Party time! Let the media onslaught begin! We’ve published a paper in Nature on the limb motions of Ichthyostega (and by implication, some other stem tetrapods). Since we did use some crocodile specimens from Freezersaurus (see below) in this study, I figured WIJF could cover it to help celebrate this auspicious event. Briefly. Particularly since we already did a quasi-blog on it, which is here:
However I want to feature our rockin’ cool animations we did for the paper, to squeeze every last possible drop of science communicationy goodness out of them. So here they are in all their digital glory. Huge credit to Dr. Stephanie Pierce, the brilliant, hardworking postdoc who spearheaded the work including these videos! Dr. Jenny Clack is our coauthor on this study and the sage of Ichthyostega and its relatives- her website is here. Also, a big hurrah for our goddess of artsy science, Julia Molnar, who helped with the videos and other images. Enjoy!
The computer model
The forelimb model
The hindlimb model
We used some of my Nile crocodile collection to do a validation analysis of our joint range of motion (ROM) methods, detailed in the Supplementary info of the paper, which I encourage anyone interested to read since it has loads more interesting stuff and cool pics. We found that a bone-based ROM will always give you a greater ROM than an intact fleshy limb-based ROM. In other words, muscles and ligaments (and articular cartilage, etc.). have a net effect of reducing how far a joint can move. This is not shocking but few studies have ever truly quantitatively checked this with empirical data from whole animals. It is an important consideration for all vert paleo types. Here is a pic of one of the crocodiles from the study, with (A) and without muscles (B; ligaments only):
I’ll close with Julia Molnar’s jaw-droppingly awesome flesh reconstruction from our model. Why Nature wouldn’t use this as a cover pic, I’ll never understand, but I LOVE it! When I first saw it enter my email inbox and then opened it to behold its glory, my squeal of geeky joy was deafening.
(edit: Aha! Fellow Berkeley alum Nick Pyenson’s group made the Nature cover, for their kickass study of rorqual whale anatomy, including a “new” organ! Well, we don’t feel so bad then. Great science– and a win for anatomy!!!)
This post will walk through the basic steps we take to do some of the major, ongoing research in my team. It comes from our lengthy project aiming to determine how elephant legs work at the level of individual muscle/tendon/bone organs. We need fancy computer simulations because anatomy, mechanics, physiology, neural control etc. are all very complex and not only impossible to completely measure in a living, moving animal but also extremely unethical and unjustified in the case of a rare, fragile animal like an Asian elephant. We want to do such complex things to test hypotheses about how animals work. For example, we want to estimate how fast an elephant could run if it wanted to, or why they cannot (or will not) jump or gallop like smaller mammals do— even as baby elephants (~100 kg or 220 lbs), which is an ancillary question we’re tackling. That’s cool basic science, and that’s enough for me. But the applications once such models and simulations are established are manifold– human clinical research now routinely employs such approaches to help treat “crouch gait” in patients with cerebral palsy, plan corrective surgeries, aid in rehabilitation strategies, and even potentially optimize athletic performance. Non-human research is pretty far behind this kind of confident application, because there are too damn many interesting non-humans out there to study and not many people using these approaches to study them (but it’s catching on).
Breaking up the monotony of the text with a baby elephant we met during our research in Thailand (Chiang Mai, here) in 2001. It was just a few days old and VERY cuddly and playful (chewing on everything!) but it’s mother did not want us playing with it so we only gave a quick hello.
I use the term model to refer to a simple abstraction of reality (such as an anatomically realistic computer graphic of a limb), and a simulation as a more complex process that is more open-ended and generally uses a model to ask a question (such as what level of extreme athletic behaviour a modelled limb could support). We use models and simulations to test how all the structures of the limb work together to produce movement. This also reciprocally gives us insight into the question, as I like to say it, of why is there anatomy? What is anatomy for? Why does it vary so much within so many groups and not so much in others? This can more easily be addressed by focusing on the consequences of a given anatomy rather than the more tricky question of why it evolved.
