenclature.
The six muscles seen above are the IC (iliotibialis cranialis), IL (iliotibialis lateralis), “AMB2?” (one of the ambiens muscles– correctly identified; ignore the ?), ITC (iliotrochantericus caudalis), CFP (caudofemoralis pars pelvica) and FCLP (a mouthful to say: flexor cruris lateralis pars pelvica). The ambiens is the one oddly, non-anatomically named muscle, and has nothing to do with helping you sleep (pssst– wake up! Muscles are exciting!), but everything to do with the state of total awesomeness, which is what “ambiens” means. Maybe. Or I am making shit up.
The IC, IL and AMB2 are parts of the triceps femoris group (discussed in my 1st Freeezermas post), or for mammal fans the quadriceps. The IC and AMB are in front of the hip so they flex it (move the thigh forward; protract it); the IL is right around the hip so it can flex or extend the hip (protract/retract the femur); all three of these can extend (straighten) the knee joint to varying degrees. The IC is fairly typical for a bird except for its size, and helps to quickly swing the leg through the air between steps. Some birds have multiple parts of the IL, but ostriches and many others have simplified it to one major mass; regardless, it is a major muscle used to support the weight of the body.
The AMB2 is a remarkable muscle unique to ostriches; it can also be called the dorsal ambiens muscle. Typical birds just have a single head of the AMB sitting on the preacetabular (pubic) tubercle, so in front of and below the hip. It has a crazy tendon that snakes past the knee (in some birds, perforating/grooving the patella) into the lower leg muscles and may be able to even pull on the toes. But ostriches, for some reason, added a second head of this muscle that was shifted way up onto the front of the pelvis (the ilium; dorsal bladelike bone). Crocodilians also have a 2nd ambiens muscle but in a different position, and almost certainly as an example of convergent evolution. The function of the ambiens is mysterious, but this muscle has featured prominently in avian systematics/taxonomy, evolution (invoked as a key muscle used in perching) and more.
These muscles of the triceps femoris group are easily identifiable in crocodiles and other reptiles because they are remarkably similar in their attachments. The main changes these muscles experienced during the evolution of bipedalism, dinosaurs and later birds are simply proportional– they got bigger, with stronger, larger attachments on the pelvis and the front of the knee (the CC/LC, if you remember from Freezermas day 1).
The ITC is a muscle that is very dear to me. I’ve written a lot about it, and I love saying the name “Iliotrochantericus caudalis”- it is musical to me. For mammal fans, think gluteal muscles (medial gluteal in particular). It is a huge, pennate muscle (short and strongly angled muscle fibres in a “sandwich” with a tendinous sheet between the two layers of fibres). It has a short, broad tendon that wraps around the trochanteric crest (a structure on the upper front end of the femur with a history that goes wayyyy back into dinosaurs; long story!) to insert in a scarred depression. The ITC seems to mainly rotate the femur around its long axis to help support the body. I could go on and on about this muscle, which is part of the enigmatic “deep dorsal” thigh muscle group — the homologies of this group among land vertebrates are still controversial and confusing. But I will spare you the on-and-on. Incidentally, the ITC is the “oyster” in birds that is the best cut of meat. And in ostriches it makes a massive steak.
The CFP also has a cool evolutionary history. It runs from the back of the pelvis to the middle of the femur, closely adjoined to the caudal head of the muscle (CFC), which is more vestigial. In birds the CFP is usually not a large muscle, but in other sauropsids/reptiles it can be fairly hefty, although almost never as hefty as its more famous counterpart the caudofemoralis longus (= CFC in birds). Probably any dinosaur specialist is familiar with its origin and its insertion: respectively, the “brevis fossa” on the back of the ilium; a big shelf of bone; and the fourth trochanter of the femur; a crest of bone that is reduced to a scar/tubercle in birds. Much like its tail-based counterpart, the CFP became progressively reduced closer and closer to birds. This is related to a reduction in the amount of movement of the femur/thigh during locomotion, as birds shortened their tails and shifted their balance forward, as Steve Gatesy showed in a classic 1990 paper. Hopefully there will be more about this subject in a future paper from my team…
The FCLP is another muscle that didn’t change much, except by getting larger as we trace its evolution from early reptiles to birds. It is a “hamstring” muscle that is an important power source during locomotion in birds like the ostrich, because it retracts the lower limb (flexes the knee; hence flexor cruris in its name) as well as the femur/thigh (extends the hip). Your semitendinosus muscle is a good comparison to it. Indeed, these two differently named muscles are homologous– our very distant tetrapod ancestor had the same single muscle, and its descendants didn’t change it that much on our lineage or on the avian/reptile one.
I’ve reflected the IL muscle out of the way so we can see the second layer of muscles underneath it. Now we see two more muscles of the thigh, and large ones at that– the FMTL (femorotibialis lateralis) and ILFB (iliofibularis).
