Deck the ‘Nets With PeerJ Papers— please sing along!
♬Deck the ‘nets with PeerJ papers,
Fa la la la la, la la la la.
‘Tis the day to show our labours,
Fa la la la la, la la la la.
Downloads free; CC-BY license,
Fa la la, la la la, la la la.
Read the extant ratite science,
Fa la la la la, la la la la.
See the emu legs before you
Fa la la la la, la la la la.
Muscles allometric’ly grew.
Fa la la la la, la la la la.
Follow the evolvin’ kneecaps
Fa la la la la, la la la la.
While we dish out ratite recaps
Fa la la la la, la la la la.
Soon ostrich patellar printing
Fa la la la la, la la la la.
Hail anat’my, don’t be squinting
Fa la la la la, la la la la.
Dissections done all together
Fa la la la la, la la la la.
Heedless of the flying feathers,
Fa la la la la, la la la la♪
(alternate rockin’ instrumental version)
Stomach-Churning Rating: 5/10: cheesy songs vs. fatty chunks of tissue; there are no better Crimbo treats!
Today is a special day for palaeognath publications, principally pertaining to the plethora of published PeerJ papers (well, three of them anyway) released today, featuring my team’s research! An early Crimbo comes this year in the form of three related studies of hind limb anatomy, development, evolution and biomechanics in those flightless feathered freaks of evolutionary whimsy, the ratites! And since the papers are all published online in PeerJ (gold open access), they are free for anyone with internet access to download and use with due credit. These papers include some stunning images of morphology and histology, evolutionary diagrams, and a special treat to be revealed below. Here I’ll summarize the papers we have written together (with thanks to Leverhulme Trust funding!):
1) Lamas, L., Main, R.P., Hutchinson, J.R. 2014. Ontogenetic scaling patterns and functional anatomy of the pelvic limb musculature in emus (Dromaius novaehollandiae). PeerJ 2:e716 http://dx.doi.org/10.7717/peerj.716
My final year PhD student and “emu whisperer” Luis Lamas has published his first paper with co-supervisor Russ Main and I. Our paper beautifully illustrates the gross anatomy of the leg muscles of emus, and then uses exhaustive measurements (about 6524 of them, all done manually!) of muscle architecture (masses, lengths, etc.) to show how each of the 34 muscles and their tendons grew across a more than tenfold range of body mass (from 6 weeks to 18 months of age). We learned that these muscles get relatively, not just absolutely, larger as emus grow, and their force-generating ability increases almost as strongly, whereas their tendons tend to grow less quickly. As a result, baby emus have only about 22% of their body mass as leg muscles, vs. about 30% in adults. However, baby emus still are extremely athletic, more so than adults and perhaps even “overbuilt” in some ways.
This pattern of rapidly growing, enlarged leg muscles seems to be a general, ancestral pattern for living bird species, reflecting the precocial (more independent, less nest-bound), cursorial (long-legged, running-adapted) natural history and anatomy, considering other studies of ostriches, rheas, chickens and other species close to the root of the avian family tree. But because emus, like other ratites, invest more of their body mass into leg muscles, they can carry out this precocial growth strategy to a greater extreme than flying birds, trading flight prowess away for enhanced running ability. This paper adds another important dataset to the oft-neglected area of “ontogenetic scaling” of the musculoskeletal system, or how the locomotor apparatus adapts to size-/age-related functional/developmental demands as it grows. Luis did a huge amount of work for this paper, leading arduous dissections and analysis of a complex dataset.

Superficial layer of leg muscles in an emu, in right side view. Click any image here to emu-biggen. The ILPO and IC are like human rectus femoris (“quads”); ILFB like our biceps femoris (“hams”); FL, GM and GL much like our fibularis longus and gastrocnemius (calf) muscles, but much much bigger! Or, perhaps FL stands for fa la la la la?

Data for an extra set of emus studied by coauthor Russ Main in the USA, which grew their muscles similarly to our UK group. The exponents (y-axis) show how much more strongly the muscles grew than isometry (maintaining the same relative size), which is the dotted line at 1. The numbers above each data point are the # of individuals measured. Muscle names are partly above; the rest are in the paper. If you want to know them, we might have been separated at birth!
