If you want to see a new/reinvigorated, exciting direction that palaeoart is headed, check out the All Yesterdays book by Conway, Kosemen and Naish. This review is fully cognizant that I’m late to the party of hailing this book as part of a palaeoart renaissance. I confess I haven’t read any of the many reviews of this book; I just know it is highly regarded and popular, from excitement on social media sites I frequent. So if my review covers ground others have too, so be it; it’s purely my own thoughts but I expect that mine fall in line with many others’. I’m reviewing the book on this blog because I love the interface between science and art (which is very important in anatomy), and because anatomy, and how one infers it when it is unknown, is the fundamental theme of the book.
You can buy All Yesterdays for around £18 (ASIDE: oddly, used copies (“May not include CD, access code, or DJ”– ???) are around £42 on the same site; perhaps those are artist-signed??? I have no idea!). It is a good deal at that price. While you’re at it, get “Dinosaur Art: The World’s Best Paleoart” by White et al. (including Conway) for a similar price. My review will return to some comparisons between these two books, released just a few months apart.


All Yesterdays is about not only how we reconstruct dinosaurs and other prehistoric animals, but also about thinking outside-the-box in the ways we reconstruct them and thereby bucking some recent clichés and tropes. Some of those outside-the-box ideas might seem ludicrous, and some probably are. But one of the main points of All Yesterdays is that there is plenty about extinct animals, and even living animals, that we don’t know, even though the field of paleoart has matured into greater scientific rigor than in the days of Knight, Zallinger and others (1920s-1950s). There is a focus on uncertainties about integument (e.g. feathering, spines, colour/patterning, body contours) and behaviour (e.g. avoiding stereotypes like perpetually aggressive predators and frightened prey animals– amen to that!). And the capstone of the book, which in some ways I loved the most, is turning the issue on its head and pretending that we only had skeletons of extant animals, then proceeding to reconstruct those animals (elephants, whales, horses and swans stand out prominently in this section; some of these are shown below). I wish more scientists in my general area would practice this; e.g. validation of a methodology used to reconstruct extinct animals in science.
The ‘speculative zoology’ of All Yesterdays deserves favourable comparison to one of my favourite science-art books, 1981’s After Man by Dougal Dixon. I fell so in love with that book as a 10-year-old that I wouldn’t let my parents return it to the library and I made them pay the hefty lost-book-fee (yes, I was a little bastard!). I still have it, too. (Sorry, Sequoya Branch Public Library of Madison, WI!) Likewise, the whimsy of the Rhinogrades is evoked by this work, and of course Tetrapod Zoology blog readers will be no strangers to it, either.
The book begins with a clear, succinct (7 page) summary of the history and science of reconstructing animals, with a focus on paleoart’s approach rather than science’s. I would have found it interesting (but space constraints presumably precluded) to feature more of the interface/parallels with scientists at the same time, such as the careful reconstructions of musculature in A.S. Romer’s masterful work in the 1920s (e.g. below), or later efforts by palaeontologists like Alick Walker and Walter Coombs. Many of these luminaries sought not to reconstruct animals for artistic purposes, but for almost purely scientific ones: to understand what skeletal anatomy meant in terms of broader biology (e.g. comparative anatomy) and phylogeny (e.g. origin of birds or archosaur evolution). The quality of their own artistic representations as well as scientific interpretations varied a lot. Indeed, sometimes the choice of model organisms (crocodile for Romer; lizard for Walker; birds in the post-1960’s) reveals much about the author’s preconceptions about phylogeny, marshalled towards a favoured hypothesis (e.g. a crocodile origin of birds for Walker; or an avian origin amongst dinosaurs for Bakker, Paul and others), rather than a circumspect assessment of all relevant evidence.

Figure 6 from Romer, 1923; very crocodylian T. rex right hindlimb muscles.