The FMTL simply is a part of the triceps femoris group that only comes from the femur and hence only, but due to its large size powerfully, straightens the knee. Unlike the other muscles in this group, it has no action about the hip joint. It is very similar to your vastus lateralis muscle: its fleshy origin dominates the surface of the femur (thigh bone). There are two other parts of that muscle, hidden in this figure, much like our vastus group has multiple parts. Again, this is a muscle that enlarged on the lineage leading to modern birds.
And that evolutionary enlargement applies, too, to the ILFB, whose prominent insertion I discussed on day 1 of Freezermas. This huge “biceps” muscle (it is single-headed unlike in humans, so the name “biceps” does not apply well) is the most powerful of the “hamstring”-type muscles that extend the hip and flex the knee. Therefore it is important for the “knee-driven” locomotion of birds. And hence the ILFB enlarged during avian evolution– which is very evident from changes of both its bony origin on the back of the pelvis/ilium and its insertion on the fibula.
Here, for the terminus of today’s trio of struthious tributes and tribulations, I’ve moved the ILFB out of the way so you can see the various inner/medial layer of thigh muscles. Some of the former muscles are more exposed now, and we can see three new ones: the FCM (flexor cruris medialis), PIFM+(PIF)L (the tongue-twisting puboischiofemoralis medialis et lateralis), and tiny ISF (ischiofemoralis).
The FCM (~mammalian semitendinosus) is merely another, smaller part of the FCLP’s “hamstring” group, and its thin tendon blends with that of the FCLP, so it very much works with that muscle in locomotion, and has a similar evolutionary history.
The PIFM+L are “adductors”, but in birds they don’t really do any adduction (drawing the legs inwards) because they are right behind, rather than below or inside, the hip. They act as hip extensors/retractors of the femur, and probably aid more in holding the femur steady (“postural muscles”) than playing a major role in producing power for locomotion like the ILFB/hamstring group does. In earlier reptiles, they were much more important, for preventing the legs from splaying too far away from the body.
The ISF is usually quite a large muscle in birds, but ostriches and some other ratites have reduced it to a thin slip of muscle– often mistaken for other muscles (indeed, like a few other muscles I’ve described here, modern anatomists still get confused by this muscle– an otherwise superb recent description by Gangl et al., among others, mis-identifies this and some other muscles— an error an upcoming paper from my group will rectify). Normally the ISF sits atop a bone-free window on the outer surface of the pelvis, the ilio-ischiadic fenestra (literally a window in Latin) in birds; in ostriches it has moved more onto the ischium. In contrast, in other sauropsids it lies inside the pelvis, so during its evolution it became more lateral, but the insertion on the upper femur was maintained. It is a weak rotator and extensor of the hip, especially in ostriches in which its role is probably proportionately puny.
And there you have read a healthy chunk of my 2001 PhD thesis, condensed into less jargonious language. You might now know almost half of the key muscles of the avian hind limb. If you made it this far, you are one awesome anatomical enthusiast. If you eat meat, apply this lesson to the next chicken thigh you consume, to consumate this enthusiasm.
A broader point I’d like to make here is that anatomy is best conveyed not only along with the functional narrative (How does anatomy work?) but also the evolutionary tale (Where did anatomy come from and What were the consequences of its changes? Why did it change?). This takes it away from dry memorization of terms and locations, and carries it into the realm of explaining why nature is the way it is, and how every organism’s biology has a richly detailed and complex background. This style portrays nature as much more like that tangled bank that Darwin so enchantingly envisioned. I’ve tried to do that justice here, using this one ostrich whom we affectionately called Twinkletoes, or Twinkie, when we dissected it back in 2002.
Happy Freezermas! Sing it: “On the third day of Freezermas, this blo-og gave to me: one tibiotarsus, two silly pictures, a-and three muscle layers from Twinkie!”
…a daily picture of anatomy! And today it is two pictures, tra-la-lee!
Welcome back to Freezermas! And HAPPY DARWIN DAY! Last year our whole lab got involved in DD2012, but this blog was just a twinkling in my keyboard back then. This year it was a more mellow, somber occasion for DD2013. But Heinrich Mallison of the dinosaurpalaeo blog took part, and took photos (all credits go to him), and the result kicked ass and took names. Bring it on!
Here is Darwin amidst a selection of greatest hits from my bone collection; post-freezer denizens. How many can you identify? Have a go in the comments below. A few should be quite familiar to blog followers… More about these bones later this week. Incidentally, Darwin is standing on a Kistler forceplate. So biomechanics afficionados can geek out about this, too.
And here I am hamming it up again. Give it a rest, John! But ’tis merely a humble offering to The Master. I’m sure he’d appreciate it. Any guesses what it is?
Happy Freezermas! Sing it: “On the second day of Freezermas, this blo-og gave to me: one tibiotarsus, a-and two silly pictures with Chucky D!”