2) Regnault, S., Pitsillides, A.A., Hutchinson, J.R. 2014. Structure, ontogeny and evolution of the patellar tendon in emus (Dromaius novaehollandiae) and other palaeognath birds. PeerJ 2:e711 http://dx.doi.org/10.7717/peerj.711
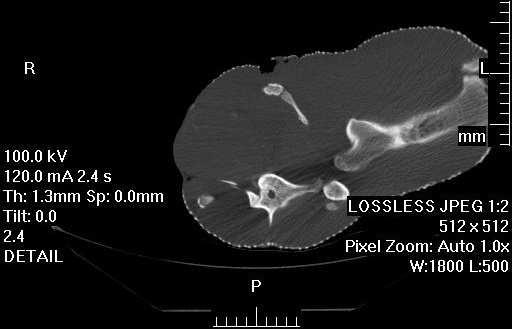
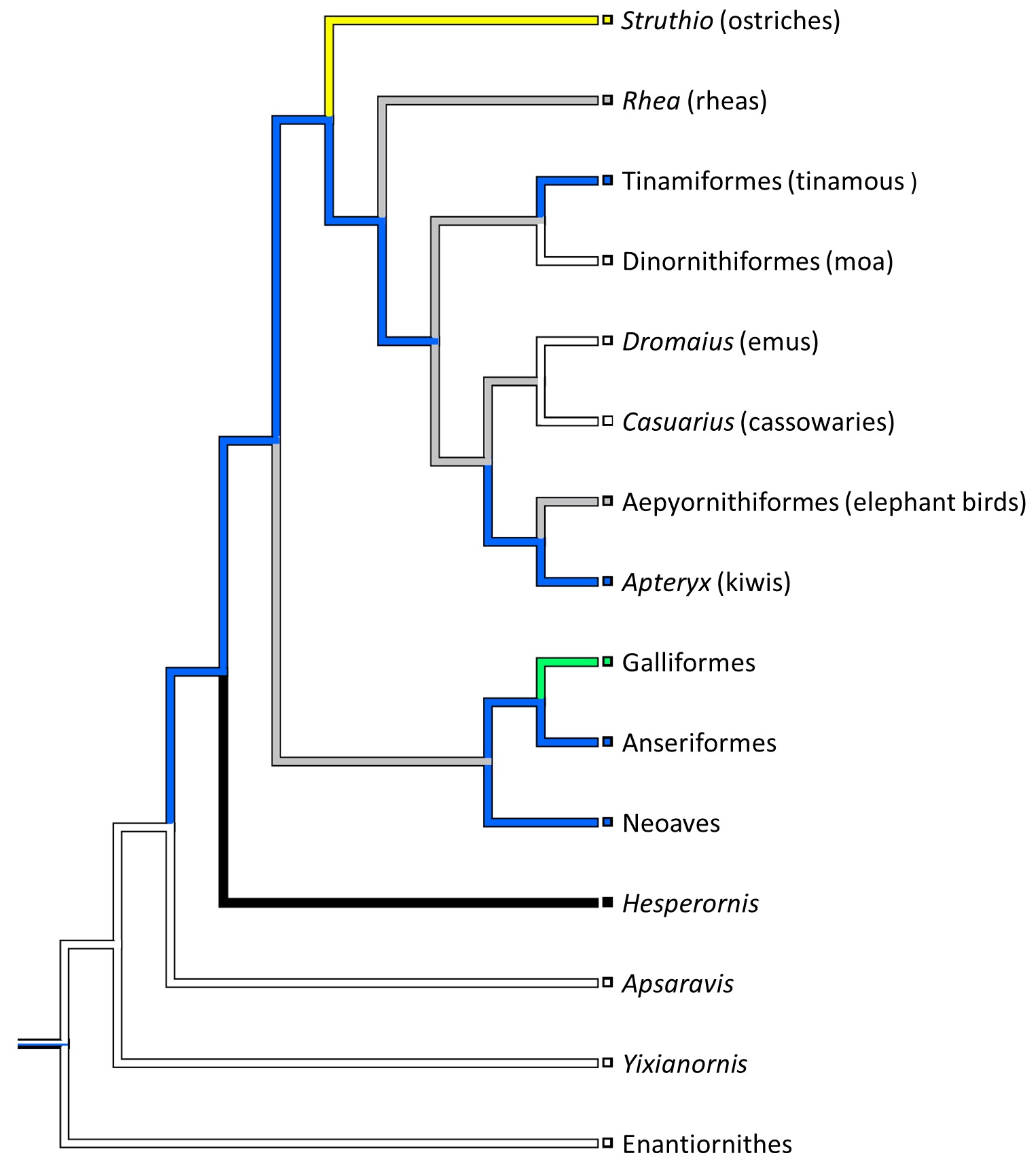
My second year PhD student Sophie Regnault (guest-blogger here before with her rhino feet post) has released her first PhD paper, on the evolution of kneecaps (patellae) in birds, with a focus on the strangeness of the region that should contain the patella in emus. This is a great new collaboration combining her expertise in all aspects of the research with coauthor Prof. Andy Pitsillides‘s on tissue histology and mine on evolution and morphology. This work stems from my own research fellowship on the evolution of the patella in birds, but Sophie has taken it in a bold new direction. First, we realized that emus don’t have a patella– they just keep that region of the knee extensor (~human quadriceps muscle) tendon as a fatty, fibrous tissue throughout growth, showing no signs of forming a bony patella like other birds do. This still blows my mind! Why they do this, we can only speculate meekly about so far. Then, we surveyed other ratites and related birds to see just how unusual the condition in emus was. We discovered, by mapping the form of the patella across an avian family tree, that this fatty tendon seems to be a thing that some ratites (emus, cassowaries and probably the extinct giant moas) do, whereas ostriches go the opposite direction and develop a giant