But eventually the “model organism” approach to reconstructing extinct animals gave way to the extant phylogenetic bracket; very popular today; which itself is an adaptation of the outgroup method for polarity assessment in phylogenetic systematics (cladistics). I am sure many modern paleoartists explicitly consider the “EPB” in their reconstructions, although this leaves many ambiguities (e.g. integument of crocodiles and birds being totally different!) that they must overcome, whereas scientists might just give up. This interface of art and science is part of what make palaeontology so enjoyable.
The EPB mindset has been a big step forward for evolutionary morphology and palaeontology, but still some of the greatest questions (e.g. what were the actual sizes, colour patterns, or behaviours of extinct animals? How did novelties arise and which novelties did dinosaurs have that extant relatives lack?) are left ambiguous by the EPB. This is because either the EPB itself is ambiguous (crocodiles or other taxa do one thing; birds do something altogether different), or because features leave no osteological correlates (e.g. muscle/tendon/ligament scars) on fossils that can be compared with the EPB. This quandary leads to the fun side of this book– filling in the huge gaps left by both basic anatomical interpretation and the restrictions imposed by the EPB, and then playing with the frontiers of anatomical, behavioural and ecological reconstruction, using informed speculation.
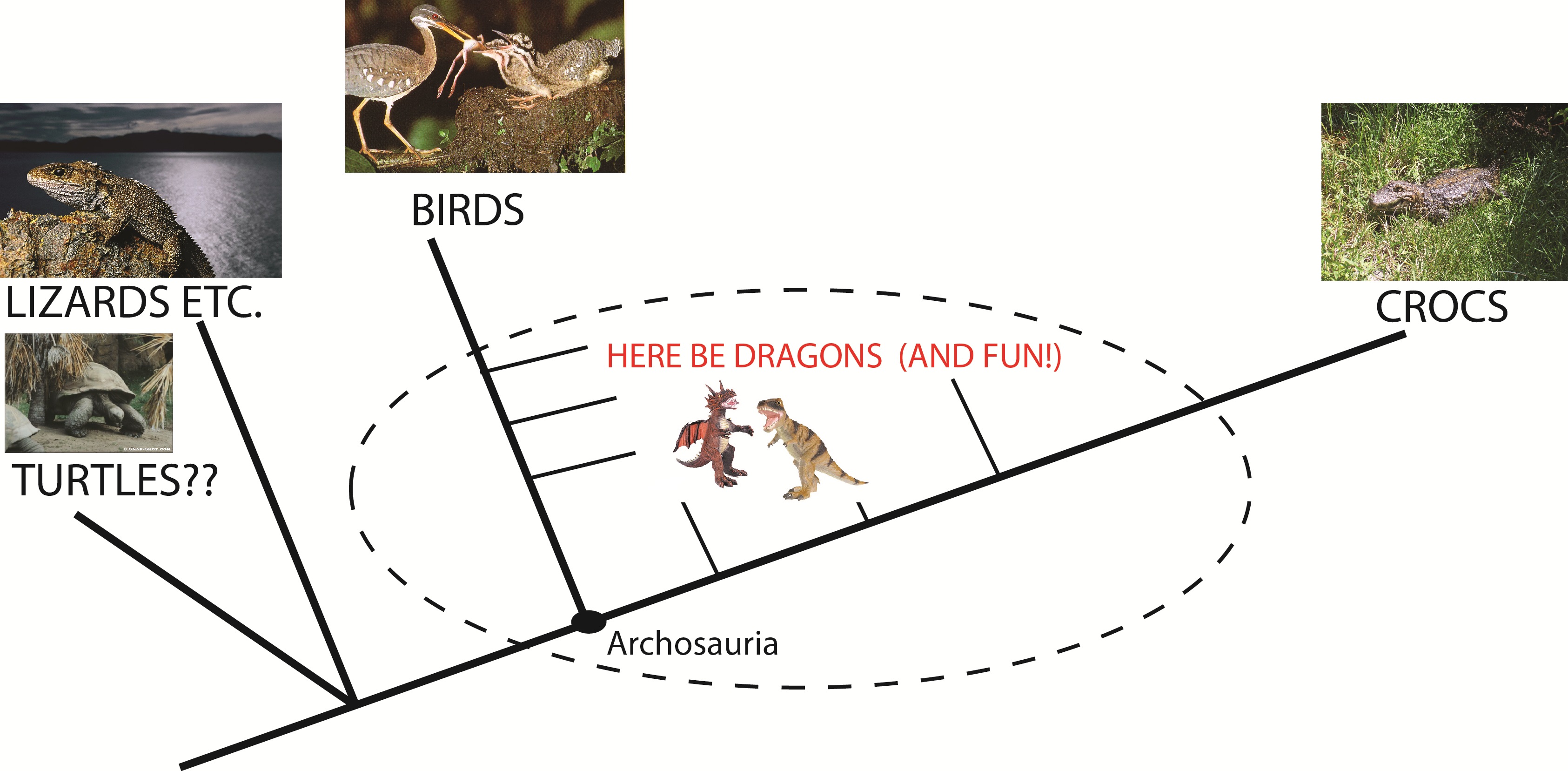
The extant phylogenetic bracket for archosaurs.
