…a daily picture of anatomy! And today it is six picture-facts; doo-raa-dee! ♫
Welcome back again, again, and again (gasp, pant)— and again (exhausted howl) to Freezermas!
And Happy World Pangolin Day!
Stomach-Churning Rating: 4/10; pretty tame images of anatomy today– but 9/10 if you consider how vile a practice it is to eat pangolins.
Much like rhinoceroses are, pangolins (“scaly anteaters”) are threatened with extinction across Africa and Asia largely because tradition holds that they have magical skin. It comes down to that. It’s simply pathetic.

Sunda pangolin, Manis javanica; from Wikipedia. It’s not a pig in a convenient artichoke-like wrapper. It’s a precious, rare creature.
To make matters worse, pangolins are smaller than rhinos and covered in the tough armour that makes them so desirable, and hence they are more portable and easy to hide. They also are thought to taste delicious — or just have the social cachet that it is a sign of affluence to be able to afford to eat them — to some people, especially from some southeast Asian cultures. Habitat loss/growing populations/deforestation/climate change aren’t helping, either.
Around 60,000 pangolins were illegally smuggled or otherwise slaughtered for human uses in 2012 worldwide; contrasting with 668 rhinos in South Africa that year (perhaps 2,000 worldwide?); so the scale of the problem is immense. Smaller-bodied pangolins will be more numerous in the wild than large, wide-roaming rhinos, but the drain on those numbers is obviously not sustainable. Sometimes pangolins are smuggled alive, a cruel practice that delivers them fresh but in a poor welfare state at the point of sale, compounding the urgency to turn the tide of exploitation.
Please take the time today to lend your hand to a good conservation group. Learn about the crisis facing pangolins (e.g. this recent article; and this video) and speak out about it. Of course, don’t eat pangolins, either.
Let’s not let humanity fail in its moral imperative of stewardship.