These approaches also can answer the frightening question of “Does anatomy really matter?” Sometimes it does not. And those “sometimes” can be impossible to predict- although sometimes they can be easy to predict, too. I think we are not at a point in the maturity of biomechanics/functional morphology to usually know a priori when either is the case. Many factors in addition to anatomy determine function, behaviour, or performance; that’s why; and biomechanics aims to unravel those relationships. A lot of anatomists, palaeontologists, etc. assume that form can be reliably used to predict function, but plenty of studies have shown already (and if you peer deeply into the details, it comes from first principles) that one cannot be sure without either measuring what anatomy is doing in a particular behaviour or estimating that function in a computer model or simulation.
Anyway, I’ve covered my perspective on this in a paper which you can read if you want to go into deep philosophical details of the science (and read me blabbering on more about this particular hobby horse of mine?). This post will proceed mostly with pretty images and simple explanations, although I welcome comments and queries at the end. As part of this post, I’ll try to give an idea of the timespans involved in doing the research. Some steps are quick and easy; others can take dauntingly long — especially to do well, without building a digital house of cards.
I’ll start, as my posts often do, with a deceased animal, and in this case it will again be an Asian elephant. Incidentally it is the same animal from the “Inside Nature’s Giants” series (see previous post).
Above: the hindlimb viewed from the rear, showing the medial (inside) region of the thigh skinned down to the superficial musculature. The hip is toward the left of the screen, and the knee is to the far right (whitish rounded area), with the shank (still bearing most of its grey hide) heading to the bottom right corner of the picture. Muscles pictured include ST (semitendinosus) and SM (semimembranosus); major hamstring muscles; as well as the thin, sheet-like gracilis, the straplike sartorius, and the massive adductors toward the top of the image.
When collecting data from dissections for functional analysis including computer models and simulations, we dissect the muscles one by one as we identify and photograph/sketch them, then remove them and do a suite of measurements to characterize how their form relates to some basic functional parameters. From the mass (weight) of the muscle and the length and angulation (pennation) of its fibres (bundled as fascicles) we can estimate what is called the physiological cross-sectional area (PCSA) of each muscle, which is known to strongly correlate with the force the muscle can produce. Different muscles have different PCSAs; for example check out these pictures of a long-fibred, lower-PCSA muscle and a short-fibred, highly pennate and high PCSA muscle:
Above: the long muscle fibres (bands running from left to right, somewhat diagonally from the bottom left corner toward the top right) of a hip adductor muscle in our specimen. The adductors are fairly simple muscles that run from the underside of the pelvis to the inside of the thigh (femur).
Above: the tensor fasciae latae (TFL; pretty sure of ID but going from memory) hip muscle of our specimen, cut open to show the short, angled fibres (each leading at around a 45 degree angle to attach onto a thick central internal tendon). The TFL is just out of view at the top of the screen in the whole leg anatomy picture above; it is on the front outer, upper margin of the hip/thigh and runs down to the outer side of the knee, invested with thick sheets of connective tissue (fascia).
The maximal isometric force (Fmax) of a muscle is computed as the PCSA times the muscle stress (force/unit area), which is fairly conservative in vertebrates. A square meter of PCSA can produce around 200-300 kilonewtons of force, or about 60,000 cheeseburger-weights (the standard unit of force on this blog). That’s a lot of quarter pounders! And an elephant has pretty close to that many cheeseburgers worth of leg muscle (around 150 kg mass, very close to a square meter of PCSA; total Fmax would be around 80,000 cheese-burger weights!). That much muscle is important because an Asian elephant like this one weighed 3550 kg or about 9000 cheeseburger-weights. So if all the muscles in one elephant hindlimb could push in one direction at once, in theory they could hold about 9 elephants aloft. However, as the picture above shows, they do not all act in the same direction. Furthermore, there are many other factors involved in determining how hard a leg can push, such as the leverage of the muscle forces versus the actions of gravity and inertia (mechanical advantage). All those factors, again, are why we need computer models to address the complexity. But the end result is that elephants cannot support 9 times their body weight on one hind leg.