In addition to the startling, bizarre “All Todays” reconstructions at the end of the book, the highlights for me were the camouflaged Majungasaurus and plesiosaur, the “feathered mountain” (below) of a therizinosaur (can anyone illustrate a plausible therizinosaur and make it normal and boring? I wager not!) and the neck-swinging elasmosaurs engaged in “honest signalling” of their fitness. Many of the illustrations riff on notions popular in the modern palaeo-zeitgeist (and subject of many conversations at conferences, or even publications), such as evidence for the spiny integument of some ornithischians, fat ornithopods, Microraptor of somewhat-known-colouration, and so on. But plenty of other images riff on a “well why not?” theme, challenging the viewer to consider that extinct animals could have many surprises left in store for us with future discoveries, or else plausible features that we’ll never know of but might seem laughable or unfashionable to illustrate now. Each image has text explaining the logic behind it- this is not just a montage of pictures. This is a thinking person’s book- you should buy it for rumination, to challenge your preconceptions, not to have a flashy coffee table book. It’s not eye candy — it’s more like brain jerky.

John Conway‘s mountain-of-feathers therizinosaurs: eerily beautiful.
I think this is a bold, fun (re)new(ed) direction for palaeoart. There’s always a place for rigorous, conservatively evidence-based, by-comparison-almost-uncreative scientific illustration of extinct organisms. The World’s Best Paleoart presents loads of this, often in vividly colourful, photo-realistic, lavish, glossy detail, whereas the approach in All Yesterdays tends toward a more soft, matte, informal style including sketches or abstractions, toning down the serious and intense (even cluttered?) approach that can characterize modern palaeoart, including The World’s Best Paleoart. Sometimes those reconstructing life of the past (scientists included!) may emphasize that detailed realism too much and lose some of the joyful playfulness that palaeoart can revel in, at its best, most inspirational or thought-provoking. The former style might be considered the more “safe” or technical practice; the latter more risky or unconstrained.