Welcome to Freezermas! In the dead of winter, the WIJF blog jumps down your internet to deliver mind-warming science, and images, and evolution! To celebrate Charles Darwin’s birthday (204th = tomorrow Feb 12, 2013), I’m bringing you one Anatomy Vignette each day this week (we’ll see if I can manage the weekend or not)! Let’s do this!
Stomach-Churning Rating: 2/10; just bones; one picture of them, and then a lot of discussion of muscle anatomy but no pictures of it.
The above image comes from one of my old, somewhat obscure anatomy papers (link to pdf here), from 2003. It’s possibly the first figure I made, entirely by myself, that I’m sort of proud of. It doesn’t totally suck compared with some of my other attempts. I did the stippled line drawing on the left, and on the right is one of my first usages of a digital photo in a paper (digital cameras were finally up to the task around that time; I used my new Nikon Coolpix 900, if memory serves). It was a greatly improved figure over what I’d submitted for this paper originally, which was a rushed, half-baked manuscript for a SICB conference symposium on tendons. I’ll never forget one of the peer reviews of the manuscript, which said something like “the text of this paper is a joy to behold, but the figures are a horror.” They were right, and luckily the images in the paper I submitted changed a lot in revision. (I’m still embarrassed by the incident, though!)
Anyway, the picture is of the lower hind limb of two theropod dinosaurs: (a,c) an adult Tyrannosaurus rex, and (b,d) a wild turkey (Meleagris) from my personal collections of dissected-then-skeletonized animals (this turkey became a biomechanical model in a 2004 paper of mine, too!). In both cases we’re looking at a right hind limb; in (a) and (b) from a caudal/posterior/rear view, and in (c) and (d) from a lateral/side-on/profile view.
If you’re having trouble visualizing these bones in the real animal, check this T. rex skeleton in rear and side views and try to find these bones. You can do it! You might also want to look back at my paroxysmic outburst of love for knee joint anatomy.
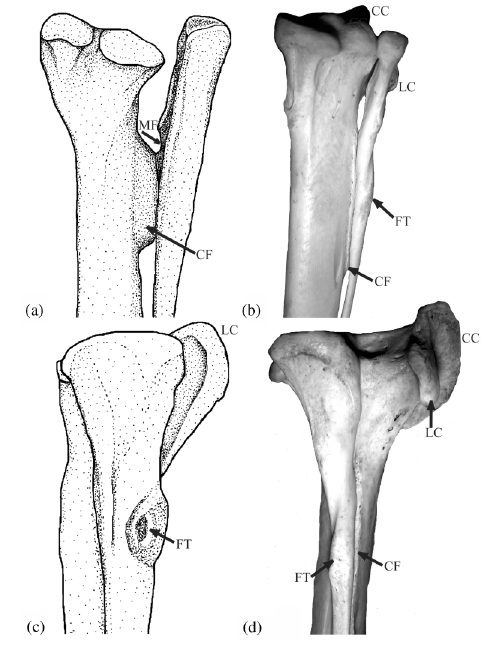
The thicker long bone is the tibia (your main shank bone; or in a lamb shank, chicken drumstick, etc); the thinner outer bone is the fibula. Together with some smaller bones, for brevity we can call them the tibiotarsus — but only in theropod dinosaurs, or you will anger the freezer gods.
The labels show some cool anatomical features, as follows:
“CC” the cranial cnemial crest of the tibia (a projection of bone unique to the knees of birds);
“CF” the crista fibularis; or fibular crest; of the tibia (more about this below);
“FT” the fibular tubercle (insertion of the big hamstring/biceps muscle M. iliofibularis);
“LC” the lateral cnemial crest of the tibia (a big arching swath of bone that both birds and non-avian theropods like Tyrannosaurus have; the CC is just pasted on top of this in birds); and
“MF” which denotes a muscle fossa (depression) on the inner surface of the upper end of the fibula, which presumably housed a muscle (M. popliteus) binding the fibula to the tibia in earlier dinosaurs, but is vestigial in birds.
The CF, or fibular crest, is a feature that only theropod dinosaurs, among reptiles, develop like this. It evolved early in their history and thus was passed on to birds with other ancient features like hollow bones and bipedalism. It binds the fibula closely appressed to the tibia, making those bones act more like a single functional unit –and sometimes they even fuse together. The CF also transmits forces from the whopping big M. iliofibularis muscle’s insertion (the FT label) across the puny fibula onto the robust tibia. The MF once held a muscle that also helped keep those two bones together, but additionally it could have contracted to move them relative to each other a little bit, as in other living animals (many mammals and reptiles have a big M. popliteus and/or M. interosse[o]us). So these features all have a common functional, anatomical and evolutionary (and developmental; different story for evo-devo fans) relationship. By binding the fibula and tibia together, these structures helped early bipeds (the first theropods and kin) support themselves on one leg at a time during standing and moving, and also helped begin to reduce the limbs to lighten them for easier, faster swinging. So we can think of these features as specializations that helped theropod dinosaurs, and ultimately birds, get established as bipedal animals.