My photo of a pangolin body and skeleton, from the University of Cambridge Museum of Zoology’s exhibits.
In celebration of World Pangolin Day, for today’s Freezermas we have six impressive facts about pangolin anatomy. Much like rhinos, these are animals we don’t know as well as we should. I’ve never had one in my freezers, and would feel a bit weird if I did, since I find them so adorable, but they do have fascinating anatomy, natural history and evolutionary heritage. All the more reason to preserve them as they should be: alive and with the freedom they deserve.
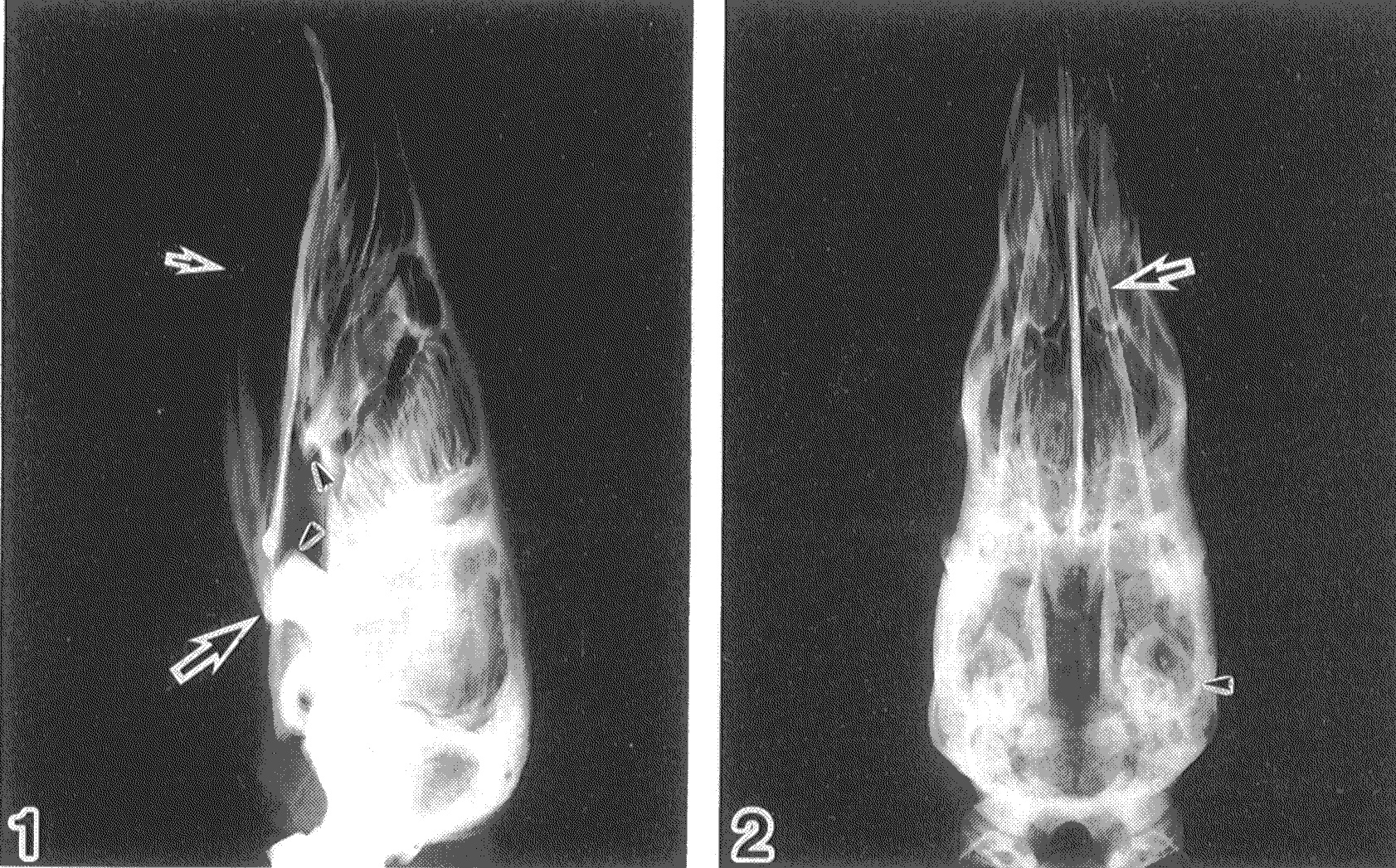
Pangolin Fact 1: Pangolins have highly modified skulls with myrmecophagous adaptations-– these are specializations for eating arthropods (especially ants/termites): toothless, tubular snout, reduced mandibles, and more– shown below.

X-ray of Malayan pangolin (Manis javanica) skull in side(1) and top(2) views, modified from Endo et al., 1998. The small arrow denotes the V-shaped, splint-like mandible, and the large arrow is directed at the jaw joint (zygomatic process on the temporal bone). The zygomatic arch, crossing from the jaw joint toward the front of the upper jaw (maxilla), is incomplete, so there is no bony bridge across the cheek as in many mammals. The large masseter and temporalis muscles run across this region, forming a more flexible, muscular cheek involved in feeding. Some nice labelled skull photos are here.
EDIT: Aaagh! Of course I should have checked Digimorph, which has a kickass CT scan/movie of the skull. Play with that; hours of anatomy-tainment!
Pangolin Fact 2: Pangolins have long tongues whose attachment extends way across the breastbone.
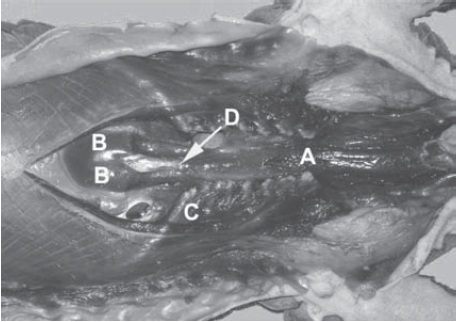
Tongue anatomy of a Malayan pangolin, from Prapong et al., 2009. This shows the chest region in ventral view (head is to the right side), with the main body of the sternum removed. A indicates muscles forming a sac around the tongue base; B is where the tongue finally inserts on the sternum/xiphoid processes; C is the ribcage; D is the xiphisternal joint (middle of the sternum parts).
Your tongue, even Gene Simmons‘s, just extends a little ways down your chin. It is, however, a common misconception that a pangolin’s tongue is longer than the animal. It can’t be longer than the distance between its sternal origin and the tip of the snout, so it might extend up to 40cm out of the mouth when fully extended in a large pangolin. Around 1988, there was the scientific misconception that the tongue extended way back to the pelvis (hips) or stomach. This is not true according to the latest literature I’ve read (e.g. in caption above), but is widespread in pangolin information on the internet. If someone has secondary confirmation of this either way, I’d love to see some concrete evidence.
EDIT: This image of a dissected pangolin fetus indicates a quite long tongue, maybe even long enough to attach near the pelvis, although that site agrees that there is no pelvic attachment. The latter site also depicts a fascinating cartilaginous sheath for the tongue. The misinformation about pangolin tongues does make me wonder: perhaps there is a lot of diversity in tongue attachments/lengths among the 8 pangolin species? Who knows.
Pangolin Fact 3: Pangolins have toughened, keratinized stomach linings.

