…a daily picture of anatomy!
Welcome to Freezermas! In the dead of winter, the WIJF blog jumps down your internet to deliver mind-warming science, and images, and evolution! To celebrate Charles Darwin’s birthday (204th = tomorrow Feb 12, 2013), I’m bringing you one Anatomy Vignette each day this week (we’ll see if I can manage the weekend or not)! Let’s do this!
Stomach-Churning Rating: 2/10; just bones; one picture of them, and then a lot of discussion of muscle anatomy but no pictures of it.
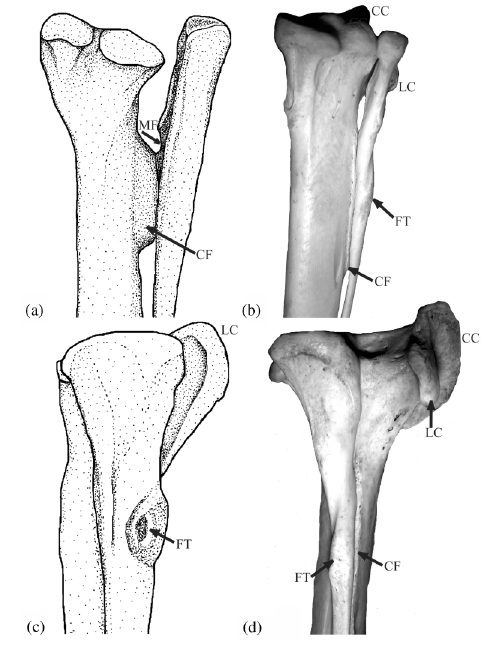
The above image comes from one of my old, somewhat obscure anatomy papers (link to pdf here), from 2003. It’s possibly the first figure I made, entirely by myself, that I’m sort of proud of. It doesn’t totally suck compared with some of my other attempts. I did the stippled line drawing on the left, and on the right is one of my first usages of a digital photo in a paper (digital cameras were finally up to the task around that time; I used my new Nikon Coolpix 900, if memory serves). It was a greatly improved figure over what I’d submitted for this paper originally, which was a rushed, half-baked manuscript for a SICB conference symposium on tendons. I’ll never forget one of the peer reviews of the manuscript, which said something like “the text of this paper is a joy to behold, but the figures are a horror.” They were right, and luckily the images in the paper I submitted changed a lot in revision. (I’m still embarrassed by the incident, though!)
Anyway, the picture is of the lower hind limb of two theropod dinosaurs: (a,c) an adult Tyrannosaurus rex, and (b,d) a wild turkey (Meleagris) from my personal collections of dissected-then-skeletonized animals (this turkey became a biomechanical model in a 2004 paper of mine, too!). In both cases we’re looking at a right hind limb; in (a) and (b) from a caudal/posterior/rear view, and in (c) and (d) from a lateral/side-on/profile view.
If you’re having trouble visualizing these bones in the real animal, check this T. rex skeleton in rear and side views and try to find these bones. You can do it! You might also want to look back at my paroxysmic outburst of love for knee joint anatomy.
The thicker long bone is the tibia (your main shank bone; or in a lamb shank, chicken drumstick, etc); the thinner outer bone is the fibula. Together with some smaller bones, for brevity we can call them the tibiotarsus — but only in theropod dinosaurs, or you will anger the freezer gods.
The labels show some cool anatomical features, as follows:
“CC” the cranial cnemial crest of the tibia (a projection of bone unique to the knees of birds);
“CF” the crista fibularis; or fibular crest; of the tibia (more about this below);
“FT” the fibular tubercle (insertion of the big hamstring/biceps muscle M. iliofibularis);
“LC” the lateral cnemial crest of the tibia (a big arching swath of bone that both birds and non-avian theropods like Tyrannosaurus have; the CC is just pasted on top of this in birds); and
“MF” which denotes a muscle fossa (depression) on the inner surface of the upper end of the fibula, which presumably housed a muscle (M. popliteus) binding the fibula to the tibia in earlier dinosaurs, but is vestigial in birds.
The CF, or fibular crest, is a feature that only theropod dinosaurs, among reptiles, develop like this. It evolved early in their history and thus was passed on to birds with other ancient features like hollow bones and bipedalism. It binds the fibula closely appressed to the tibia, making those bones act more like a single functional unit –and sometimes they even fuse together. The CF also transmits forces from the whopping big M. iliofibularis muscle’s insertion (the FT label) across the puny fibula onto the robust tibia. The MF once held a muscle that also helped keep those two bones together, but additionally it could have contracted to move them relative to each other a little bit, as in other living animals (many mammals and reptiles have a big M. popliteus and/or M. interosse[o]us). So these features all have a common functional, anatomical and evolutionary (and developmental; different story for evo-devo fans) relationship. By binding the fibula and tibia together, these structures helped early bipeds (the first theropods and kin) support themselves on one leg at a time during standing and moving, and also helped begin to reduce the limbs to lighten them for easier, faster swinging. So we can think of these features as specializations that helped theropod dinosaurs, and ultimately birds, get established as bipedal animals.
The CC and LC have a similar story to tell; for one, they are muscle attachments, again mainly for thigh muscles. And again, the LC dates back to early theropods (and some other dinosaurs had a version of it; usually smaller). These crests serve mainly as insertions for the “quadriceps” (in human/mammal terms) or triceps (in reptile/bird terms) muscle group’s major tendon, spanning from the pelvis/femur across the thigh and knee to this region. In birds, we call this structure of insertion the patellar tendon or (less appropriately) ligament. But dinosaurs had no patella, ever, so the triceps femoris tendon would be the proper technical term. Regardless, that crest (LC, and later LC too) helped the attached muscles to straighten the knee joint or support body weight during standing/moving, by giving them better leverage. So it would have been important for early bipeds, too, like the CF, MF and other features above. Your cnemial crest (tibial tuberosity) is pathetic by comparison. Don’t even look at it. Droop your knees in primate shame!
Bumps and squiggles on bones might seem puny details just for anatomists to study and describe in long, tedious monographs, but each is part of the great story of evolution, and each has a story to tell that fits into that story. Back in Darwin’s day, some of the world’s greatest scientists of the age (Richard Owen and Thomas Huxley being but two spectacular examples) pored over these seemingly innocuous features, and so they became part of nascent evolutionary theory even then. This week, I’ll be celebrating a lot of those details, which I still feel are important today, and the stories they help to tell.
Happy Freezermas! Sing it: “On the first day of Freezermas, this blo-og gave to me: a tibiotarsus with a CF and FT!”