double-boned kneecap in each knee (see below), whereas some other relatives like tinamous and kiwis develop a more “normal”, simple flake-like bit of bone, which is likely the state that the most recent common ancestor of all living birds had.
There’s a lot in this paper for anatomists, biomechanists, palaeontologists, ornithologists, evo-devo folks and more… plenty of food for thought. The paper hearkens back to my 2002 study of the evolution of leg tendons in tetrapods on the lineage that led to birds. In that study I sort of punted on the question of how a patella evolved in birds, because I didn’t quite understand that wonderful little sesamoid bone. And now, 12 years later, we do understand it, at least within the deepest branches of living birds. What happened further up the tree, in later branches, remains a big open subject. It’s clear there were some remarkable changes, such as enormous patellae in diving birds (which the Cretaceous Hesperornis did to an extreme) or losses in other birds (e.g., by some accounts, puffins… I am skeptical)– but curiously, patellae that are not lost in some other birds that you might expect (e.g., the very non-leggy hummingbirds).

Fatty knee extensor tendon of an emu, showing the absence of a patella. The fatty tissue is split into superficial (Sup) and deep regions, with a pad corresponding to the fat pad in other birds continuous with it and the knee joint meniscus (cushioning pad). The triceps femoris (knee extensor) muscle group inserts right into the fatty tendon, continuing on over it. A is a schematic; B is a dissection.