The CC and LC have a similar story to tell; for one, they are muscle attachments, again mainly for thigh muscles. And again, the LC dates back to early theropods (and some other dinosaurs had a version of it; usually smaller). These crests serve mainly as insertions for the “quadriceps” (in human/mammal terms) or triceps (in reptile/bird terms) muscle group’s major tendon, spanning from the pelvis/femur across the thigh and knee to this region. In birds, we call this structure of insertion the patellar tendon or (less appropriately) ligament. But dinosaurs had no patella, ever, so the triceps femoris tendon would be the proper technical term. Regardless, that crest (LC, and later LC too) helped the attached muscles to straighten the knee joint or support body weight during standing/moving, by giving them better leverage. So it would have been important for early bipeds, too, like the CF, MF and other features above. Your cnemial crest (tibial tuberosity) is pathetic by comparison. Don’t even look at it. Droop your knees in primate shame!
Bumps and squiggles on bones might seem puny details just for anatomists to study and describe in long, tedious monographs, but each is part of the great story of evolution, and each has a story to tell that fits into that story. Back in Darwin’s day, some of the world’s greatest scientists of the age (Richard Owen and Thomas Huxley being but two spectacular examples) pored over these seemingly innocuous features, and so they became part of nascent evolutionary theory even then. This week, I’ll be celebrating a lot of those details, which I still feel are important today, and the stories they help to tell.
Happy Freezermas! Sing it: “On the first day of Freezermas, this blo-og gave to me: a tibiotarsus with a CF and FT!”
I’m not sure if this is a new tradition at this blog or not (probably not), but hey let’s give it a name: an Anatomy Vignette. Just something curious I notice during my research that deserves more than just a tweet. I borrowed some bones from the University of Cambridge Museum of Zoology (whom I love, because they have great exhibits and are very research-friendly) to CT scan for some projects. I noticed this:
And I thought “Ouch! That’s nasty, dude.” (the holes in the bone just above the knee joint– these should just be a roughened area where the adductor muscles and other leg muscles attach)
So I was interested to see the CT scan images to find out how these possibly osteomyelitic lesions continued into the bone. They’re really pervasive, continuing into the marrow cavity quite far up the femur, as this shows (good CT-viewing practice to match up what you are seeing in the photo above with this movie):
I would be surprised if this was not the reason this animal died (presumably being euthanased at a UK zoo). There would have been extensive infection and pain resulting from this bony disease. How did it originate? Who knows. Maybe the animal strained a muscle and bacteria got inside, or maybe there was a fall or other injury. Hard to tell.
Oh, and also note the lack of a true marrow cavity in hippos, which is true for all the long bones. The “cavity” is filled in with cancellous bone. Same with rhinos, elephants, and many other species… science doesn’t entirely know why but this feature surely does help support the body on land, and grants at least some extra negative buoyancy in water; at a cost of some extra weight to lug around, of course.
One for the weekend morning crowd here. The early bird gets the… cadaver?
At last I’ve managed to pore thru my photos and find something that works for a Mystery Dissection image, so without further adieu here it is! Answer will come tomorrow (Monday) night.
What is the largest structure evident (i.e. what is the picture mainly featuring) and from what group of organisms (be as specific as you can).
Remember, we have a scoreboard now, and rules for scoring. See here. Regular points for this round– Xmas is over, folks!
To recap, Mark Robinson is in the lead w/14pts, tied w/Filippo, but with Heinrich and RH close behind at 12 pts, followed by the 5-person Gang of Awesomeness at 7 pts.
Today I’m doing something a bit unusual for this blog, but which very comfortably fits within its theme. Enough talking about my papers and media appearances and such. Too much self-indulgence, I feel. I want to talk about someone else. And then I will get back to the usual business of this blog: sharing the joy of cool anatomy, with a Mystery Dissection/Image post that is long overdue. Yet first, I wish to share the joy of knowing a cool anatomist– and artist.
One of my great shames as a scientist is that I never cultivated some decent artistic skills that I had as a young boy. And now as an anatomist I feel that my work often suffers from a lack of artistic talent (e.g. the image below, which still makes me hang my head in shame). In addition to scientific know-how, anatomy, when done best, demands the eyes and the hands of an artist. I might have the eyes but I definitely lack the hands. I envy people that have both; Julia Molnar from my team is but one example. And for me, encountering them is always a special delight. What follows is my personal perspective on one of the shining stars in the field.
Right ischium (hip bone; pelvis/synsacrum) of an adult ostrich in side view, showing some muscle origins and stuff, with a 1cm scale bar. Cringe. My boring, amateurish, pixelly line drawing from a paper on pelvic evolution. I hate it and wish I’d done better.