Click to embigitate. Histology of the stomach lining in Manis tricuspis (modified from Ofusori et al., 2008), showing layers of keratinized stratified squamous epithelium (thick stomach lining). These layers seems to act as a protective coating against the rasping, chitinous exoskeletons of the ants and termites that are consumed, helping to reduce the risk of ulcers while reportedly eating up to 200,000 ants/per night. There is also an increased preponderance of elastic and collagenous fibers in layers of the stomach, helping it to expand to enclose many ants from one feeding.
Pangolin Fact(ish) 4: Pangolins are not closely related to other ant-eating living mammals, but to carnivores.

Eurotamandua; a possible fossil relative of pangolins from the early Eocene (Messel, Germany); image from Wikipedia.
Together, the eight living pangolin species are remnants of the group Pholidota, which has a respectable fossil record– particularly considering that they lack teeth, which are often such a diagnostic feature for mammalian fossils. Controversy persisted for many years about whether they were related to anteaters, sloths and armadillos (Xenarthra) within the group Edentata, along with possibly aardvarks (Tubulidentata) and other digging and/or myrmecophagous animals. There has also been controversy about some fossil mammals and their relationships, including Eurotamandua (above) and the Palaeonodonta— the latter seems to be approaching a consensus, though, as an extinct sister group to Pholidota.
Nonetheless, the main features that were once thought to unite ant-eating mammals as close relatives now seem to be a prime example of convergent evolution. Xenarthans are definitely closely related to each other, but aardvarks are afrotherians (closer related to hyraxes and elephants), and pangolins seem not to be closely related to either of those groups.
More conclusively, with the addition of genetic data, it has emerged that Pholidota is most closely related to Carnivora (mongooses, dogs, cats, bears, pinnipeds, etc.) among living mammals. A good example of this conclusion is the very recent paper by O’Leary et al. in Science. Furthermore, this image shows a nice example of such a phylogenetic result. This relationship with Carnivora raises fascinating questions about the tempo and mode of the evolution of all their digging/ant-eating specializations- when, where and how did they become so much like other ant-eating mammals?
Pangolin Fact 5: Pangolins have many digging (fossorial) and climbing (scansorial) adaptations, especially in their forelimbs.