Enough talk about cheeseburgers and enough possibly savory pictures of giant steak-like leg muscles. I don’t want to be blamed for hunger-induced health problems in my beloved blog-readership, dear Freezerinos! The above steps take about a week to complete for 2 legs of a big elephant, rushing against decomposition to try to get the best quality data we can. On to the digital stuff- let’s turn the geekitude dial up to 11 with some videos of computer modelling.
Our next step, often featured on this blog because I do this so often, is to take CT (and/or MRI) scans of the specimen that we wisely did before we cut it to bits, and use those to make a computer model. That’s the easy step; a scan nowaways takes me less than an hour to complete, including moving the specimen back and forth between the freezer and imaging centre. MRI scans can take quite a bit longer. Here is a CT scan of a similar hindlimb (right leg for the toes up to the knee, from a juvenile elephant; the above leg was too big for our scanner!). See what you can identify here:
And then here is a resulting computer model of the same animal (just knee down to toes), showing how we took each CT slice of even the muscles and turned them into fully or partially 3D digital organs, in our case using commercial software that makes this procedure (a step called segmentation) very easy:
The segmentation step for bones is usually incredibly simple; it can take anywhere from an hour to a day or so, depending on anatomical complexity and image quality. For muscles, this is harder because the images are often more hazy and muscles tend to interweave with each other, segue into tiny tendons, take sudden turns through bones or other narrow spaces, or even fuse with other muscles. So when we do this kind of musculoskeletal modelling, it gets pretty laborious, and can take weeks or months to finish.
Ahh, but once you’re done with the basic anatomy, the real fun begins! We take the 3D images of bones, muscles, etc. and import them into our biomechanics software. We use two packages: one commericial item called SIMM (Software for Integrative Musculoskeletal Modeling) for making models, and a nice freebie called OpenSim for doing simulations (although actually we’re finding SIMM is often better at doing both modelling and simulation for more unusual animals). Quite a bit more anatomical work is required to get the joints to move properly, then position the muscles in accurate or at least realistic 3D paths (depending on segmented image quality), then check the muscles to ensure they move properly throughout the joints’ ranges of motion, then import all the PCSA and Fmax and other data we need from dissections, then do a lot more debugging of the model… this takes months, at least.
But the greatest joy and pain comes in getting the biomechanics done with the models and simulations. You can get quite simple data out of the models alone; such as the leverages (moment arms) of individual muscles and how these change with limb joint position, across a gait cycle, etc… That’s pretty interesting to us, and can just take a few days to crank out from a finished model. Yet the ultimate goal is to do either a tracking simulation, in which we make the model try to follow forces and motions that we measured in experiments from the same or a similar animal (standard, harmless gait analyses), or a theoretical simulation, in which we set the model a task and some rules (‘optimization criteria’) and then set it to run (for hours, days or weeks) to solve that task while following the rules. In both cases, the simulations estimate the muscle activation timings (on/off and intensity) and forces, as well as the kinematics (motions) and kinetics (forces) of the limbs. Then we check the results, play around with the inputs (unknown parameters) as part of a sensitivity analysis, and re-run the analyses again, and again, and again… Here is a draft of a tracking simulation we’ve run for our elephant’s hindlimb:
Above: again, a right hindlimb of an Asian elephant. This test of our tracking simulation is replicating real experimental data (from motion capture and force platform analysis) of an elephant running at near its top speed; over 4 meters/second (>10 mph/16kph). The red lines are the individual muscles, and the green arrow is the ground reaction force, equal and opposite to the force that the limb applies to the ground. In a fast elephant that force can exceed the elephant’s body weight, so the muscles need to crank out kilo-cheeseburger-units of force!
And that’s about as far as I’ll get today. My team’s previous research (explore links for some fun videos) has shown that elephants can run about 7 meters/second (~15mph; 24kph) and that they have pretty poor mechanical advantage when they do run, so their muscles must have to work pretty hard (about 6 times more cheeseburger units in a fast run vs. a slower walk). So how do they do it? And what prevents them from going faster? What would happen if they jumped? What limits speed more; muscles, tendons or bones? Stay tuned. I’m still not sure how much longer this final step of the research will take… (presumably will precede the heat death of the universe by a long shot) But overall, the whole process when everything works nicely can take a year or so to do, proceeding from whole limbs to a simulated limbs.