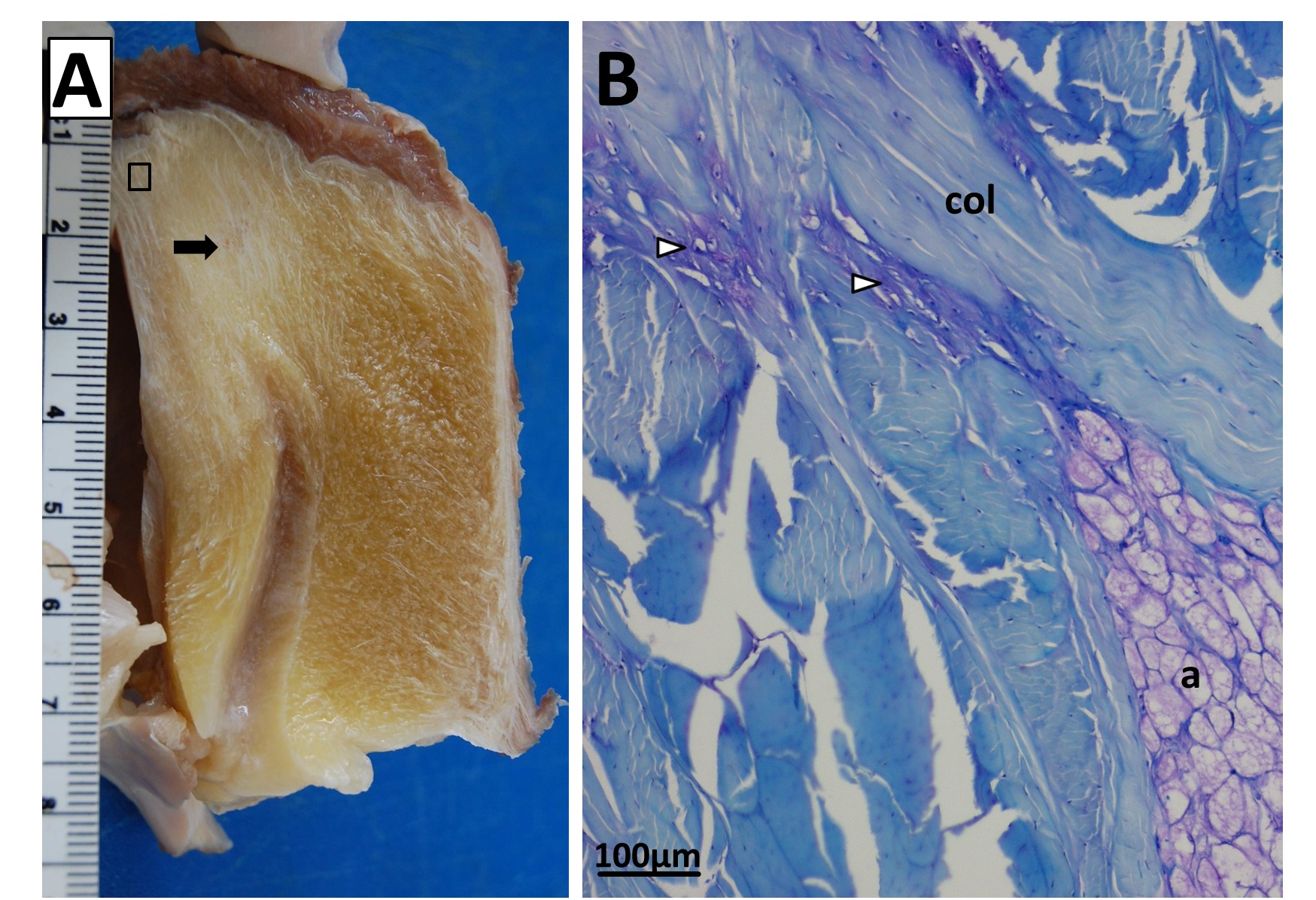
Sectioning of a Southern Cassowary’s knee extensor tendon, showing: A, Similar section as in the emu image above. revealing similar regions and fibrous tissue (arrow), with no patella, just fat; and B, With collagen fibre bundles (col), fat cells (a), and cartilage-like tissue (open arrows) labelled.

Evolution of patellar form in birds. White branches indicate no patella, blue is a small flake of bone for a patella, green is something bigger, yellow is a double-patella in ostriches, black is a gigantic spar of bone in extinct Hesperornis and relatives, and grey is uncertain. Note the uncertainty and convergent evolution of the patella in ratite birds (Struthio down to Apteryx), which is remarkable but fits well with their likely convergent evolution of flightlessness and running adaptations.