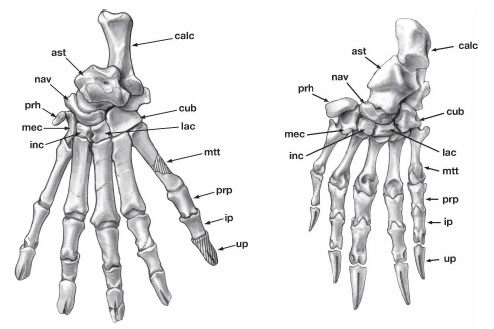
Click to embignify. Above (modified from Gaudin et al., 2006; by Julia Morgan-Scott): drawings of hind feet of (left) the Eocene fossil pangolin Cryptomanis and (right) Manis; Below: line drawings of front feet (from Humphrey, 1869) showing the convergent evolution of digging/climbing hands in (left to right) pangolins, an anteater (2-toed; Myrmecophaga), Ai (3-toed sloth; Bradypus) and Unau (2-toed sloth; Choloepus).
A striking feature of pangolin claw bones (unguals); evident above; is their characteristic fissured anatomy (split ends), which even the fossils have. This probably is how they develop strong, keratinous digging claws that remain anchored to those bony claw cores. If you look really closely, you may be able to see the fused scaphoid and lunar (scapholunar) bones of the wrist in the manus of Manis. Cryptomanis (above left) had more climbing specializations than living pangolins; is this how pangolins first evolved, and then later added more ant-eating features? This makes sense in terms of their phylogeny (above), as they are related to primitively climbing carnivores.
Other possibly digging/climbing-related features characteristic of pangolins include the loss of a coracoid process on the pectoral girdle, and curious enrolled zygapophyses (joints) on the lumbar (lower back) vertebrae — the functional significance of the latter feature is almost unstudied, but is reminscent of the complex xenarthrous vertebrae that gave Xenarthra their name (see above and this past post). A nice photo of a pangolin ribcage/vertebrae is here. There is an exceptional page on pangolins and their once-thought-to-be-close relatives among Xenarthra here, with lots of anatomical detail.
A feature that first got me scientifically curious about pangolins in my research is the presence of “predigits“- prepollex and prehallux- in their hands and feet (“prh” in upper left two figures). Many mammals have these, and some have expanded them into larger structures like the “sixth toes of elephants” (hence my interest), but precious little is known about their evolution or function in many other groups.
Pangolin Fact 6: Pangolin skin armour, like rhinoceros horns, is just modified skin (hair/epidermis) keratin; shaped into imbricating scales.


That apocryphally “magic skin”. Images from Wikipedia: closeup above, and below it a suit of armour made from those scales– coated in gold and given to King George III in 1820.
These scales, the double-edge sword of pangolins (both protecting them in nature and making them desirable in part of the human world for silly reasons), form as pangolins grow. In the fetus they are still soft, making fetuses more of a delicacy in some Asian cultures. Much like the stomach lining described above, the skin is formed from keratinized, stratified squamous epithelium– much more densely formed than in our skin, but more like in our fingernails. Asian pangolins, unlike African species, may have some more normal hairs beneath the scales, too.
There is no convincing evidence that the scales are any more healthy to eat, in any form, than your own fingernails, dead skin, or hair. Given the ready availability of the latter to any humans, we’re all wearing, and growing, our own goldmine…
I’ve barely dug into the fascinating biology of pangolins. I haven’t talked about their bipedal locomotion, much as it fascinates me, because we know next to nothing about that. I’m not aware of good scientific studies on their prehensile tail, either. A great page on pangolin biology, with a focus on reproduction and anatomy, is here. A lovely illustration and discussion of the convergent evolution between anteaters and pangolins is here. Awesome photos and facts are here. More about pangolins’ plight here, and very thoroughly here.
If you have favourite links to more material, or want to provide more information or especially questions, don’t hold back and experience painful pang(olin)s of remorse– chime in in the comments below!
Happy World Pangolin Day! Visit these great pages, please! Here, here and here! And…
Happy Freezermas! Sing it: “On the sixth day of Freezermas, this blo-og gave to me: one tibiotarsus, two silly Darwins, three muscle layers, four gory hearts, five doggie models a-and six facts of pangolin anatomy!” ♪♫
Read Full Post »