As a final teaser, here is work we’ve done on using a different kind of model, called finite element analysis (FEA), to estimate how many cheeseburgers it would take to break an elephant’s femur (thigh bone), for example. How “overbuilt” are bones vs. muscles or tendons? This is still a poorly resolved question in biology. We’ve established some rigorous methodology for doing this, now we just need to see what answers it gives us…
(the colour shows the strain (deformation) in the bone in a simple bending experiment; “hot” colours are higher strain. The visualization of the strain is greatly exaggerated; in the real results they are barely visible, as bone only bends a tiny amount before fracturing)
Like other birds, ostriches are fluffy. Too fluffy for some anatomists– so fluffy, it’s hard imagining or estimating what they look like beneath all the feathers. A few years ago, we received an ostrich from a UK farmer. The male bird had been killed by a kick to the neck from another rival, and at the time was supposedly “Britain’s largest ostrich.” As the feathers were valuable to him, the farmer delivered the animal to us whole but plucked. I wanted to dissect it mainly to refresh my memory on ostrich anatomy while developing a biomechanical model of their limbs (see below). Taphonomy expert Jason Moore then buried it for his studies of how bodies decompose.
[Side note: ostriches and other ratites (flightless birds, members of the palaeognath group, whose evolution remains fascinatingly complex) are often brought up as uniquely dinosaur-like. That’s rather misleading; all birds are living dinosaurs, so all birds are descended from an ancestor that was equally ‘dinosaur-like’. What we see of them today is a snapshot that is biased by their recent evolutionary history. During their apparently multiple losses of flight, ratite birds increased in body size and “re-evolved” (or simply enhanced) some traits that were more marked in extinct dinosaurs than in the most recent common ancestor of living birds. Some of those more ‘primitive’ traits may be due to flightlessness, some due to large size, some due to their extreme running specializations; science hasn’t sorted all that out just yet. But the point is, ostriches and other ratites are far from the ancestral form that all birds sprung from, which was probably more like a small, flying tinamou-like animal. Their similarities are due to convergent evolution. And they’re still quite different from something like an “ostrich-mimic” dinosaur- which is a sad misnomer because it’s more that ostriches mimicked (in a naughty teleological sense) ostrich-mimic dinosaurs like Struthiomimusthan the other way around; the ornithomimosaurs did it first (Huzzah!). Ratites have just gone back, in some ways but not others (e.g. no long tail or large arms) to a superficially more primitive body form. There have been some wacky ideas to the contrary before, such as the idea that ratites evolved entirely separately from other living birds from different dinosaur stock, but they’re so discredited now by multiple lines of evidence that I won’t glorify them by spending time discussing each. This tangent has gone on too long and must die.]
Anyway, back to the plucked ostrich in question. My first look at it really stunned me. It was a powerful example of just how ‘dinosaurian’ most of the anatomy of living birds is, for reasons noted above. I’d never seen a naked ostrich and now I’ll never look at them the same again. Maybe you won’t, either…
First, some images of the animal once it was brought into our dissection room (which you might recognize from the great Inside Nature’s Giants documentary).
The device near the top of the screen is a digital scale; we were weighing the bird before we cut in…
Close-up view of the hugely muscular legs (each leg is around 25% of the animal’s body weight, and mostly muscle; about 50% more bulky than our legs), and the arms (shown more below).
129 kg weight sans feathers; not bad! That’s about 284 pounds for those folks still mired in the medieval Imperial system of units. 🙂
The swollen, bloody region just below the head (on the left above) is where the mortal blow struck. Ouch!
I love the hands of ratite birds. Yes, those are little claws attached to the three vestigial fingers (thumb/first finger at top, long middle finger, and tiny third finger bound to it). Darren Naish covered some of this in a previous post, and let’s not forget SV-POW’s excellent series of “things to make and do” involving various critters including ostriches.