Mieke Roth is a scientific illustrator from the Netherlands, with a Masters degree from the prestigious and hallowed halls of Wageningen University. Her homepage has the tagline “Complex processes beautifully revealed”. This is a wonderfully succinct and eloquent way to describe the magic that she is able to conjure with her skills in scientific illustration. Her “Ultimate Croc Anatomy” project will be the greatest weaving of that sorcery yet. That project is described and will be documented on this page, and has an Indiegogo crowdfunding page here. Mieke describes its goals best:
“I will meticulously dissect a Nile crocodile and document it. I will share the dissection in text, illustrations and video via my website. I will process the data I gathered and each time build a new part of the digital crocodile. From there I will adapt the model for illustrations, books, animations and apps.”
I first became a happy victim of the artistic spells that Mieke casts when I saw her blog entry “How to make an octopus,” which you absolutely must read if you have not already or else you’ll be firmly spanked and sent away from this blog with only lumps of coal in your freezer for Freezermas (what’s Freezermas? Find out soon, and in the meantime be nice– or else!). In her post, Mieke didn’t just look up a picture of an octopus somewhere and redraw it in an abstract, schematic, flat and deceptively simplified way. She went and did her own hands-on research by dissecting an octopus, and then described the steps in converting those observations into a brilliantly novel set of digital illustrations that really brought octopus anatomy to life.
Octopus image by Mieke Roth
Part of what first mesmerized me is that this whole investigative and creative process was lovingly documented on her blog. In doing this, Mieke played the roles of both scientist and artist, by displaying the mundane-but-wonderful labour she did to come up with her final, gorgeous results, and the passion, dedication, scholarship and originality that make her stand head and shoulders above so many gifted scientific or medical illustrators. This thrilled me at both visceral and intellectual levels. It literally gave me chills to witness how good the final product was. As I write this and look back on that blog again, months later, I still feel the magic.
So then I started browsing around her homepage and became punch-drunk from repeated blows of amazement—again and again and again the quality and novelty and thoroughness leaped off the computer screen. Images of nature that I thought I knew well, such as the growth of a chick into a chicken(really great blog here documenting this with sketches), were conveyed in a way that made me see them as if for the first time, with joy and wonder. In the space of a day, I became a huge Mieke Roth fan.
Young chicken sketched by Mieke Roth
And now Mieke is taking it not just a notch, but a huge step, to spend a year documenting the anatomy of the Nile crocodile—how cool is that!?! As an expert in the postcranial anatomy of the Crocodylia, I can confidently state that the available scientific literature on the subject is patchy in coverage and often poor in quality by modern standards. There are big flashes of excellence here and there, such as Larry Witmer’s and Chris Brochu’s teams’ very thorough work on head and neck anatomy, or Colleen Farmer’s and Emma Schachner’s studies of lung morphology. But then I glance at some scholarly books (to avoid offending, I will not cite them here) that are supposed to be key references on the complete anatomy of Crocodylia and I frankly am left cold. While there is some superb work from the 1800s (Gadow, Fürbringer and others come immediately to mind), it is in the flat, often colourless style which the printing technology of that age imposed upon those great masters of anatomy. And while there is superb work by Romer, a tome on the Chinese alligator and a few others, again they tend to be limited to line drawings or spotty coverage of various anatomical systems. We need a visionary with both the scientific and artistic skill to make a subject that could seem dry or arcane become miraculous and accessible. I think you can guess whom I have in mind.
I can think of no one better suited to the ambitious goals and demands of the Ultimate Croc Anatomy project than Mieke Roth. She combines the attention to anatomical detail of a classical 19th century anatomist with the technical wizardry of a modern digital artist. She will have a supportive team of experts to ensure the content is exceptional and up-to-date. I’ll be one of them (and the crocs will come from my freezer), because of my enthusiasm for the project, and I’ve been helping to recruit others. So I urge you not just to join in her crowdfunding effort to carry out a very worthy and exciting project that the world can share, but also to share the joy I had in discovering her work by browsing her online collection of awesomeness. I predict that many of you will feel the power of her spell and become Mieke Roth fans, too, if you are not fortunate enough to be one yet.
A quick plug here for BBC Radio 4’s fourth episode of “Just So Science”, playing at 13:45 GMT today (this is the link). I was interviewed a few weeks ago for the show “How the Rhinoceros Got His Skin,” a la the classic Kipling tale. This series is revisiting Kipling’s tales in light of modern evolutionary science and evidence, whereas Kipling only had crude, Lamarckian or early Darwinian insight. Check out their earlier episodes on whales, leopards and armadillos– good stuff, and with real scientists. Richard Dawkins may appear again (EDIT: yep! Dawkins manifested) in this episode to provide some gravitas and evolution street cred, too.
And Freezersaurus gets a big plug! From the website: ” Rhinos and horses have much in common. John Hutchinson studies both, but just don’t ask to look inside his freezer.” 🙂 NOTE: I am not a vet (I am a biologist), and definitely not a horse specialist like others in our lab, but I do study horses a little, in a comparative context.