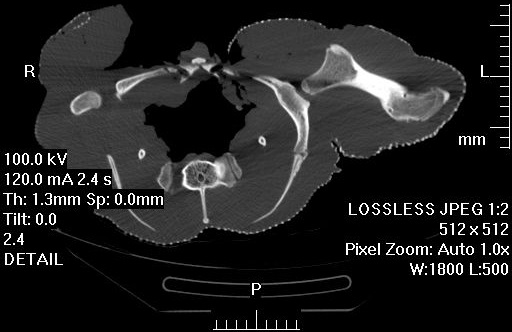
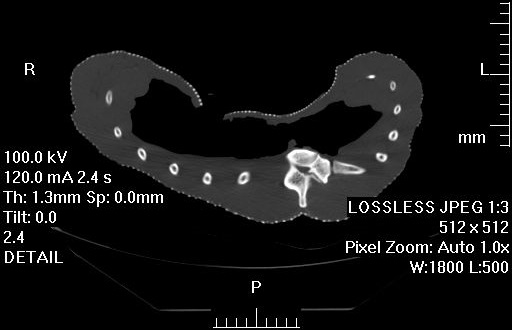
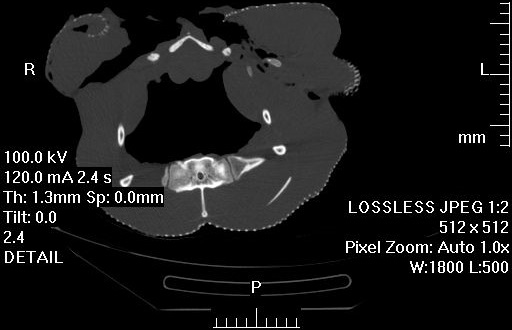
3) Chadwick, K.P., Regnault, S., Allen, V., Hutchinson, J.R. 2014. Three-dimensional anatomy of the ostrich (Struthio camelus) knee joint. PeerJ 2:e706 http://dx.doi.org/10.7717/peerj.706
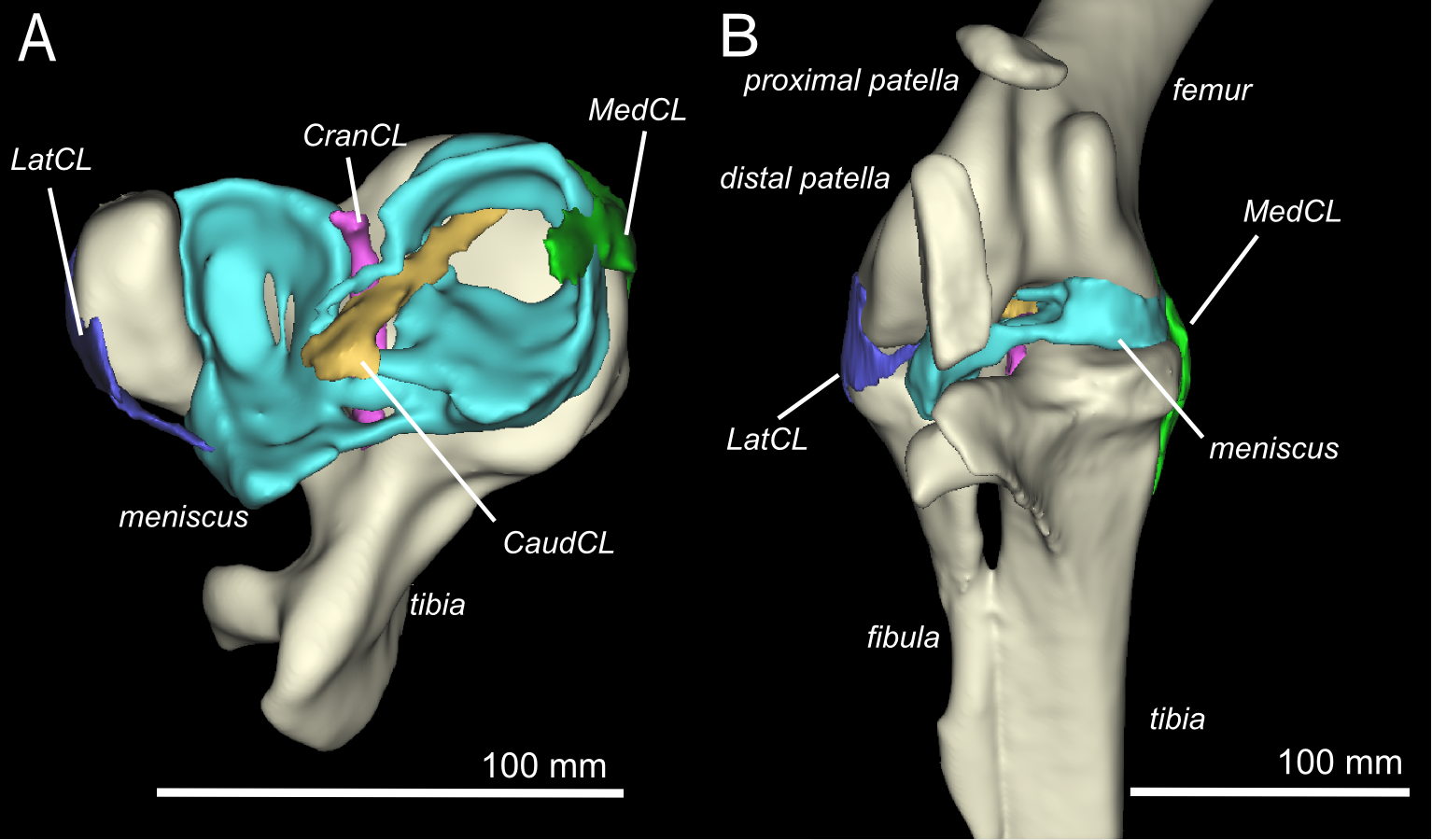
Finally, Kyle Chadwick came from the USA to do a technician post and also part-time Masters degree with me on our sesamoid grant, and proved himself so apt at research that he published a paper just ~3 months into that work! Vivian Allen (now a postdoc on our sesamoid bone grant) joined us in this work, along with Sophie Regnault. We conceived of this paper as fulfilling a need to explain how the major tissues of the knee joint in ostriches, which surround the double-patella noted above, all relate to each other and especially to the patellae. We CT and MRI scanned several ostrich knees and Kyle made a 3D model of a representative subject’s anatomy, which agrees well with the scattered reports of ostrich knee/patellar morphology in the literature but clarifies the complex relationships of all the key organs for the first time.
This ostrich knee model also takes Kyle on an important first step in his Masters research, which is analyzing how this morphology would interact with the potential loads on the patellae. Sesamoid bones like the patella are famously responsive to mechanical loads, so by studying this interaction in ostrich knees, along with other studies of various