Ostriches and I go way back. Here I am from my less bald immature postdoctoral days at Stanford University in 2002, dissecting a smaller (female, 65kg) ostrich for some biomechanical modelling (still mostly unpublished; aaargh!).
And yes, I had a third hand back then; later lost during a tragic dissection incident involving a battleaxe and a bottle of tequila. I don’t want to talk about that.
Ostrich packed for transport. Just barely fit in the trunk of my little 1993 Toyota Tercel (R.I.P.)!
Once we complete dissections. we put everything together in some fancy biomechanical computer models (a subject of a future post), resulting in a nice, 3D, poseable, anatomically-realistic model of the entire limb musculature, shown above. This is a right hindlimb in side view, with the individual muscle paths abstracted as red lines. More about this when it is finally published…
This is just a teaser showing off some of the cool external anatomy of ostriches-in-the-buff, and what we’ve done with the anatomical data we’ve gathered. I’ll do a post later showing what’s inside, which is also pretty amazing. Hope you enjoyed it!
Like I said, oh hai! Wow, I wasn’t expecting all of you here! Enjoy a random photo of some awesome anatomy, Boing Boing universe, and thanks for the nice story!!
Note that an elephant’s heart does not end in the stereotypical single apex (point) at the left ventricle. Elephants have a 2-pointed heart, with two large ventricles used to pump blood to the body AND lungs… who else has this feature? Ahh yes, their closest living relatives: seacows (manatees, dungongs; the Sirenia)! Probably a leftover swimming adaptation. Big, muscular ventricles (the darker purplish tissue to the left and right of the yellow-pinkish line of tissue running from top to bottom along the middle) are useful for pumping blood against resistance, such as when using the trunk as a snorkel while swimming.
The bottom of the heart is at the bottom of the screen; you’re looking straight at the front of the heart.
Human heart for comparison, from Wikipedia; scale relative to pic above is not too far off (elephant relatively a bit undersized):
For more info on elephant hearts, see here and here, and for the hardcore anatomists, here and here.
Good morning, Freezerinos! Here is a twin treat for you to puzzle over. Two things, perhaps rather squidlike at first glance, but not cephalopods. There is a conceptual connection between the two images. Can you identify both of these structures? Huge bonus points if you can identify the taxon they belonged to, but stabs at it are encouraged; there are clues in the images…
(labels have been removed to protect the innocent)
With the help of Heinrich Mallison of dinosaurpalaeo and his student Sebastian Marpmann (now having finished the Great Giraffe Deconstruction), I did a quick cleanup and reorganization of the big walk-in freezer we’ve all come to know and love as Freezersaurus. This had to be done because some of the big stuff was becoming a terrible obstacle to cross in order to get anything from inside; cue health-and-safety paperwork nightmares. And yes, the ice penis is now gone. End of an era…
All photos henceforth by Doktor Mallison:
Emerging from Freezersaurus; most big stuff removed (note our giraffe's metacarpus trying to escape, on right side)
The big stuff. Emus, elephant feet, too much giraffe (metacarpus continuing to sidle away), and horse legs.
Carnage remaining inside; needed to be taken out gradually and reorganized, to make more space on floor for intermittent human presence (i.e. walking in).
Elephant feet, mystery giraffe pelvis, oh my!
For ME!?!? Aww shucks, you shouldn't have. Reorganizing the small stuff on shelves. PhD student Mike Pittman makes a guest appearance, delivering crocodile vertebrae.
Sebastian poses with giraffe buddy. We emphasize the Buddy System when dealing with Freezersaurus; she is a treacherous hostess. Note that Sebastian has also cunningly halted the abscondence of the giraffe metacarpus.
Job done! Farewell Freezersaurus! You look mahvelous! Wall of archosaurs on the left; wall of synapsids on the right, and sundry giant mammals in the middle.
And so we finished, and so you’ve now had a very intimate look at Freezersaurus too! Don’t you feel lucky? 🙂
Without further ado, what’s up with this specimen from The Freezers? What is it, what animal etc?