While the original Kipling story focuses on rhino skin, and the producers were interested because of my popular post here on rhino skin, we discussed other issues such as gait, fossil record, feet, and more. I owe thanks to rhino skin expert Dr Tobin Hieronymus for helping me bone up on the unusual skin of rhinos, which has a surprising amount in common with the tough hide of walruses, boars, some water deer, and a few other species. It’s not just normal thickened skin, as Tobin and others have shown. Anyway, I don’t want to give away what’s on the radio programme; afterwards I might embellish this post more with some rhino anatomy and mechanics facts.
Coincidentally, I’m receiving four white rhinoceros feet today from a zoo mortality. So it’s rhino-fest here!
I hope you like the show— please let me know what you think in the comments below! I really enjoyed listening to it, but I’d like to know what you thought.
White rhinoceros forelimb (left side), ready for dissection.
To kick off the New Year just right, our tetrapod team has a new paper in Nature, following up on last year’s Ichthyostega not-so-good-at-walking study (also see here). Yet this paper has a more anatomically descriptive — and also an “evo-devo” — twist to it. For brevity, I’ll let our press release tell the story, since I think it does a good job of it (like I always preach scientists should do, we worked with our PR company to write this together, so we’re happy with how the press release came out). In a nutshell, our study used some very fancy synchotron radiation techniques to image the 3D anatomy of the backbone in early land vertebrates. Our findings surprised even us, and ended up turning around palaeontology/comparative anatomy’s view of how the backbone evolved, giving us a new glimpse into our inner tetrapod.
Stick around for the videos at the end, which are the first four supplementary movies from the paper and are rather pretty (there are two more, for imaging/segmenting afficionados, but they are not as pretty or interesting for most of this blog’s readership). The final figure (Figure 1 from our paper) gives some extra visual context.
The paper is:
Pierce, S.E., Ahlberg, P.E., Hutchinson, J.R., Molnar, J.L., Sanchez, S., Tafforeau, P., Clack, J.A. 2013. Vertebral architecture in the earliest stem tetrapods. Nature, published online [here].
I should note that I’m just 3rd author, so I deserve only modest credit. But I helped. Even though no freezers were involved, or harmed, in the process.
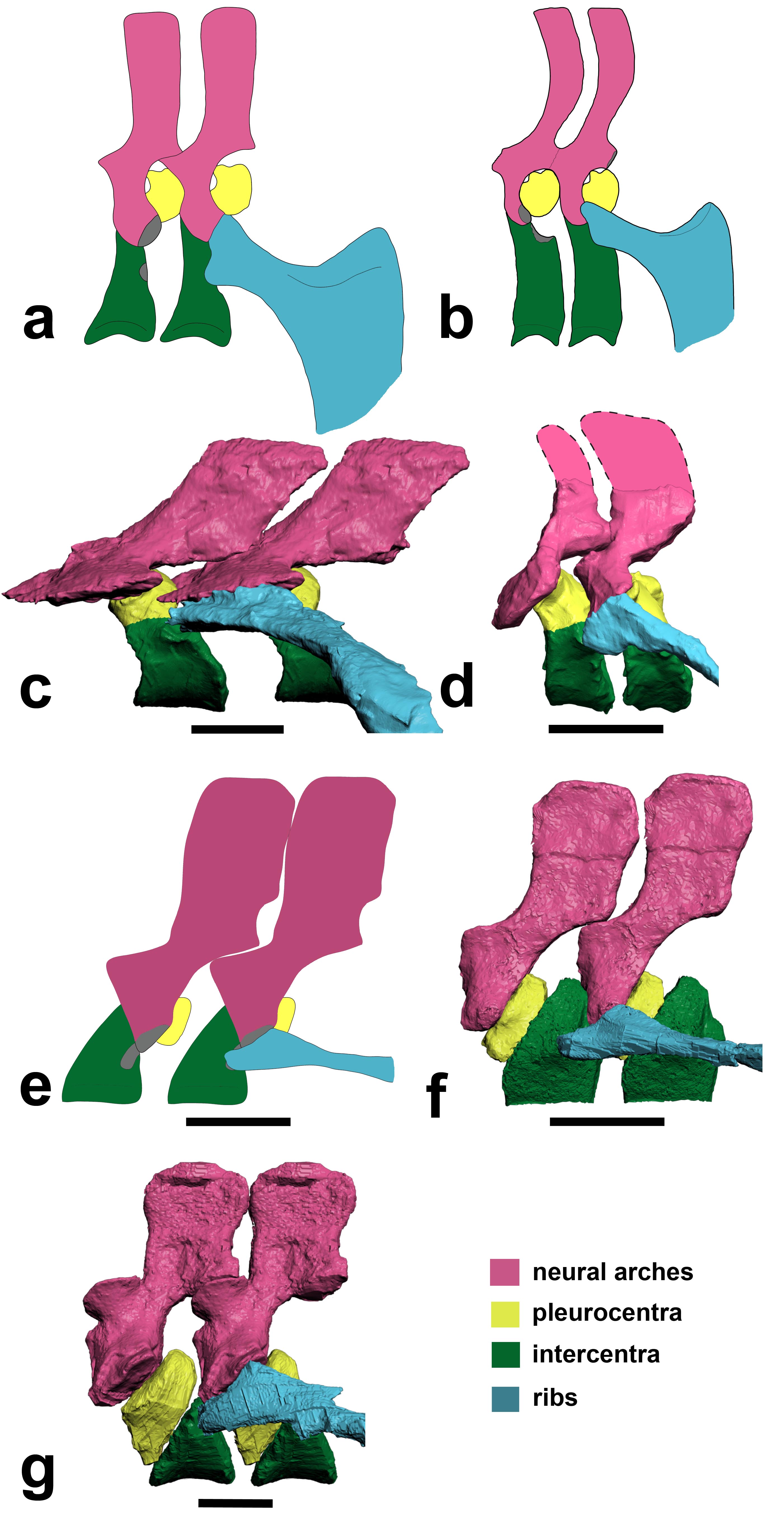
Above image: Julia Molnar‘s illustration of Ichthyostega showing anatomical changes of its spine from front to back, with neural arch/spine in pink, twin pleurocentra in yellow, and intercentrum in green. These four parts, three kinds of bones, made up the backbone of the first land vertebrates. These parts evolved in different ways in later animals, but formed one main bone in all living lineages of vertebrates.
RVC PRESS RELEASE:
Scientists reassemble the backbone of life using a particle accelerator
Research published today (Sunday 13 January 2013) in the journal Nature documents, for the first time, the intricate three-dimensional structure of the backbone in the earliest four-legged animals (tetrapods).
The backbone, also known as the spine or vertebral column, is a bony structure found in all tetrapods, along with other vertebrates such as fish. It is formed from many elements or vertebrae all connected in a row – from head to tail. Unlike the backbone of living tetrapods (e.g. humans), in which each vertebra is composed of only one bone, early tetrapods had vertebrae made up of multiple parts.