species with and without patellae, we hope to use to understand why some species evolved patellae (some birds, mammals and lizards; multiple times) and why some never did (most other species, including amphibians, turtles, crocodiles and dinosaurs). And, excitingly for those of you paying attention, this paper includes links to STL format 3D graphics so you can print your own ostrich knees, and a 3D pdf so you can interactively inspect the anatomy yourself!

Ostrich knee in side view: A, X-ray, and (B) labelled schematic.

3D model of an ostrich knee, showing: A, View looking down onto the top of the tibia (shank), with the major collateral ligaments (CL), and B, View looking straight at the front of the knee joint, with major organs of interest near the patella, sans muscles.
You can view all the peer review history of the papers if you want, and that prompts me to comment that, as usual at PeerJ (full disclosure: I’m an associate editor but that brings me £0 conflict of interest), the peer review quality was as rigorous at a typical specialist journal, and faster reviewing+editing+production than any other journal I’ve experienced. Publishing there truly is fun!
Merry Christmas and Happy Holidays — and good Ratite-tidings to all!
And stay tuned- the New Year will bring at least three more papers from us on this subject of ratite locomotion and musculoskeletal anatomy!
♬Should auld palaeognathans be forgot,
And never brought for scans?
Should publications be soon sought,
For auld ratite fans!♪
Read Full Post »