(admin note, 28 Oct 2012: Mystery Dissections 2-3 do not exist, mysteriously. At the time, in the jumbled freezers of my mind, they corresponded to Mystery CT Slice(1) and Mystery CT Slice 2. But we can pretend that MD2-3 are just an eternal mystery of this blog, subject of numerous conspiracy theories that you are welcome to expound upon!)
There’s no better way to kick things off after a holiday than with a celebration of the Inside Nature’s Giants series, which I had a small part in early on, including these shots I took during the time they spent filming at the RVC >3 years ago (!?!?); most of these animals spent multiple holidays inside The Freezers:
Elephant arriving…
Elephant revealed
Private moment with elephant
Stunning emergence of The Guts
So you are impressed by the guts too, ehh? It was pretty amazing to watch it happen. The tension was intense- the animal had been dead for a while and was rather bloated. So cutting it open was a task gingerly taken…
Bloated elephant
RVC dissector Richard Prior stuck a scalpel in the upper abdomen when the time was right… the piercing whistle and the sulphuric odour silenced the crowd watching… and then quickly out came the guts.
Everyone was pretty amazed by the scale.
The guts just went on and on…
Not a 1-person job by any means.
Spreading them out to see the whole GI tract.
I waited patiently and watched the show filming; what a great, professional crew. Then I got to take the legs away for our research.
But not just elephants, no sirree! The Windfall Films/ING team filmed giraffe, crocodile and big cats episodes (4 total) at the RVC too; a crazy period of a few weeks (including a major blizzard that hit us during the croc filming) in 2009. Some of the stars follow:
…and I’m rather fond of that tiger’s neck– check out the hyoids (roaring/tongue apparatus in throat; bottom of movie)!
…and here is the adult Nile crocodile’s head after scanning
…and another view of that big Nile croc, just because I like how this reconstruction turned out
…and here’s one of the small (~1m long, 10kg) juvenile Nile crocodiles from the show, with a pilot CT scan showing the skeleton nicely- and possibly a last meal or stomach stone on the left side of the abdomen (bright white blob; I need to check this now that we’ve dissected it)