Lead author Dr Pierce says: “For more than 100 years, early tetrapods were thought to have vertebrae composed of three sets of bones – one bone in front, one on top, and a pair behind. But, by peering inside the fossils using synchrotron X-rays we have discovered that this traditional view literally got it back-to-front.”
For the analysis, the European Synchrotron Radiation Facility (ESRF) in France, where the three fossil fragments were scanned with X-rays, used a new protocol to reveal tiny details of the fossil bones buried deep inside the rock matrix.
Using this new technology, the team of scientists discovered that what was thought to be the first bone – known as the intercentrum – is actually the last in the series. And, although this might seem like a trivial oversight, this re-arrangement in vertebral structure has over-arching ramifications for the functional evolution of the tetrapod backbone.(see here for a now out-of-date image from Wikipedia)
Dr. Pierce explains: “By understanding how each of the bones fit together we can begin to explore the mobility of the spine and test how it may have transferred forces between the limbs during the early stages of land movement”.
But, the findings didn’t end there. One of the animals – known as Ichthyostega – was also found to have an assortment of hitherto unknown skeletal features including a string of bones extending down the middle of its chest.
Professor Clack says: “These chest bones turned out to be the earliest evolutionary attempt to produce a bony sternum. Such a structure would have strengthened the ribcage of Ichthyostega, permitting it to support its body weight on its chest while moving about on land.”
This unexpected discovery supports recent work done by the same authors that showed Ichthyostega probably moved by dragging itself across flat ground using synchronous ‘crutching’ motions of its front legs – much like that of a mudskipper or seal.
Dr Pierce adds: “The results of this study force us to re-write the textbook on backbone evolution in the earliest limbed animals.”
The next step, the researchers say, is to understand how the backbone aided locomotion in these early tetrapods using sophisticated biomechanical analysis.
These are rotating images of the anatomy, colour-coded, of the four species of early tetrapod that we examined for this study. Each shows the same basic pattern of having a “reverse rhachitomous” (pleurocentra in the front, intercentrum in the back; trying to think of a mullet joke…) anatomy. This is opposite the pattern that essentially all studies since famed evolutionary biologist/palaeontologist Edward Drinker Cope coined the term “rhachitomous” in 1878 have portrayed these and related animals as having. And this realization forces a re-examination of how the backbone structures first evolved in tetrapods and which parts (intercentra? pleurocentra? And where?) formed the spines of later animals.
For once, as authors we all felt that this finding really deserved the painfully hackneyed “rewrite the textbooks” label. It changes a lot of what we thought we knew about this classic evolutionary transition of anatomy. Check a vertebrate palaeontology/comparative anatomy textbook and you’ll likely find rhachitomous vertebrae and/or changes of pleurocentra vs. intercentra told in a way that we now are pretty sure is wrong.