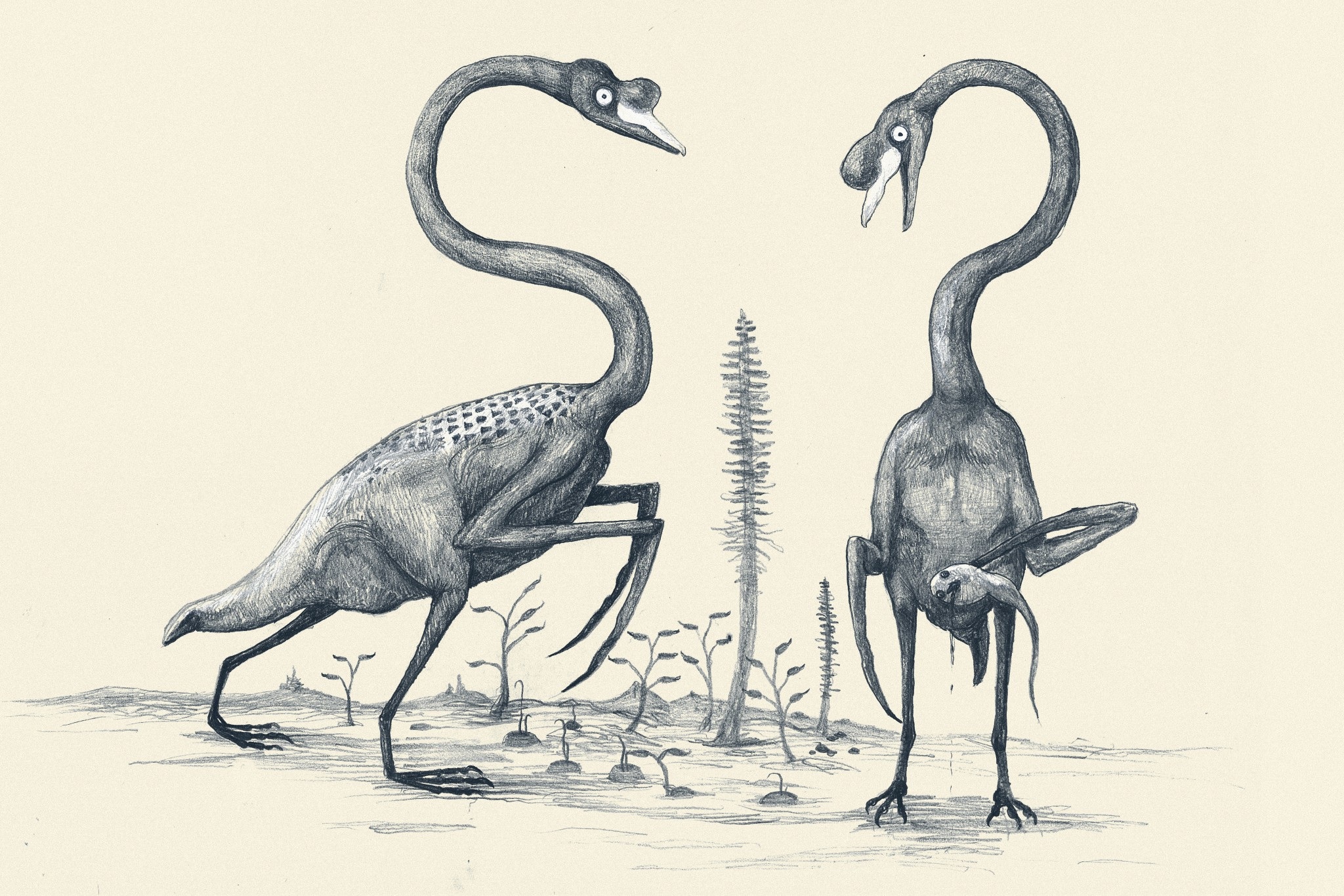
C.M. Kosemen‘s “All Todays” swans, with tadpolefish, might haunt your nightmares.
I’m not casting negative judgement on either style; both are absolutely wonderful — and valuable. I love both books! I’m glad we’re in a new age where the fun is waltzing back into palaeoart, that’s all. All Yesterdays doesn’t just waltz, either. It pounces into your field of view, wiggles its rainbow-coloured, mandrill-esque ankylosaurid bottom at you with a cheeky grin, and proceeds to make you smirk, be bemused, and even gasp at its adventurousness in rapid succession as you turn its pages. At 100 pages it doesn’t overstay its welcome either– that kaleidoscopic thyreophoran rump cartwheels off into the sunset at an opportune moment.

You won’t forget C.M. Kosemen‘s “All Todays” elephant.
If All Yesterdays makes someone uncomfortable with its swashbucklerish daring, they’re probably taking palaeontology way too seriously– and maybe missing not only some good fun, but also some potential truths. Dogma is a terrible thing, and All Yesterdays slaughters it with delightful relish. Bring on the next installment! If you have All Yesterdays too, what’s your favourite part? Or if you don’t have it, I’d be happy to answer queries in the Comments.