Foetal giraffe; stillborn; from the show, in process of dissection in our lab to measure its limb anatomy. Trust me, it looked –and smelled– better on the inside than it did from the outside. Eew.
How most of the specimens from the first 4 episodes ended up after all dissection was done (part of my/RVC’s collection of skeletons). Sadly, I did not get great photos of the 3.7m Nile crocodile or the two giraffes before they were reduced to bits, but I do have the skeletons and CT scans.
Giving a tour (including The Freezers) to A Certain Esteemed Visitor.
(Another) Gratuitous shot with one of the sweet old Red Kangaroos at Alma Park Zoo near Brisbane, Australia. Experiments on hopping we did there will be briefly featured in the new Inside Nature’s Giants show on Channel 4, 16 April @2000- details at http://t.co/SkjsMeVC.
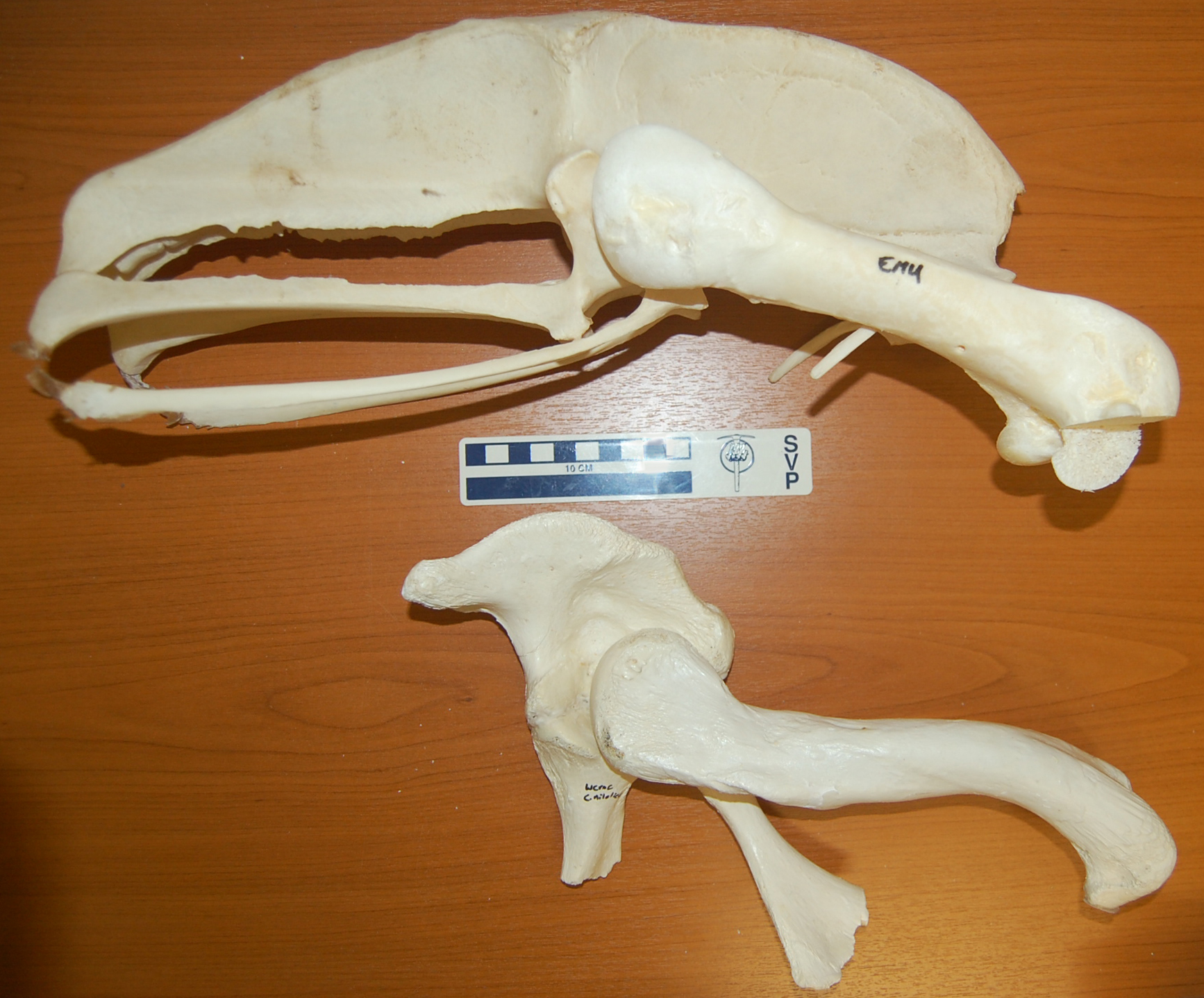
I’m off soon for a sunny break on the beaches of Morocco, but as an Easter gift to you, my (admirable, sagely, few, beautiful) readers, here is an image of two specimens, formerly from my freezers, to consider:
Both are the right pelvis (hip bones; crocodile pelvis is a bit broken toward the bottom) and femur (thigh bones) of living archosaurs– a 27kg emu above, and a 278kg nile crocodile below. The head would be toward the right side of the picture. A tenfold difference in mass between the bird and the crocodile, and yet some of the dimensions are so similar in both of them (femur length etc.), or so vastly different in the bipedal runner vs. the quadrupedal not-so-fast-runner (much bigger pelvis for leg muscle attachments in the former).
This image says it all. It is why I study the evolution of locomotion in land animals. It is why I am so fascinated by the transition from vaguely crocodile-like early archosaurs to modern birds by way of earlier dinosaurs. Anatomy, size, mechanics, behaviour, phylogeny… the photo captures all the facets of why I am so enraptured by research in this field.
It also might evoke thoughts of what features are expected in a terrestrial vs. aquatic animal, and thoughts of how some numbskulls still think big dinosaurs lived in the water (no I will not link to the execrable story from BBC today that I am thinking of!)…
I hope you appreciate it, too. Have a freezer-burn-free holiday period, folks!













")
")














")
")