You can also see the “sternebrae” (sternal elements; parts of the sternum that evolved independently in later land animals) in the first movie. This, to my knowledge, is by far the oldest such evidence. I know of ossified sternal plates in Early Permian mesosaurs like Stereosternum, but nothing earlier although perhaps in some synapsid I don’t know, or a basal diapsid of some kind? Chime in in the comments if you know of something I missed. Regardless, the sternebrae in Ichthyostega have nothing to do directly with those convergently evolved in lissamphibians, lepidosaurs, synapsids and archosaurs, although there may be some parallel developmental mechanisms involved and at least similar dermal tissues recruited into ossification patterns. Even so, these sternebrae are further evidence of how that taxon, at least, was beginning to make forays onto land, as they’d have helped it to support its belly on land and breathe.
The segmented PPC-SRµCT of Ichthyostegastensioi MGUH VP 6115 spinning in yaw and roll.
The segmented PPC-SRµCT of Ichthyostegaeigili MGUH VP 29017a spinning in yaw and roll.
The segmented PPC-SRµCT of Acanthostegagunnari MGUH f.n. 1227 spinning in yaw.
The segmented µCT of Pederpes finneyae GLAHMS 100815 spinning in yaw.
FIGURE:
Above: (a,b) How we used to think the vertebrae were composed in early tetrapods like Ichthyostega. (c) How we found that Ichthyostega‘s posterior thoracic vertebrae actually tend to look. (d) Ichthyostega‘s anterior lumbar vertebral morphology. (e) Acanthostega according to Coates’s important description. (f) Our revision of the anatomy of Acanthostega(anterior dorsal). (g) Our new interpretation of Pederpes‘s morphology, from a posterior dorsal. Focus on the yellow vs. green elements. In a,b and e they are in different positions (reversed) compared with our new versions in c,d,f,g.
To put the above figure and movies into broader context, check this Wikipedia image. We think the red/pink bones (pleurocentra) are in the wrong place relative to the blue ones (intercentrum); the ones currently there in this image actually belong to the vertebral unit behind that one, so the pleurocentra should be moved to the front (left end) of each unit. But also look down toward the bottom of the figure. Some of those vertebrae may need to have their blue/pink bits re-examined and interpreted, too. Is it turtles intercentra all the way down?
There you have it! Welcome to your new, revised, irradiated, reverse-rhachitomous inner tetrapod’s vertebrae. Propagation phase-contrast X-ray synchrotron microtomography FTW!!!!
Science media articles arising from this study–
I like to keep track of media stories covering our research, using this blog, so here are some of the stories about this paper. It’s funny… this was one of the most broadly important papers I’ve ever been on, but the coverage was relatively scant. It was too technical. We knew that would be a problem, and really had a hard time putting into words why the study was so surprising even to us! Most writers wanted the “how did the animals move?” angle, which was not what the study was about. I still feel that this angle was not even needed; the study (and again I take minimal credit for it) is exciting without it. To comparative anatomy and evo-devo specialists, anyway. Well, that’s science for you; sometimes it is just too hard to explain its value to the outside world, even when you feel its importance in your very spine… And the press coverage was not terrible by any means; no sour grapes from me. Regardless, we’re glad it has been well received by specialist researcher colleagues we’ve spoken to, and that matters a lot.
NERC’s Planet Earth (nice story from our funder)- “Scientists had fossil backbone backwards”
BBC online (the only story aside from NERC’s that did more than read the press release) “Tetrapod anatomy: Backbone back-to-front in early animals”
Discovery News online– “First Land Animals Shuffled Like Seals” (good, but is sort of mixing up our this study, our 2012 one and Ahlberg et al’s 2005 seal-analogue study; latter two were more about movement. As often happens, a lot of other media stories basically copied this one’s headline/angle.)
Discover 80beats– “Paleontologists Use 3-D Models to Rewrite Evolution” (also in “top stories”)
Popsci– “Particle Accelerator Reveals That First Land Animals Walked Like Seals”
Daily FMail (nice pics)- “Astonishing 3D images reveal the first four-legged land animals in amazing detail – and overturn a century of research” (wins longest headline award)
Red Orbit– “Study Reveals First Ever Images Of Early Tetrapod Backbone And How It Helped In Land Evolution”
Examiner.com– “X-ray study rewrites tetrapod backbone evolution (Photos)”
Business Standard– “Scientists recreate earliest quadraped’s backbone” (Proofread, editors! Quadruped.)
Geekosystem– “Early Land-Dwelling Animals Moved About Like Seals, Probably Didn’t Balance Balls on Their Noses” (scores some pts for humour)
…and the PR-copying, non-spellchecking fail of the week award goes to… Physorg! “Scientists reassemble the backbone of life with a particle acceleratorynchrotron [sic] X-rays”
Warming up the acceleratorynchrotron for our next study… 🙂